
Abstract
Recombinant adeno-associated viral (AAV) vectors have unique properties, which make them suitable vectors for gene transfer. Here we assess the liver transduction efficiency and biodistribution of AAV-pseudotyped capsids (serotypes) 1, 5, 6, and 8, combined with single-stranded and double-stranded genomic AAV2 structures carrying the luciferase reporter gene after systemic administration. The analysis was performed in vivo and ex vivo, in male and female mice. Gender-related differences in AAV-mediated transduction and biodistribution were shown for the four serotypes. Our data confirm the superiority of AAV8 over the rest of the serotypes, as well as a significant advantage of double-stranded genomes in terms of liver transduction efficiency, particularly in females. Regarding biodistribution, AAV5 displayed a narrower tropism than the other serotypes tested, transducing, almost exclusively, the liver. Interestingly, AAV1 and AAV8, in particular those having single-stranded genomes, showed high transduction efficiency of female gonads. However, no inadvertent germ line transmission of AAV genomes was observed after breeding single-stranded AAV8-injected female mice with untreated males. In conclusion, double-stranded AAV8 vectors led to the highest levels of liver transduction in mice, as demonstrated by luciferase expression. Nevertheless, the transduction of other organs with AAV8 vectors could favor the use of less efficient serotypes, such as AAV5, which display a narrow tropism.
Introduction
The transduction efficiency of AAV vectors in various somatic tissues is determined primarily by the viral capsid proteins (serotype). Indeed, the different serotypes are typically characterized by unique receptor tropism (Rabinowitz et al., 2002; Wu et al., 2006). After administration, different serotype AAV vectors transduce hepatocytes with different efficiencies, as some serotypes have tropism for other tissues with species-specific differences (Davidoff et al., 2005; Gao et al., 2006; Grimm et al., 2006; Nathwani et al., 2006). Interestingly, one characteristic related to AAV2-mediated liver transduction is the differences found at the level of gene expression depending on animal gender (Davidoff et al., 2003; Berraondo et al., 2006). Higher levels of transgene expression are observed in male mice and woodchucks, compared with females after intravenous injection of AAV2 (Berraondo et al., 2005). It has been reported that androgens play an important role in this phenomenon, although the molecular basis is not fully understood (Davidoff et al., 2003).
Single- to double-stranded DNA conversion is both essential and rate-limiting in the recombinant AAV vector-mediated transduction process. To overcome this step, several investigators have developed AAV containing double-stranded DNA (dsDNA) genomes (self-complementary or scAAV) by mutating one of the flanking inverted terminal repeats (ITRs) (McCarty et al., 2001, 2003; Wang et al., 2003) leading to the formation of a double-stranded AAV genome if below the maximal packaging limit. The use of this AAV double-stranded genomic structure has been shown to induce a 10- to 20-fold higher level of transgene expression than the same dose of single-stranded AAV (ssAAV) (Wang et al., 2003; Nathwani et al., 2006).
An aspect of viral gene transfer that deserves special attention for safe clinical application is that administration of the vector must not lead to germ line transmission. Several studies have shown that whereas systemic administration of AAV serotype 2 results in gene transfer to the gonads, no germ line transmission of vector sequences was detected (Arruda et al., 2001). Information regarding inadvertent AAV gene transfer into germ cells by other AAV serotypes has not been reported.
The purpose of this study was to evaluate the ability of AAV serotypes 1, 5, 6, and 8 to deliver the reporter gene luciferase to female and male mouse liver. We have also compared the effect of the AAV genomic structure on liver transduction, using all four serotypes. The analysis was performed with a noninvasive optical bioluminescence imaging system that allowed long-term in vivo follow-up of luciferase gene expression. Ninety-two days after administration of AAV-Luc vectors, the animals were killed and the biodistribution of the various serotypes was analyzed ex vivo. Furthermore, germ line transmission by single-stranded AAV8 was analyzed.
Materials and Methods
Animals
BALB/c mice (6 to 8 weeks old) were obtained from Harlan Laboratories (Barcelona, Spain) and maintained according to the guidelines of our institution. For all procedures, mice were anesthetized with ketamine–xylazine.
Construction of AAV single-stranded/double-stranded vectors
The pro-AAV-luciferase plasmids used in this study were ssAAV-phPBGD-Luciferase-PolyA and dsAAV-phPBGD-Luciferase-PolyA. The expression cassette contained in both plasmids consists of the reporter gene luciferase (GenBank accession number M15077) under the transcriptional control of the constitutive and ubiquitous promoter of human porphobilinogen deaminase (phPBGD, sequence 816–986; GenBank accession number M95623). In the ssAAV-phPBGD-Luciferase-PolyA construct the expression cassette is flanked by both AAV2 wild-type ITRs. An appropriate stuffer sequence and a poly(A) sequence from the rabbit β-globin gene were added to adjust the size of the AAV genome to the optimal packaging capacity described for single-stranded AAV vectors (Dong et al., 1996). In the dsAAV-phPBGD-Luciferase-PolyA plasmid, the stuffer was omitted and a minimal synthetic poly(A) sequence was used to adjust the size of the AAV genome to the optimal packaging capacity described for the generation of a double-stranded AAV genome (McCarty et al., 2001). The expression cassette in this case was flanked by a truncated 5′ ITR (the transfer pro-dsAAV plasmid was obtained from Applied Viromics [Fremont, CA]). In vivo analysis of luciferase expression after hydrodynamic injection of mice demonstrated comparable levels of luciferase expression between the two plasmids (data not shown).
