
Abstract
Numerous preclinical studies have shown that osseous defects can be repaired by implanting bone morphogenetic protein (BMP)-2-transduced muscle cells. However, the drawback of this treatment modality is that it requires the isolation and long-term (∼3 weeks) culture of transduced autologous cells, which makes this approach cumbersome, time-consuming, and expensive. Therefore, we transferred BMP-2 cDNA directly to muscle tissue fragments that were held in culture for only 24 hr before implantation. We evaluated the ability of such gene-activated muscle grafts to induce bone repair. Two of 35 male, syngeneic Fischer 344 rats used in this study served as donors for muscle tissue. The muscle fragments remained unmodified or were incubated with an adenoviral vector carrying the cDNA encoding either green fluorescent protein (GFP) or BMP-2. Critical-size defects were created in the right femora of 33 rats and remained untreated or were filled (press fitted) with either unmodified muscle tissue or GFP-transduced muscle tissue or with BMP-2-activated muscle tissue. After 6 weeks, femora were evaluated by radiography, microcomputed tomography (μCT), histology, and biomechanical testing. Six weeks after implantation of BMP-2-activated muscle grafts, 100% of the bone defects were bridged, as documented by radiographs and μCT imaging, and showed formation of a neocortex, as evaluated by histology. Bone volumes of the femora repaired by BMP-2-transduced muscle were significantly (p = 0.006) higher compared with those of intact femora and the biomechanical stability was statistically indistinguishable. In contrast, control defects receiving no treatment, unmodified muscle, or GFP-transduced muscle did not heal. BMP-2 gene-activated muscle grafts are osteoregenerative composites that provide an expedited means of treating and subsequently healing large segmental bone defects.
Introduction
Recombinant, human bone morphogenetic protein (BMP)-2 and BMP-7 are osteoinductive cytokines that have been approved for clinical use and constitute novel tools for spinal fusion and the treatment of long-bone fractures (Boden et al., 2000; Friedlaender et al., 2001; Reddi, 2001; Einhorn, 2003; Mont et al., 2004). However, such recombinant growth factors have short biological half-lives and are difficult to retain at the defect site (Talwar et al., 2001; Uludag et al., 2001). Therefore, there is great interest in developing gene therapy approaches. By delivering complementary DNAs that encode osteogenic growth factors to recipient cells it is possible to achieve high, sustained concentrations of these factors locally for sufficient periods of time (Scaduto and Lieberman, 1999; Lieberman et al., 2002; Evans et al., 2004).
Preclinical studies suggest the use of viral vectors such as adenoviruses, adeno-associated viruses, and retroviruses as carriers of BMP-2 cDNA (Baltzer et al., 2000; Lee et al., 2002; Chen et al., 2004; Sugiyama et al., 2005; O.B. Betz et al., 2006). Adenovirus may be the most suitable vector for bone repair because it is nonintegrating and able to infect a variety of cells with high transduction efficiency. In addition, as the in vivo expression profile after transduction with first-generation adenovirus is characterized by transgene expression at high levels for 2–3 weeks and complete loss of expression by 6 weeks (Verma and Somia, 1997; Cao et al., 2004), it may be well suited for bone healing from the perspectives of optimal activity window and safety.
Only a few research groups have successfully induced bone formation in animal models using direct, in vivo adenoviral gene delivery (Baltzer et al., 2000; Bertone et al., 2004; O.B. Betz et al., 2006). In the experiment of Baltzer and colleagues (2000) bone growth was induced in femoral bone defects in a rabbit model by intralesional injection of a first-generation adenovirus carrying the human BMP-2 gene (Ad.BMP-2). In previous studies, we evaluated and further developed the same method in a rat model (O.B. Betz et al., 2006, 2007; V.M. Betz et al., 2007). Although data from these studies are encouraging, in vivo gene delivery is unlikely to find applications in human clinical orthopedics unless vectors are available that display attractive safety profiles and allow specific cell targeting.
