
Abstract
Type 1 diabetes (T1D), characterized by permanent destruction of insulin-producing beta cells, is lethal unless conventional exogenous insulin therapy or whole-organ transplantation is employed. Although pancreatic islet transplantation is a safer and less invasive method compared with whole-organ transplant surgery, its treatment efficacy has been limited by islet graft malfunction and graft failure. Thus, ex vivo genetic engineering of beta cells is necessary to prolong islet graft survival. For this reason, a novel gene therapy approach involving adenovirus-mediated TRAIL gene delivery into pancreatic islets was tested to determine whether this approach would defy autoreactive T cell assault in streptozotocin (STZ)-induced diabetic rats. To test this, genetically modified rat pancreatic islets were transplanted under the kidney capsule of STZ-induced diabetic rats, and diabetic status (blood sugar and body weight) was monitored after islet transplantation. STZ-induced diabetic rats carrying Ad5hTRAIL-infected islets experienced prolonged normoglycemia compared with animals grafted with mock-infected or AdCMVLacZ-infected islets. In addition, severe insulitis was detected in animals transplanted with mock-infected or AdCMVLacZ-infected islets, whereas the severity of insulitis was reduced in animals engrafted with Ad5hTRAIL-infected islets. Thus, TRAIL overexpression in pancreatic islets extends allograft survival and function, leading to a therapeutic benefit in STZ-induced diabetic rats.
Introduction
The type II membrane protein TNF (tumor necrosis factor)-related apoptosis-inducing ligand (TRAIL) is a member of the TNF family (Wiley et al., 1995) that can interact with four different membrane-bound receptors: TRAIL-R1 (DR4), TRAIL-R2 (DR5), TRAIL-R3 (DcR1), and TRAIL-R4 (DcR2) (Griffith and Lynch, 1998; Griffith et al., 1999). DR4 and DR5 function as authentic death receptors that signal for apoptosis, whereas DcR1 and DcR2 are unable to transduce such signaling because they lack the intracellular death domain necessary to engage the apoptosis signaling pathway (Meng et al., 2000; Karacay et al., 2004; Sanlioglu et al., 2006). Although a significant amount of information is available about TRAIL as an anticancer agent (Thorburn et al., 2008; Wang, 2008), its functional role in autoimmune diabetes remains to be clarified (Lorz et al., 2008). The potential role of TRAIL in T1D has been studied in two animal models of autoimmune diabetes (Lamhamedi-Cherradi et al., 2003). TRAIL function was blocked in one model through the systemic administration of a soluble TRAIL receptor into nonobese diabetic (NOD) mice. The onset of diabetes was significantly increased and the degree of autoimmune inflammation was augmented in pancreatic islets as a result of TRAIL blockage. In the second model, both normal and TRAIL-deficient C57BL/6 mice received multiple low doses of streptozotocin (STZ). TRAIL-deficient animals displayed a higher degree of islet inflammation leading to an earlier onset of diabetes. These results imply that TRAIL expression is required for the downregulation of autoimmune inflammatory response in T1D. Additional studies examining TRAIL receptor expression on human beta cell lines and normal primary islet cells found the expression of all four TRAIL receptors and/or TRAIL on the beta cell lines (CM and HP62) (Ou et al., 2002). Interestingly, the beta cell lines were sensitive to TRAIL-induced apoptosis, but normal primary islet cells displayed resistance to TRAIL-induced cytotoxicity (Ou et al., 2005). The reason for this discrepancy remains to be determined.
Interestingly, there are several reports implicating TRAIL as an apoptosis-inducing agent in human pancreas. Freshly isolated T cells did not express TRAIL unless they were treated with type I interferon or by CD3 ligation (Kayagaki et al., 1999). Accordingly, patients with T1D exhibit increased TRAIL expression on the infiltrating cells of the pancreatic islets (Cheung et al., 2005). In addition, two studies from our group have linked high TRAIL expression to increased cell death in human pancreas (Sanlioglu et al., 2008a, 2009). The fact that interleukin (IL)-2-stimulated T cells are sensitive to TRAIL-mediated apoptosis suggests a role for TRAIL in the peripheral deletion of T cells (Ashkenazi and Dixit, 1999). With these data in mind, the following studies were conducted to investigate the hypothesis that TRAIL overexpression in beta islet cells would be protective against autoreactive cytotoxic T cell (CTL) assault and result in prolonging survival of transplanted islets in STZ-induced diabetic rats.