Preparation of AAV vectors: Production, purification, and titration
rAAV serotype 1, 5, 6, and 8 vectors with wild-type AAV2 ITRs were produced by calcium phosphate-mediated cotransfection in HEK-293 cells. For each production the pro-AAV-luciferase plasmid and one (AAV1, −5, and −6) or two (AAV8) packaging plasmids were cotransfected into HEK-293 cells. The packaging plasmids were as follows: pVD20 for AAV1; pDP5 and pDP6 for AAV5 and AAV6, respectively; and two packaging plasmids named pAdDeltaF6 and p5E18-VD8 were used for AAV8 (Gao et al., 2002a,b; Grimm et al., 2003; Ross et al., 2006; Cheng et al., 2007). Briefly, eight viral batches were produced in HEK-293 cells after cotransfection of the respective plasmids (Graham and van der Eb, 1973). The cells were harvested 48 hr after transfection and virus was released from the cells by three rounds of freeze–thawing. Crude lysate from all batches was then treated with Benzonase (50 U/ml crude lysate) for 1 hr at 37°C and then kept at −80°C until purification. Purification of crude lysate was performed by anion-exchange chromatography (AAV2/8) (Hermens et al. 1999; Zolotukhin et al., 1999) or by affinity chromatography using AAV-specific llama antibodies (AVB Sepharose, Cat. No. 28-4112-01; GE Healthcare Life Sciences, Piscataway, NJ) according to the instructions of the manufacturer (AAV2/1, AAV2/5, and AAV2/6). The purified batches were concentrated and diafiltrated by cross-flow filtration (Spectrum Laboratories, Rancho Dominguez, CA) with a molecular mass cutoff of 400 kDa. The batches were then concentrated further by passage through Centricon tubes (YM-100; Millipore, Bedford, MA) to a final concentration of 1 × 1012 genome copies (GC)/ml, as determined by quantitative polymerase chain reaction (Q-PCR). After concentration, the viral batches were filtered (pore size, 0.22 μm) and stored at −80°C. Viral titers in terms of genome copies per milliliter were determined by Q-PCR, performed three times in triplicate at three different dilutions. Q-PCR was performed (Applied Biosystems, Foster City, CA) with primers pr214fw (5′-CTGGATACCGGGAAAACGCT-3′) and pr215rv (5′-ACCGGACATAATCATAGGACCTCT-3′), amplifying a 70-bp fragment from the phPBGD promoter region. Capsid protein composition and purity were analyzed by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE). An in vitro infectivity assay was performed in various cell lines (data not shown).
Bioluminescence measurement in living mice
Bioluminescence measurement was performed as previously described (Berraondo et al., 2006). Briefly, mice were anesthetized with ketamine–xylazine and 100 μl of
Data analysis
All data were analyzed with Monolix 2.4, software dedicated to the analysis of nonlinear mixed effect models (
Two different parameters describe the kinetics of luciferase expression, L max and T 50 (Berraondo et al., 2006), where L max represents the maximal increase in bioluminescence from baseline and T 50 represents the time necessary to reach the bioluminescence value L max/2. Statistical analysis was performed by two-way analysis of variance (ANOVA) followed by multiple comparison, using the least significant difference test (SPSS software; SPSS, Chicago, IL). The effect of a single factor (gender, serotype, genomic structure) over luciferase expression was statistically evaluated by Student t test.
Ex vivo biodistribution
Mice were killed 92 days after rAAV injection. The following organs were extracted according to the protocol of Gonin and Gaillard (2004): brain, lung, pancreas, muscle, spleen, heart, kidney, stomach, intestine, uterus, ovaries, testes, and liver. Luciferase activity was analyzed in 40-mg tissue samples homogenized in 250 μl of passive lysis buffer (Promega, Madison, WI). Homogenates were centrifuged at 20,800 × g for 1 min at 4°C and 20 μl of each supernatant was subsequently analyzed for luciferase activity, using a tube luminometer (Lumat LB 9507; Berthold Technologies, Bad Wildbad, Germany) and a luciferase assay kit (Promega). To determine the numbers of AAV genome copies in injected mice, DNA from various tissues was isolated with a QIAamp DNA mini kit (Qiagen, Valencia, CA) according to the manufacturer's instructions. Q-PCRs were set up in a volume of 20 μl, consisting of DNA SYBR green master mix (Bio-Rad, Hercules, CA) with the primer pair LucF (CTTGCTCATGCTACAGCTGGG) and LucR (GGAATTGGAACATTGCGACA), which results in amplification of a 70-bp region within the luciferase gene. Integrity of the DNA was confirmed by amplifying a glyceraldehyde-3-phosphate dehydrogenase (GADPH) gene, using previously described primers (Berraondo et al., 2006).