In contrast, ex vivo gene therapy is considered to be safe and allows us to transfer cDNA to selected cells under controlled conditions outside of the body. Bone marrow-derived cells (Lieberman et al., 1998; Zachos et al., 2007), fat-derived cells (Dragoo et al., 2003), and skin cells (Krebsbach et al., 2000) engineered to express osteogenic genes have successfully induced bone formation in vivo. Muscle tissue represents another attractive source of cells useful for tissue engineering because it is relatively easy to harvest with low donor site morbidity. Huard and colleagues have extensively demonstrated that genetically engineered muscle-derived cells differentiate toward the osteogenic lineage and repair bone defects in a variety of animal models (Peng et al., 2002; Shen et al., 2004).
Although ex vivo gene therapy has proven to be an effective bone repair strategy, the disadvantage of this approach is that it requires the isolation and expansion of autologous progenitor cells, which makes this treatment modality cumbersome, time-consuming (∼3 weeks), and expensive (Evans et al., 2007).
Therefore, as an effort to develop an expedited ex vivo gene therapy approach, we transferred BMP-2 cDNA directly to muscle tissue fragments that were held in culture for only 24 hr before implantation. We subsequently evaluated the ability of such expedited gene-activated muscle grafts to induce bone healing in a well-established rat femoral segmental defect model.
Materials and Methods
Study design
A syngeneic rodent autotransplant model (male Fischer 344 rats, weighing 300–350 g) was used in this experiment. Muscle tissue was harvested from two killed donor rats. A 5-mm, critical-sized mid-femoral defect was created in the right hind limb of each of 33 rats and stabilized by an internal fixator. The rats were assigned to one of four groups. Bone defects received either muscle grafts activated by an adenoviral vector carrying the human BMP-2 cDNA (Ad.BMP-2) (n = 9) or muscle grafts transduced by an adenoviral vector carrying the gene encoding green fluorescent protein (Ad.GFP) (n = 8) or untreated muscle grafts (n = 8). The defects of the fourth group remained untreated (n = 8). Contralateral, intact femora were harvested from the group that received BMP-2-activated tissue grafts. At 3 and 6 weeks after surgery one animal of each group was killed and femora were harvested for histological evaluation. Healing of the remaining femora was evaluated by radiographs, microcomputed tomography (μCT), and biomechanical testing at 6 weeks after surgery.
The decision on n = 7 (BMP-2-activated tissue grafts) and n = 6 (control groups) animals per treatment group for the assessment of bone volume and biomechanical stability was based on a power analysis and our considerable experience with the here-employed animal model and similar gene transfer strategies for bone defect regeneration (O.B. Betz et al., 2006, 2007; V.M. Betz et al., 2007).
Vector production
Serotype 5, E1,E3-deleted, first-generation adenoviral vector was constructed by Cre–lox recombination as described earlier (Hardy et al., 1997). The transgenes were cloned into the E1 domain, with expression driven by the human cytomegalovirus early promoter. Recombinant adenoviruses were propagated in 293/Cre8 cells. High-titer preparations were generated by amplification in 293 cells, purification on CsCl gradients, and dialysis against 10 mM Tris-HCl (pH 7.8), 150 mM NaCl, 10 mM MgCl2, 4% sucrose buffer (Palmer et al., 2003). The viral titer was estimated as 4.9 × 1012 particles/ml by optical density and 6.3 × 1010 plaque-forming units (PFU)/ml by standard plaque assay.
Tissue harvest
Fischer 344 rats, a syngeneic breed, are genetically identical animals that allow transplantation of tissue from one individual to another without inducing a host-versus-graft immune response known from allograft transplantation (Iida et al., 2004; Benten et al., 2005; Niranjan et al., 2007). For practical and logistical reasons we used this well-established syngeneic rodent autotransplant model, harvested muscle tissue from two killed donor rats, and implanted the tissue into bone lesions of 33 syngeneic rats, thereby mimicking autograft transplantation.