Luciferase immunohistochemistry
Immunohistochemical staining for luciferase was performed with a primary goat anti-luciferase antibody (Cortex Biochem/Promega). Immunohistochemistry was performed with an EnVision+ system (Dako, Glostrup, Denmark) according to the manufacturer's recommendations. Formalin-fixed, paraffin-embedded tissue sections (3 μm) were sequentially deparaffinized, blocked for endogenous peroxidase activity with 3% hydrogen peroxidase in methanol for 10 min, and rehydrated.
Incubation with primary antibody were performed overnight at 4°C. After a brief rinse in Tris-buffered saline with Tween 20 (TBS-T), the sections were incubated with horseradish peroxidase-labeled anti-goat antibody (Dako) for 30 min at room temperature. Peroxidase activity was revealed with 3,3′-diaminobenzidine (DAB+; Dako) and sections were counterstained with Harris hematoxylin. Last, slides were dehydrated in a graded series of ethanol, cleared in xylene, and mounted in DPX resin.
Germ line transmission experiment
Twelve BALB/c female mice were intravenously injected with a single dose of ssAAV8-hPBGD-Luciferase-PolyA (5 × 1012 GC/kg) in a volume of 200 μl (n = 6). Six weeks after AAV injection, female mice were crossed with naive males of the same strain. Resulting progeny were analyzed for luciferase expression 4 weeks after birth, using the Xenogen IVIS system, as previously described. The presence of AAV viral genomes were analyzed by Q-PCR after genomic DNA extraction from tail tissue, as described.
Results and Discussion
In vivo analysis of luciferase expression after intravenous administration of rAAVs harboring single-stranded and double-stranded genomes
This study was designed to determine the effects of AAV serotype, AAV genomic structure, and animal gender on AAV-mediated liver transduction and biodistribution, using noninvasive imaging technology. AAV capsids 1, 5, 6, and 8 were used to package rAAV2 genome containing the luciferase expression cassette under the control of a constitutive and ubiquitous PBGD promoter. rAAV2 vectors were excluded from this study because the low transduction efficiency of hepatocytes is well reported (Miao et al., 2000). In addition, any clinical application of this serotype is hampered by the high prevalence of neutralizing antibodies in the human population (Halbert et al., 2006).