Muscle tissue was harvested from the upper thighs of two animals, washed with phosphate-buffered saline, and transferred into a Petri dish. The tissue was then cut under sterile conditions into slices approximately 1 mm thick, using a tissue slicer (Stoelting tissue slicer; Stoelting, Wood Dale, IL). Disks were then punched out with a 4-mm dermal biopsy punch to create fragments of a standardized size and placed in 24-well plates. No cell isolation was performed.
Activation of tissue grafts
The following procedures and conditions for tissue graft activation have been optimized in previous pilot studies. Upregulation of BMP-2 after gene delivery was measured by enzyme-linked immunosorbent assay (ELISA).
Tissue fragments were infected with 1 × 108 PFU of Ad.BMP-2 and control tissue grafts were transduced with 1 × 108 PFU of Ad.GFP or remained untreated according to their group affiliation. Viral particles were appropriately diluted in phosphate-buffered saline (PBS) and a volume of 10 μl was applied directly to the surface of the tissue grafts, using a micropipette. The tissue fragments were then placed in an incubator (37°C, 5% CO2) for 60 min. To avoid dilution of the vector concentration no cell medium was added to the tissue. The virus solution and the humidified air in the incubator kept the tissue fragments moist during incubation. Dulbecco's modified Eagle's medium (DMEM; Biochrome, Berlin, Germany) was then added and the tissues were placed back in the incubator for 24 hr.
Measurement of BMP-2 upregulation by ELISA
Four muscle disks in 700 μl of DMEM per well were held in culture, using a 24-well plate. Medium was changed every third day. The supernatants of untreated muscle, GFP-transduced muscle, and BMP-2-transduced muscle were harvested on day 9 and BMP-2 upregulation was measured by ELISA. Measurements were performed in quadruplicate (n = 4).
Surgical procedure
All operative procedures were approved by the animal welfare committee of Bavaria, Germany (Regierung von Oberbayern). A modified version of an established, critical-size, femoral defect rat model (Einhorn et al., 1984; O.B. Betz et al., 2006, 2007) was used in this study. The animals were placed under general anesthesia by the administration of isoflurane, using a small-animal vaporizer, initially at 4% isoflurane in 2 liters of O2 per minute and then at 2% isoflurane in 2 liters of O2 per minute to maintain. After anesthesia, the animals received intramuscular injections of buprenorphine (analgesic, 0.025 mg/kg) into the left thigh. After aseptic preparation for surgery, an approximately 40-mm-long skin incision was created over the right femur and underlying soft tissue was retracted to reveal the bone. Two transcortical holes were drilled in a lateromedial plane in the proximal femur, using a 0.9-mm drill bit (Gebr. Brasseler, Lemgo, Germany), and a second pair was placed in the distal femur, leaving enough space to create the defect. Four pins (1.1 mm, threaded Kirschner wires; MicroAire, Charlottesville, VA) were then placed into the femur and cut off to a length of approximately 20 mm. Two fixator plates were placed on top of each other and secured to the ends of the pins, using miniscrews. A 5-mm osteotomy was then created in the center of the bone between the two inner pins, using a sterile round dental burr (Gebr. Brasseler) attached to a dental hand piece (model 174025; Eickemeyer-Medizintechnik für Tierärzte KG, Tuttlingen, Germany). After completion of the osteotomy, the site was copiously irrigated with saline.
Implantation of tissue grafts
Excessive vector was washed off by placing the tissue grafts in a 50-ml Falcon tube (BD Biosciences, San Jose, CA) filled with PBS. PBS was replaced four times and the tube was shaken gently. The muscle grafts were then implanted within the segmental bone defect. A staple of five tissue fragments was “press fitted” into the defect without securing the implants to the periosteum. The muscle fasciae were then closed with 3-0 chromic catgut suture. The two fixator plates were lowered consecutively to a distance of about 5 mm above the surrounding muscle surface and secured to the pins. The pins were cut to be flush with the plates and the skin incision was closed over the plates, using 4-0 silk suture. Right after surgery, the animals received an intramuscular injection of buprenorphine (0.025 mg/kg) for additional analgesia. During the first 48 hr after surgery, each animal received intramuscular injections of buprenorphine (0.05mg/kg) every 12 hr.