Male and female BALB/c mice were injected via the tail vein with a single dose (5 × 1012 GC/kg) of single-stranded or double-stranded rAAV vector serotype 1, 5, 6, or 8 (n = 5 or 6 per group). Whole body imaging was performed from day 1 postinjection, up to 91 days postinjection. Luciferase expression was quantified by tracing regions of interest (ROIs) in the upper abdominal area, including the liver. Our results showed that all the serotypes tested led to sustained expression of luciferase in the liver (see Fig. 1). Luciferase expression increased significantly from 1 to 14 days after administration and thereafter reached a plateau for the duration of the experiment.
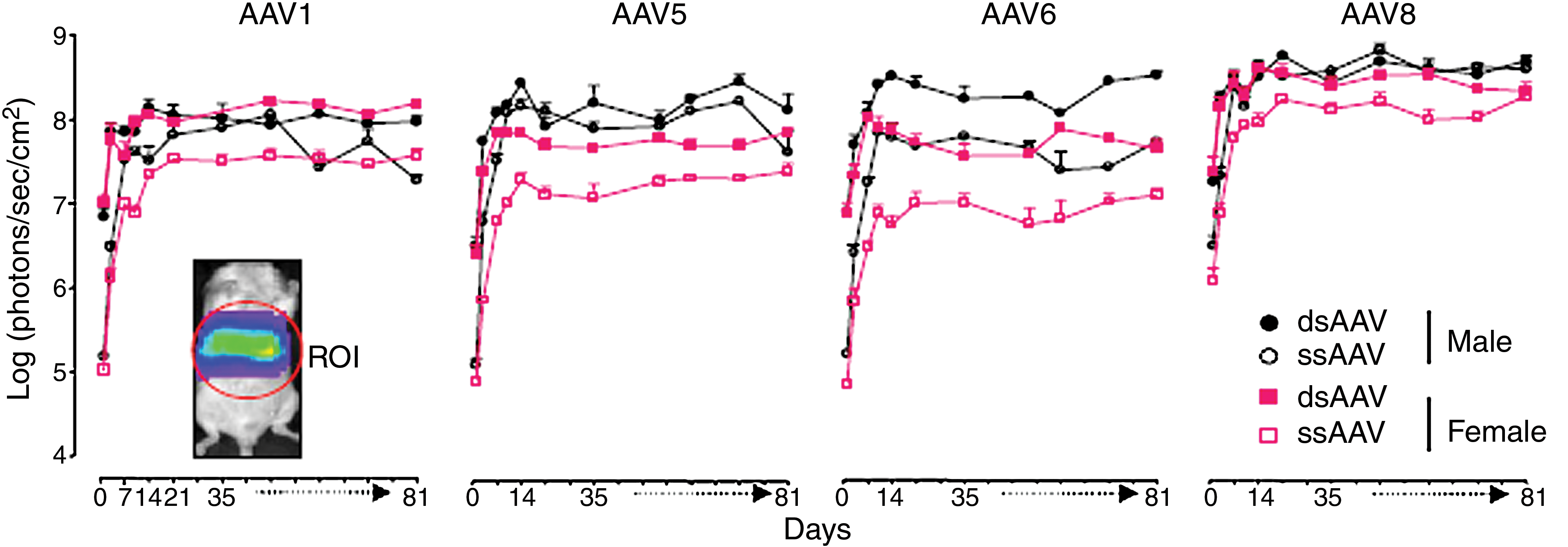
Long-term in vivo CCD signal intensity in the abdominal area of mice injected with rAAV-Luc. BALB/c male and female mice (n = 5 or 6 per group) were injected intravenously with AAV serotype 1, 5, 6, or 8 (5 × 1012 GC/kg) harboring single-stranded or double-stranded AAV-luciferase DNA genome. Imaging was performed for luciferase expression, using
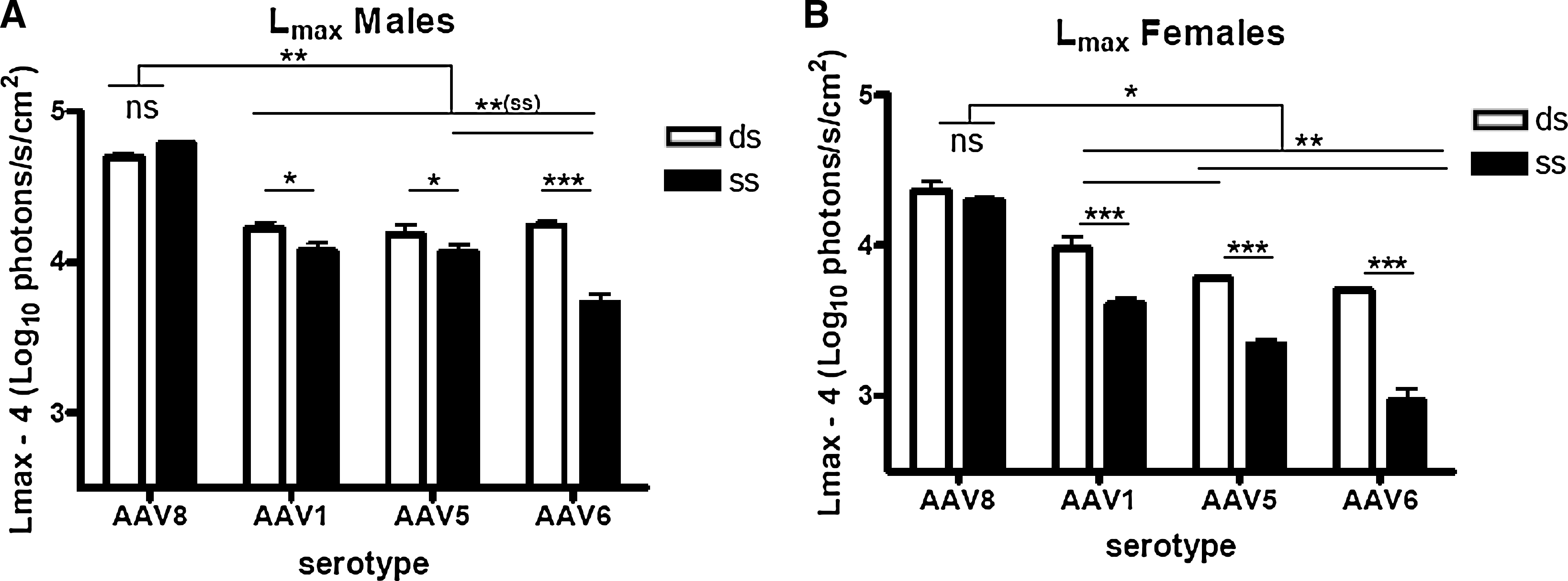
Because of the fluctuations in luciferase activity over time, instead of analyzing luciferase expression at a single time point, the bioluminescence data were analyzed using a pharmacokinetic model. This model allows calculation of two parameters that describe the kinetics of luciferase expression after AAV injection: L max and T 50 (Berraondo et al., 2006). As shown in Fig. 2A and B, significant differences in luciferase expression levels were observed depending on animal gender, rAAV serotype, and genomic structure. L max was higher in male animals for all the serotypes. Compared with other serotypes, AAV8 demonstrated the highest efficiency of transduction of the liver in mice of both genders, independent of the genomic structure (single or double stranded). Interestingly, in contrast with previous reports, no significant difference was observed between single- and double-stranded AAV8. AAV serotypes 1, 5, and 6 harboring double-stranded genomic structures displayed similar transduction efficiencies in male mice (Fig. 2A) whereas in females, dsAAV1 transduction was higher than that of dsAAV5 and dsAAV6 (Fig. 2B). For vectors with single-stranded genomes, in males AAV6 exhibited significantly reduced transduction efficacy compared with the other serotypes tested (Fig. 2A). In females, the highest expression from ssAAV vectors was obtained with serotype 1 followed by AAV5 and AAV6 (Fig. 2B).
(
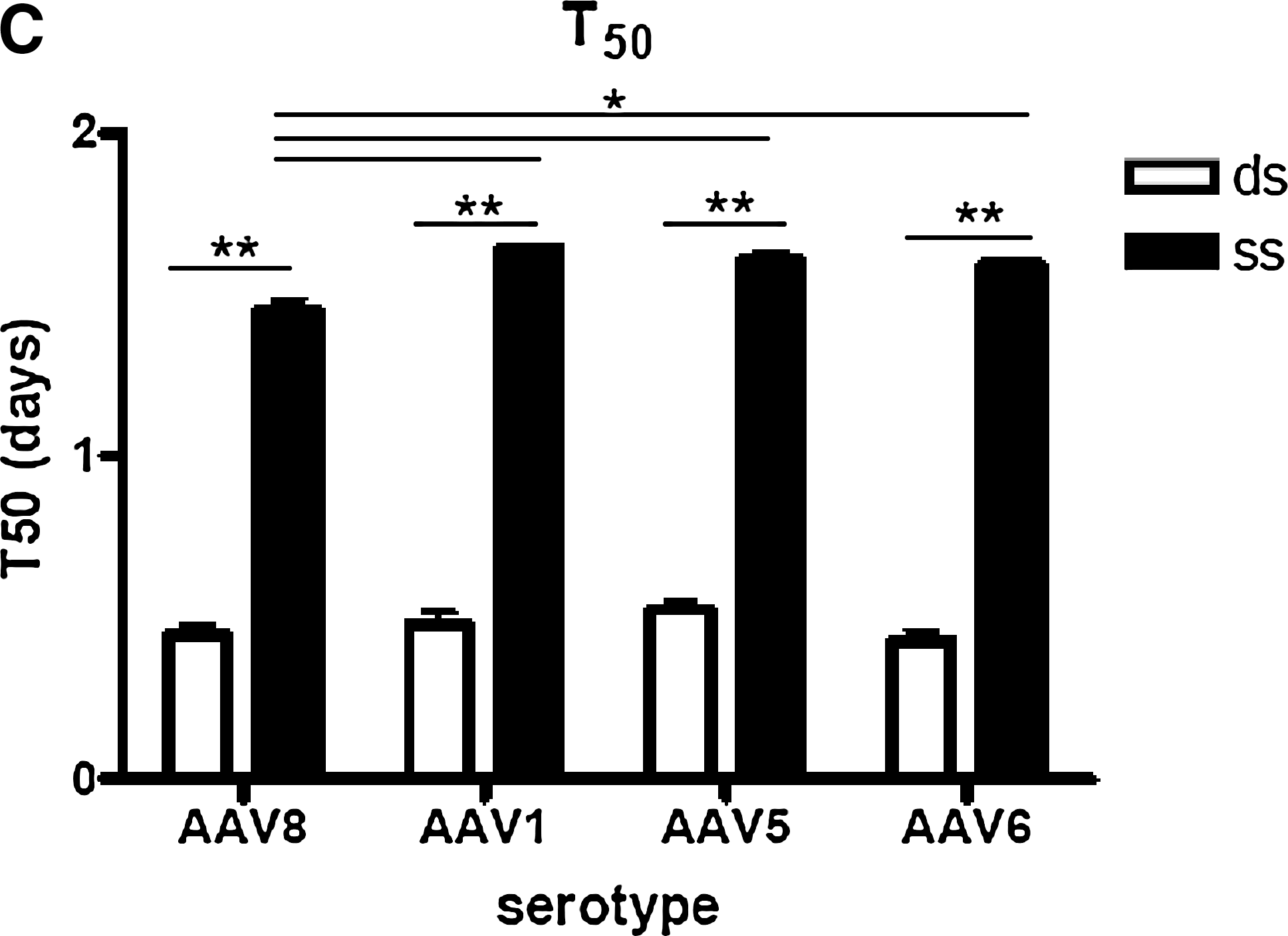
The statistical analysis of T 50 values, which inversely correlate with the time of onset of transgene expression, showed no gender-dependent effect. Thus, male and female mice receiving the same vector were grouped. Significant differences dependent on the genomic structure were observed (Fig. 2C). For the serotypes tested, the T 50 value for dsAAV was significantly lower the T 50 value for ssAAV, indicating that dsAAV genomes were expressed in mouse livers with a faster onset than single-stranded vectors, in accordance with other reports (McCarty et al., 2001, 2003; Wang et al., 2003; Thomas et al., 2004) (Fig. 2C). The T 50 value for ssAAV8 was lower than the T 50 value for the rest of the serotypes with single-stranded genomes (Fig. 2C). This might be related to the faster rate of decapsidation in the nucleus reported for this serotype by Thomas and coworkers (2004).
Dose escalation study using dsAAV8 and ssAAV8
Because of reported significant differences in liver transduction efficiency between ssAAV8 and dsAAV8 vectors (Nathwani et al., 2006), we wanted to explore whether the minor differences between ssAAV and dsAAV genomes observed in this study were due to the administration of saturating amounts of virus. To answer this question, a dose escalation study was performed with these two rAAV vectors. In this experiment 30 female BALB/c mice were injected intravenously with AAV8 at 5 × 1012, 5 × 1011, or 5 × 1010 GC/kg in their two genomic forms (n = 5 per group). Remarkable differences in liver transduction between ssAAV8 and dsAAV8 were observed when the mice received a dose 10 or 100 times lower than 5 × 1012 GC/kg (Fig. 3A). We found a near linear correlation between luciferase expression and vector dose in the double-stranded setting. Nevertheless, for the AAV8 single-stranded genome, the vector dose/expression relation was not linear. Analysis of L max values corroborated these findings (Fig. 3B). Interestingly, as shown in Fig. 3C, T 50 values depended also on the dose of virus administered. The T 50 values were higher for the lowest doses, indicating that transgene expression depends on the number of genome copies that enter into a cell, which could affect the formation of transcriptionally active AAV structures.