Animals were killed 21 or 42 days after surgery according to their group affiliation. The femora were harvested and fixed in 4% ice-cold paraformaldehyde for 3 days (histology) or fresh frozen and stored at −20°C (X-ray, microcomputed tomography, and biomechanical testing).
Radiographic evaluation and microcomputed tomography
Femora were scanned with a desktop microtomographic imaging system (μCT 80; SCANCO Medical, Brüttisellen, Switzerland) equipped with a 10-mm focal spot microfocus X-ray tube. The scout view X-ray image was saved for the radiographic evaluation. The entire defect region was scanned using a 20-μm isotropic voxel size, at 70-keV energy, 400-msec integration time, and requiring approximately 420 μCT slices per specimen. In the defect region only a 3-mm bone volume was analyzed for each specimen to ensure that no old cortical bone was included in the analyses. Images were thresholded according to an adaptive–iterative algorithm (Meinel et al., 2005; Rajagopalan et al., 2005) and morphometric variables were computed from the binarized images, using direct, three-dimensional techniques that do not rely on any prior assumptions about the underlying structure (Hildebrand et al., 1999).
Biomechanical testing
After all noninvasive imaging, seven femora treated with BMP-2-activated muscle grafts and seven intact, contralateral femora were tested to failure in torsion on a biaxial testing machine (FastTrack 8874; Instron, High Wycombe, UK). Both ends of each specimen were embedded in polymethylmethacrylate (PMMA) (Technovit 3040; Heraeus Kulzer, Wehrheim, Germany) to provide an appropriate and reproducible gripping interface with the testing module. To avoid lateral forces and bending moments caused by clamping, a special manufactured device was used to place the specimen during the embedding process. This enabled us to embed the bone ends in the center of the PMMA disk and align the longitudinal axis of the bones with the torsion axis. Specimens were tested to failure rotation at a constant deformation rate of 5 rad/min angular deformation and applied load data were acquired at 10 Hz. Because of the potential shortening of the femora during torsion and the resulting tensile force along the longitudinal axis of the bones, a constant axial load of 5 N was applied during the test. The torsional stiffness was calculated from each torque rotation diagram. The torque to failure was defined as the first maximum in the torque rotation diagram.
Histology
The specimens were decalcified in 14% EDTA for up to 3 weeks. Pins were then removed from the bones before embedding and sectioning. Fixed and decalcified tissues were dehydrated in graded ethanol up to 100%, transferred to xylene, and embedded in paraffin. Five-micron-thick paraffin sections were placed on poly-
Statistical methods
Statistical significance of the biomechanical data and the bone volume data was determined by Mann–Whitney U test and Kruskal–Wallis test (p < 0.05 was considered statistically significant and p < 0.001 was considered statistically highly significant).
Results
Seven femora of the group treated with Ad.BMP-2 gene-activated muscle grafts and six of each control group were evaluated by radiographs and microcomputed tomography (μCT). Seven femora of the group treated with Ad.BMP-2 gene-activated muscle grafts and seven intact, contralateral femora underwent torsional testing. Two femora of each group served for histological investigations. Genetically modified muscle grafts were well tolerated and produced no obvious adverse effects during the 6 weeks of the experiment. All animals showed a normal eating pattern and no weight loss was recorded. Intra- and postoperatively, there were no complications and we did not see wound infections or delayed wound healing. The rats were able to bear their full weight on the operated limbs from the day of surgery until they were killed.
Measurement of BMP-2 upregulation by ELISA
On day 9 after transduction BMP-2 upregulation was confirmed by ELISA. BMP-2-transduced muscle disks produced BMP-2 at 5019 ± 3043 pg/ml. BMP-2 concentration in supernatants of controls was 42 ± 35 pg/ml (unmodified muscle) and 26 ± 36 pg/ml (GFP-transduced muscle).