Long-term in vivo CCD signal intensity in the abdominal area of mice injected with AAV8-Luc. (

Biodistribution
All the serotypes tested in this study transduce the murine liver. However, luciferase signals were also detected within other organs. A representative image of one male and one female mouse per group is depicted in Fig. 4A. In Fig. 4, the intensity of luciferase expression among the animals cannot be compared, because the light intensity scale has been adjusted for each animal to demonstrate extrahepatic expression. Female mice that received ssAAV6 or dsAAV6 showed strong luciferase expression in their lower limbs. We observed significant luciferase expression in the lower abdominal area of female but not male mice injected with AAV1 and AAV8, particularly when the AAV genome was single-stranded. Analysis of the L max value revealed that ssAAV1 and ssAAV8 transduced more efficiently cells of the lower abdominal area than did double-stranded genomes (Fig. 4B). This is in clear contrast to the efficiencies of these serotype vectors observed when transducing the liver as double-stranded genomes (Fig. 2). AAV5 single-stranded and double-stranded vectors showed no significant extrahepatic expression in mice of both genders.
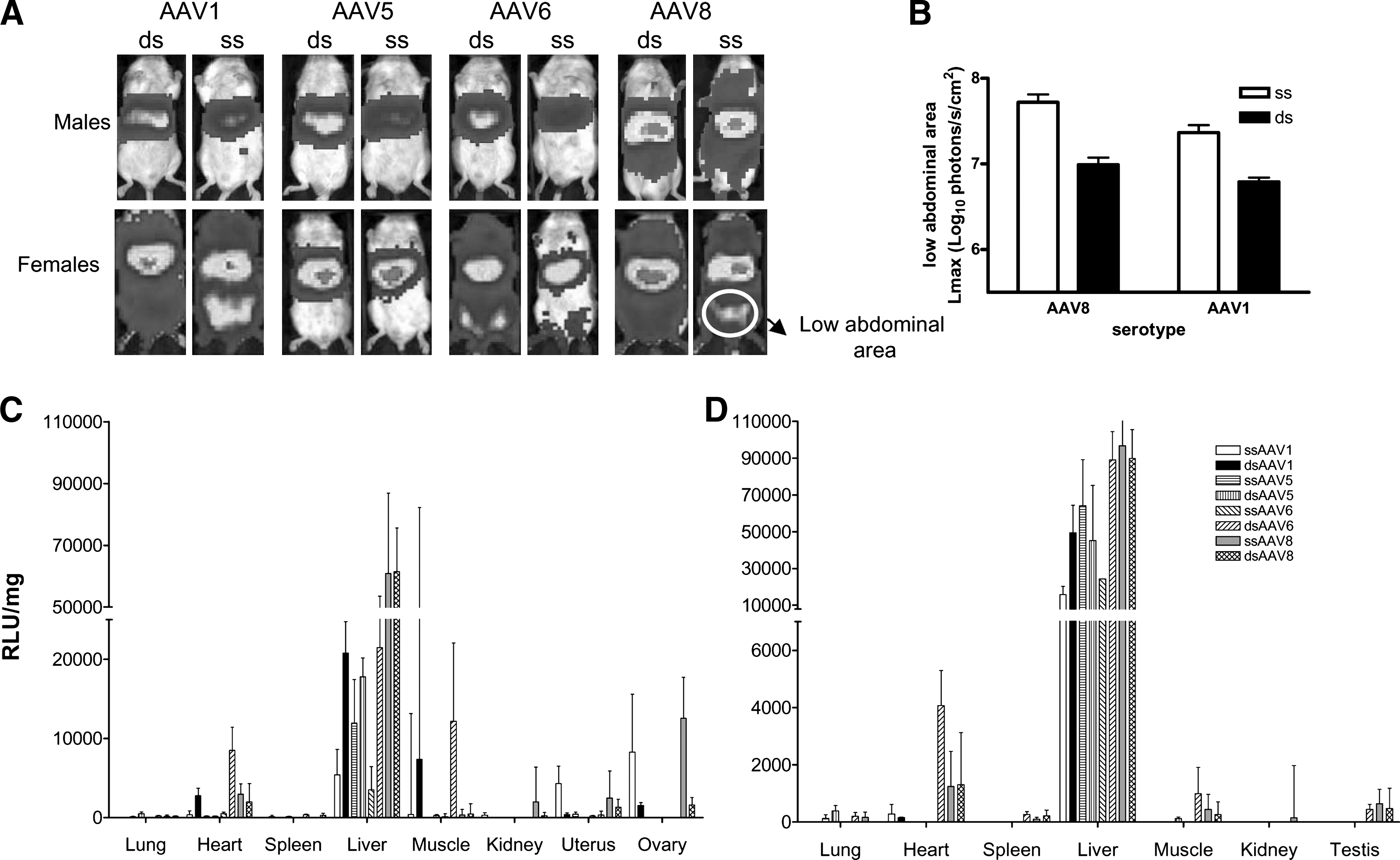
Bioluminescence imaging of AAV-luciferase gene expression in living mice, using a cooled CCD camera and quantification of luciferase activity in the abdominal area and ex vivo analysis of luciferase activity in various organs. Six-week-old BALB/c male and female mice were injected with AAV serotypes 1, 5, 6, and 8 (5 × 1012 GC/kg) harboring single- or double-stranded DNA genomes containing the luciferase gene. The mice were repeatedly scanned with the CCD camera, as described in the caption to Fig. 1. Shown are optical CCD images for luciferase expression of representative female and male animals from each group, 1 month postinjection (once a steady state level of luciferase expression was reached). Luciferase signal was enhanced in order to highlight extrahepatic luciferase expression. (
To determine the origin of extrahepatic expression with these vectors, mice were killed 91 days after injection and 12 organs were dissected (see Materials and Methods). Luciferase expression was analyzed in tissue extracts with a luminometer and the results are shown in Fig. 4C and D. Ex vivo data are consistent with in vivo analyses. For all the serotypes tested, the highest levels of luciferase expression were obtained from liver samples, where AAV8 led to the highest levels. The first observation was that all AAV serotypes displayed a narrower tropism in male than in female mice. Ex vivo analysis also confirmed the narrowing of tropism of AAV serotype 5. After injection of this serotype vector, luciferase expression was detected predominantly in the