Radiographic evaluation
Of the animals treated with Ad.BMP-2 gene-activated muscle grafts, 100% (seven of seven) showed united bones 6 weeks after treatment (Fig. 1A–C), whereas none of the control groups showed any bridging of the defects or noteworthy bone formation within the defects (Fig. 1D–L).
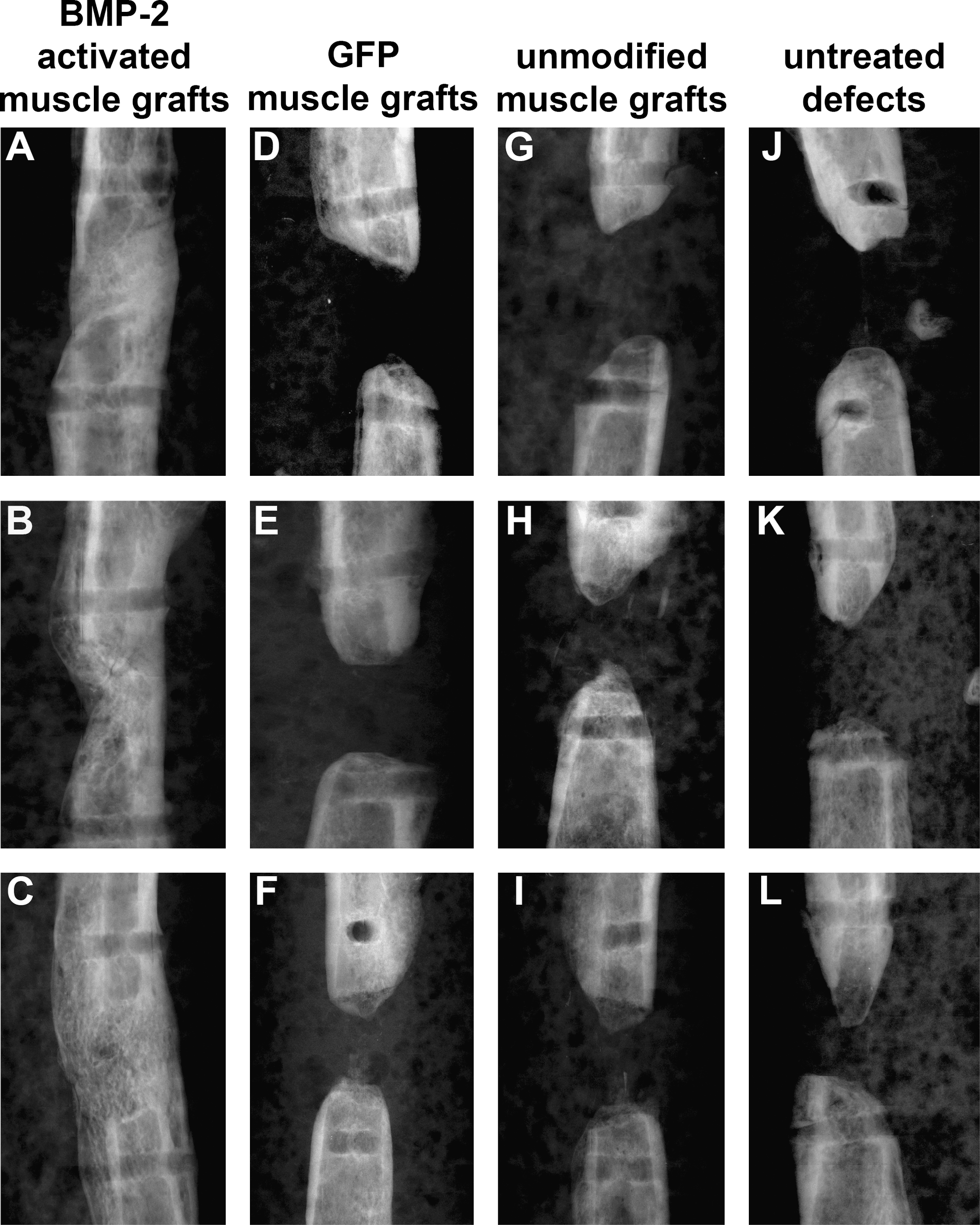
Radiographic images of femoral segmental bone defects of three representative specimens per group. (
Microcomputed tomography
Images
All seven femora receiving Ad.BMP-2 gene-activated muscle grafts displayed bridging of the 5-mm gaps at 6 weeks (Fig. 2A–C). None of the control animals showed bridging or relevant bone formation in the defect area (Fig. 2D–L).

Microcomputed tomography (μCT) images of femoral segmental bone defects of three representative specimens of each group. (
Bone volume
The bone volume of femora treated with BMP-2 gene-activated muscle tissue (25.78 ± 10.38 mm3) was significantly higher than that of the intact femora (16.94 ±0.51 mm3) (p = 0.006) and both groups displayed significantly higher bone volume than the control groups (p =0.002). There was no statistically significant difference among the three control groups: GFP-transduced muscle grafts (0.24 ± 0.16 mm3), untreated muscle grafts (0.49 ± 0.65 mm3), and untreated defects (0.29 ± 0.39 mm3) (p = 0.553) (Fig. 3).

μCT evaluation of the bone volume of corresponding areas of intact femora, defects treated with BMP-2-activated muscle grafts, and control groups. Values given represent means ± SD; (intact femora, n = 7; BMP-2-activated muscle grafts, n = 7; control groups, n = 6). *Statistically significant higher bone volume of intact bone compared with the control groups GFP-transduced muscle grafts, unmodified muscle grafts, and untreated defects (p = 0.002). **Statistically significant higher bone volume of femora receiving BMP-2-activated muscle grafts compared with intact bone (p = 0.006) and the control groups GFP-transduced muscle grafts, unmodified muscle grafts, and untreated defects (p = 0.002).
Biomechanical testing
The mechanical properties of femora treated with BMP-2-activated muscle grafts were evaluated by torsional testing and compared with those of intact femora. The mechanical properties of the three control groups could not be evaluated because of their insufficient structural integrity. Femora treated with BMP-2-activated muscle grafts showed a torque to failure (142.19 ± 91.64 N · mm) and a torsional stiffness (18.82 ± 13.44 N · mm/rad) in the range of intact bones (210.40 ± 63.24 N · mm and 30.19 ± 5.24 N · mm/rad, respectively) with no statistically significant difference (p =0.338 and p = 0.180, respectively) (Fig. 4A and B).

(
Histology
Micrographs of tissues within the defects of one specimen per group at 3 and 6 weeks postsurgery are presented in Figs. 5 and 6. The safranin O–fast green sections are presented to demonstrate the presence of cartilage, indicative of endochondral bone formation. Specimens with BMP-2-activated muscle grafts showed a large amount of trabecular bone within most of the defect at 3 weeks and some fibrous tissue in the middle of the defect area (Figs. 5A and 6A) and complete bridging of the defect at 6 weeks postsurgery (Figs. 5E and 6E). The formation of a neocortex, and at the same time a small amount of fibrous tissue, was seen within the defect. No presence of cartilage could be detected (Fig. 6E). None of the control groups showed bridging or bone formation within the defects (Fig. 5B–D and F–H, and Fig. 6B–D and F–H). Specimens that received a GFP muscle graft or an unmodified muscle graft showed still larger amounts of muscle tissue at 3 weeks (Fig. 5B and C, and Fig. 6B and C), but mostly fibrous tissue at 6 weeks postsurgery (Fig.5-A and G, and Fig. 6A and G).