liver. Surprisingly, AAV1 and AAV8 were also found to transduce the ovaries and the uterus of female mice, explaining the extrahepatic lower abdominal expression. Luciferase expression levels were higher in the ovaries and uterus of ssAAV1- and ssAAV8-injected female mice as compared with dsAAV1- and dsAAV8-injected females. It will be of interest to elucidate the molecular basis for this observation. As previously described, we observed high levels of luciferase expression in muscle of the lower limbs after intravenous administration of AAV6. In this case, double-stranded vectors transduce the musculature more efficiently than do single-stranded vectors, mirroring the advantage of dsAAV over ssAAV vector constructs in the liver. The presence of viral genomes in the various organs was analyzed by Q-PCR and DNA levels correlated with luciferase expression (data not shown).
To further confirm transduction of the ovaries by AAV vectors, luciferase expression was analyzed by immunohistochemistry. As shown in Fig. 5, luciferase expression can be clearly detected in the ovaries of ssAAV8-Luc-injected mice. Transgene expression was located predominantly in central and peripheral stromal cells. No luciferase expression was detected in follicles or oocytes. The absence of previous data in this respect could be attributable to the fact that some studies used only male mice, and other studies used liver-specific promoters and hence female gonads, uterus, and ovaries were not analyzed (Sarkar et al., 2004; Nakai et al., 2005; Wang et al., 2005; Graham et al., 2008). However, our observations are in accordance with several articles reporting high prevalence of wild-type AAV in female genital samples (52.7% of the samples analyzed by PCR techniques) (Han et al., 1996; Venturoli et al., 2001).
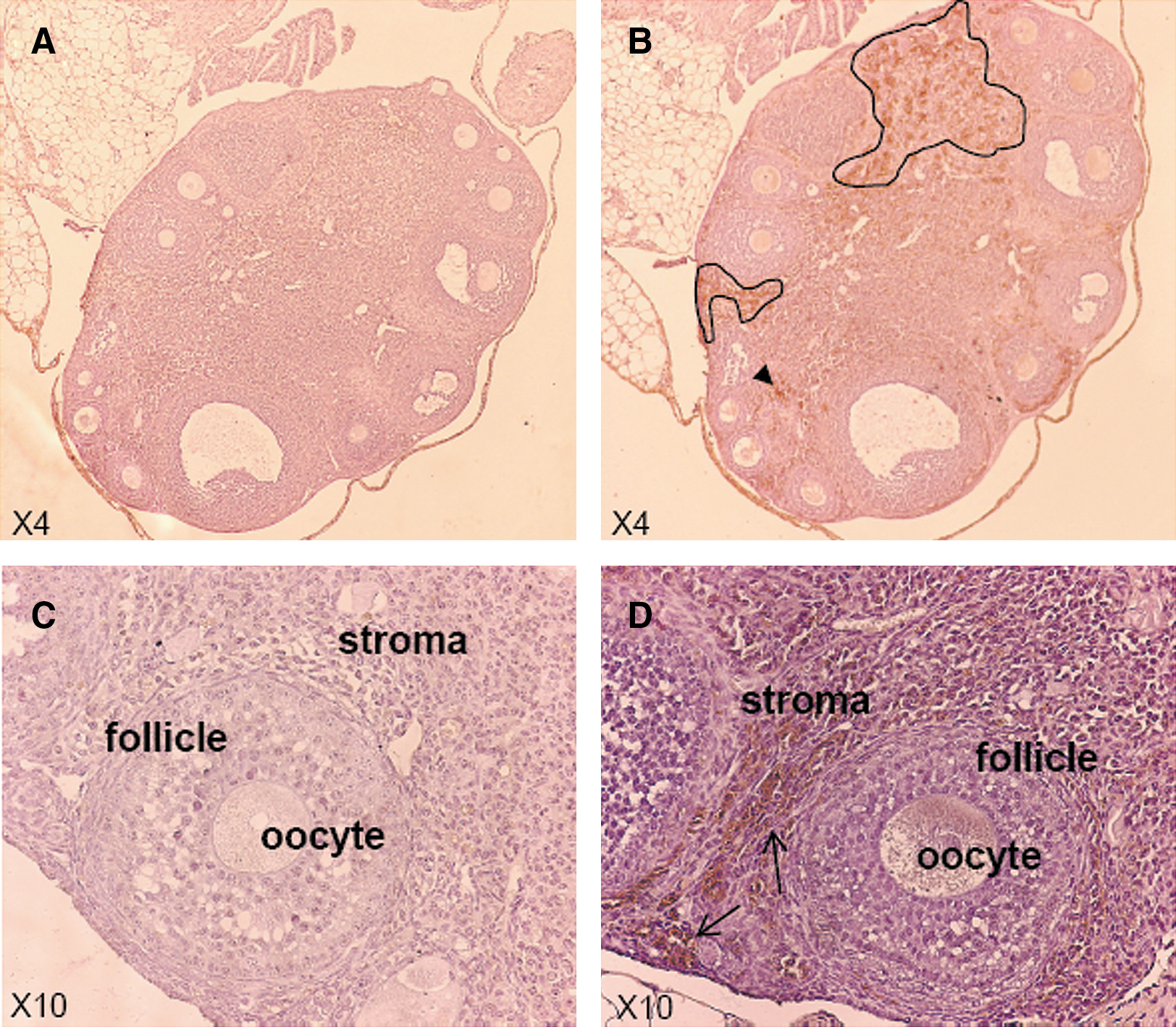
Luciferase expression is detected in the ovaries of ssAAV8-injected female mice. Six-week-old BALB/c female mice were injected with ssAAV8-Luc at a dose of 5 × 1012 GC/kg. Mice were killed 91 days after virus injection and ovaries were dissected and luciferase expression analyzed by immunohistochemistry. Outlined portions in
Germ line transmission
Because high levels of luciferase expression were detected in the gonads of female mice injected with ssAAV1 and ssAAV8, we wanted to test whether the injected mice could transfer the AAV genome to progeny. Female BALB/c mice (n = 6) were injected with ssAAV8-luciferase at 5 × 1012 GC/kg. Six weeks later, these injected females were subsequently bred with naive BALB/c males. Luciferase expression was analyzed in more than 100 pups 4 weeks after birth, but no luciferase expression was detected (Fig. 6). Because the activity of the promoter can be inhibited during development of the animals, the presence or absence of AAV DNA was determined from the genome of the BALB/c pups. There was no evidence of recombinant AAV DNA in any of the pups analyzed (data not shown).

Bioluminescence imaging of AAV-luciferase gene expression in injected female mouse and pups. Six-week-old BALB/c female mice were injected with AAV serotype 8 harboring single-stranded DNA genome containing the luciferase gene (dose, 5 × 1012 GC/kg). Six weeks after vector injection, females were crossed with naive BALB/c males. Optical CCD image for luciferase expression from female and male mice and their 4-week-old litters is shown. The light emission from female mice was analyzed separately from male and litters. Light acquisition from male and litters was performed at the maximal acquisition time recommended (5 min) whereas the light emitted by AAV-injected females was acquired for 1 sec. Luciferase expression was detected only in the AAV-injected female mother. A representative family from all the families in the study is shown.
Conclusion
In conclusion, we have carried out a detailed and quantitative study examining the effect of AAV serotype, AAV genomic structure, and animal gender on rAAV transduction efficiency and biodistribution in mice. We have confirmed the previous observation by other groups regarding the higher efficacy of AAV8 relative to the rest of the serotypes used in this study and the improved performance of double-stranded genomes over single-stranded genomes. Regarding biodistribution, we found that rAAV5 exhibited the narrowest tropism, transducing almost exclusively the liver. We found a wider distribution in females compared with males for all serotypes, as well as a significant transduction of female gonads by AAV1 and AAV8. On the basis of our data we would select AAV serotype 5 for liver applications. Although this serotype rAAV vector is not the most efficient in terms of transducing the liver, its biodistribution is more restricted and its presence was not detected in ovaries.
Footnotes
Acknowledgments
The authors acknowledge technical support from Cristina Olague, Nerea Gonzalez, and Africa Vales. This work was supported by grants from the UTE project CIMA; SAF 2006-03623 (Ministerio Educación y Ciencia) to G.G.-A., J.C., and P.B.; and by the Fundación de Investigación Médica Mutua Madrileña to G.G.-A. and A.F. A.P. was supported by the Fundación Inocente and by a Torres Quevedo research grant. L.V. is the recipient of an FPU fellowship. This work is part of the doctoral thesis of L.V.
Author Disclosure Statement
No competing financial interest exists for Astrid Pañeda, Lucia Vanrell, Itsaso Mauleon, Julien S. Crettaz, Pedro Berraondo, Jesus Prieto, Antonio Fontanellas, and Gloria Gonzalez-Aseguinolaza. Eric J. Timmermans, Stuart G. Beattie, Jaap Twisk, Sander van Deventer, and Maria Sol Rodriguez-Pena are AMT employees. Sander van Deventer owns stock of AMT.