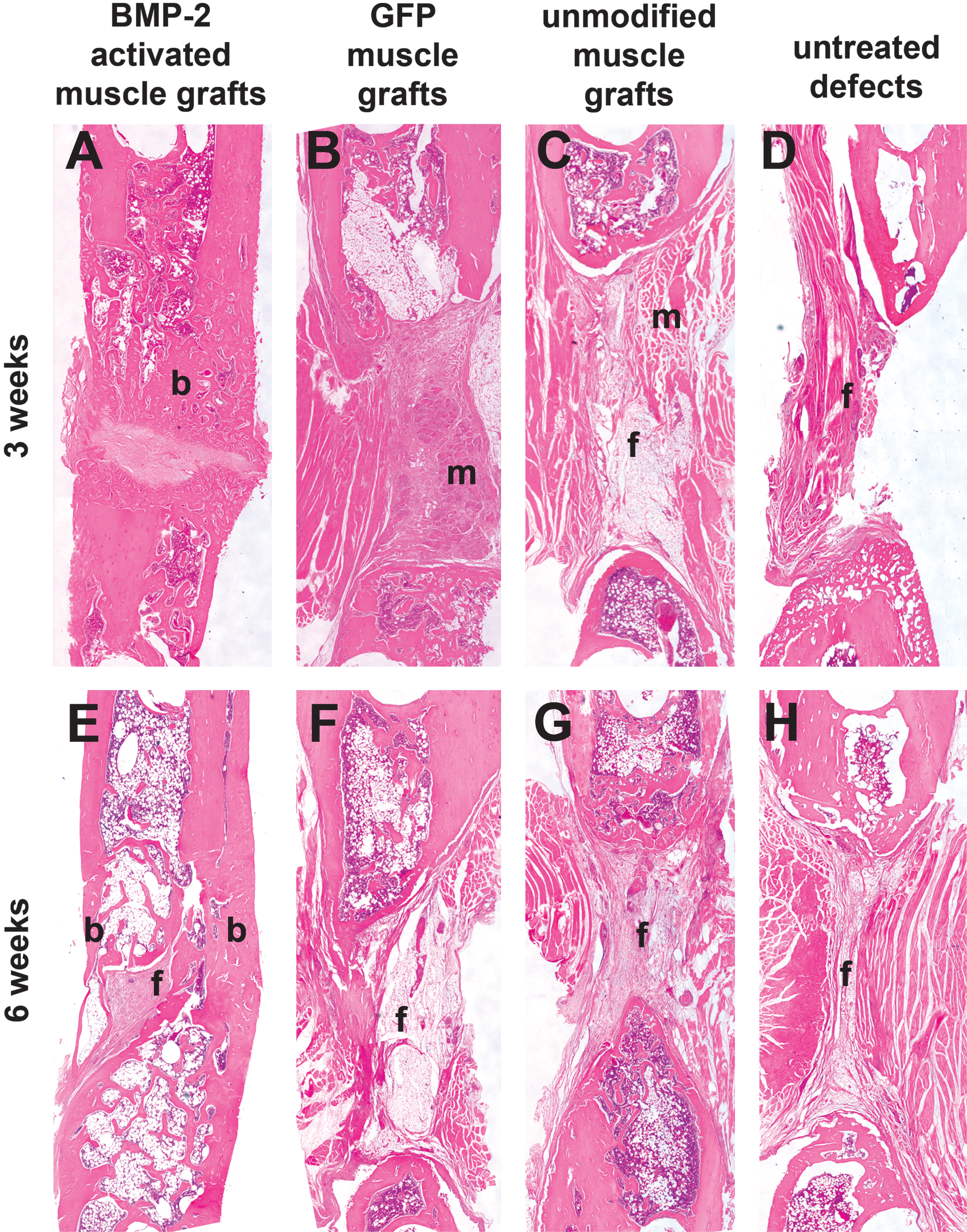
Histological sections of femoral segmental bone defects treated with BMP-2 gene-activated muscle grafts (

Histological sections of femoral segmental bone defects treated with BMP-2 gene-activated muscle grafts (
Discussion
Since the landmark work of Urist in 1965 it has been known that bone will form ectopically in skeletal muscle in response to bone morphogenetic proteins (Urist, 1965). In our experiment we exploited this property and were able to demonstrate that expedited BMP-2 gene-activated muscle grafts are osteoinductive, regenerative structures that have the ability to repair critical-size segmental bone defects in rats.
Bone formation was reliably induced after implantation of BMP-2-activated muscle tissue. All animals of this group had bridged defects at 6 weeks, as evaluated by radiographs and μCT. Histologically, cortical remodeling was visible at 6 weeks. Bone volumes of the femora repaired by BMP-2-transduced muscle were significantly higher compared with those of intact femora and the biomechanical stability was statistically indistinguishable. In contrast, defects of the three control groups did not heal. Only nonmineralized, fibrous tissue was seen in defects that remained untreated as well as in the groups that received unmodified or GFP-transduced muscle tissue. We have not yet investigated the underlying biological mechanisms after such heterotopic muscle transplantation but we assume that unmodified muscle was degraded and replaced by fibrous tissue because of the lack of vascularization.
These results are much more impressive than the results of our previous studies, in which we evaluated the effect of direct, adenoviral in vivo gene delivery to bone defects in the same animal model (O.B. Betz et al., 2006, 2007; V.M. Betz et al., 2007). The reason might be that, in the present study, we not only delivered an osteogenic molecule to the defect site but also provided a space-filling scaffold and a source of progenitor cells by implanting muscle tissue. Besides better effectiveness, we believe that another advantage over the direct, percutaneous injection of Ad.BMP-2 is that ex vivo gene therapy using gene-activated muscle grafts might be safer because free vector is not directly introduced into the body.
The advantage over conventional ex vivo approaches is that the expedited technology presented here does not require the isolation and expansion of autologous progenitor cells, which makes this method less cumbersome, less time-consuming, and less expensive. Instead of culturing cells over a period of 1 to 3 weeks as required for the expansion of progenitor cells, we kept the muscle grafts in culture for only 24 hr. Bearing in mind that cost-effectiveness is a crucial criterion for tissue-engineering technologies to enter widespread clinical use (Evans et al., 2007), we believe that this novel bone repair strategy may have potential for translation into a clinical setting.
Muscle flaps are frequently used in reconstructive surgery for the reconstruction of soft tissue defects. In clinical studies, donor site morbidity after the harvest of the vastus lateralis muscle or the gracilis muscle was minimal and the complete absence of these muscles rarely caused lower extremity weakness (Deutinger et al., 1995; Chen et al., 1999). The amount of muscle tissue that is available in humans by harvesting gracilis or vastus lateralis muscle would be enough to fill large, critical-size defects. We are planning to evaluate this novel approach in a lager animal model, using autograft tissue.
More than 360 human clinical gene therapy trials involving adenoviral vectors have already been conducted. One death occurred in 1999 when a high dose of adenovirus was infused into the hepatic portal vein of a subject with ornithine transcarbamylase deficiency. The death was caused by an uncontrollable systemic reaction of the immune system leading to respiratory failure (Stolberg, 1999; Somia and Verma, 2000). Other clinical trials suggest that administration of adenoviral vectors does not necessarily provoke serious immune responses, especially when the vector is delivered locally (Yen et al., 2000; Nemunaitis et al., 2006). This and the fact that BMP-2-activated muscle grafts were well tolerated by all animals in this study makes us believe that regional, ex vivo gene therapy, as envisioned here, could be safe for human application. However, detailed toxicological and histological long-term evaluations must be performed to obtain a definite answer regarding the safety of genetically enhanced bone tissue engineering.
There is great interest in identifying expedited, cost-effective bone repair strategies (Evans et al., 2007). In this preclinical study we have shown that expedited implantation of muscle fragments transduced with an adenoviral BMP-2 vector reliably induces bone formation in rodents. If this abbreviated ex vivo gene therapy approach proves to be safe and equally effective in humans, it could provide a clinically expeditious technology for the regeneration of critical-size bone defects.
Footnotes
Acknowledgment
This work was supported by funds of the Department of Orthopedic Surgery, Klinikum Grosshadern, Ludwig-Maximilians-University Munich.
Author Disclosure Statement
No competing financial interests exist.