Materials and Methods
Production of recombinant adenoviral vectors
Amplification of the vectors Ad5hTRAIL (Griffith et al., 2000), AdEGFP (Yang et al., 2003; Sanlioglu et al., 2004), and AdCMVLacZ (Doerschug et al., 2002) was performed as previously described (Engelhardt et al., 1993). Briefly, adenoviral vectors were amplified by infection of HEK-293 cells (Kahraman et al., 2009). After reaching appropriate levels of cytopathologic effect (CPE), the medium was collected and cells were lysed by freezing and thawing. Cell debris was pelleted, and viral supernatant was saved for subsequent experiments. Adenovirus was purified on a CsCl–Tris gradient, separated into aliquots, and stored at −80°C in 10 mM Tris with 20% glycerol until use. The particle titers of adenoviral stocks were ∼1013 DNA particles/ml, and the typical particle/plaque-forming unit ratio was 50:1.
Isolation and purification of rat pancreatic islets
Outbred female Wistar rats (8–10 weeks old; body weight, 250–300 g) were obtained from the Animal Laboratory Unit of Akdeniz University (Antalya, Turkey). All animals were housed under conventional conditions (food and water were provided ad libitum), and all studies were performed in accordance with guidelines established by the Helsinki Declaration and with the approval of the Akdeniz University Institutional Animal Care and Use Committee. Islets were isolated as previously described, with some modifications (Lacy and Kostianovsky, 1967). Briefly, after intraperitoneal ketamine–xylazine anesthesia, a midline abdominal incision was made, and the pancreas was exposed and injected with 10 ml of Liberase RI (0.25 mg/ml; Roche, Indianapolis, IN) in serum-free Dulbecco's modified Eagle's medium (DMEM; Sigma-Aldrich, St. Louis, MO) through the pancreatic duct. The pancreas was surgically removed and incubated at 37°C for 23 min. After incubation, 40 ml of ice-cold DMEM with 10% newborn calf serum (Sigma-Aldrich) was added to stop the digestion and the tissue was passed through a 425-μm wire mesh (Thomas Scientific, Swedesboro, NJ). The digested pancreas was rinsed with DMEM, and islets were separated from the exocrine tissue by centrifugation through a Histopaque-1077 density gradient (Sigma-Aldrich). After several washes with DMEM, islets were hand-picked under a microscope. To standardize the islet mass to be transplanted, 1 islet equivalent (IE) was defined as one 150-μm-diameter islet. Islet yield was calculated by taking samples of 100 μl from the final preparation. Before transplantation, islets were placed in culture overnight in RPMI 1640 medium (Sigma-Aldrich) supplemented with 10% newborn calf serum at 37°C and 5% CO2.
Gene transfer experiments
Freshly isolated islets were cultured with Ad5hTRAIL, AdCMVLacZ, or AdEGFP in serum-free RPMI 1640 for 2 hr at 37°C. Fetal bovine serum (FBS) was added 2 hr later to reach a final concentration of 10% and islets were further incubated at 37°C for 30–36 hr before transplantation. Islet morphology was assessed daily under an inverted microscope.
Fluorometric measurements of AdEGFP-transduced rat pancreatic islets
Freshly isolated rat pancreatic islets were maintained in RPMI 1640 medium without serum in 96-well plates. Islets were infected with increasing doses of AdEGFP vectors (multiplicity of infection [MOI] of 10, 50, 200, and 1000) 2 hr postisolation. FBS was added 2 hr later to achieve a final concentration of 10%. After 48 hr, fluorescence microscopy (IX81 motorized inverted microscope; Olympus, Planegg, Germany) was used to monitor and quantify cell infection [using a fluorometer [Fluoroskan Ascent FL, Thermo Fisher Scientific, Waltham, MA]).
Immunohistochemical analysis of TRAIL overexpression after Ad5hTRAIL infection of isolated pancreatic islets
Immunohistochemical analysis was performed as described previously (Sanlioglu et al., 2007a,b). Overexpression of hTRAIL on isolated islets was achieved by Ad5hTRAIL infection, and AdCMVLacZ was used as a control vector in the assay. Briefly; Ad5hTRAIL- or AdCMVLacZ-infected and uninfected islets were spun onto positively charged slides (Shandon Cytospin 4 cytocentrifuge; Thermo Fisher Scientific). After the Cytospin procedure, the slides were left to air dry for 5 min before fixing in 1:1 ice-cold methanol–acetone for 8 min at −20°C and washed twice in phosphate-buffered saline (PBS) for 15 min. To prevent nonspecific staining of endogen peroxidase, slides were treated with H2O2 (ScyTek Laboratories, Logan, UT) and Ultra V Block (Lab Vision/Thermo Fisher Scientific, Fremont, CA). Samples were stained with mouse anti-human TRAIL monoclonal antibody (mAb) (diluted 1:200; Alexis Biochemicals, San Diego, CA) for 1 hr, followed by incubation with biotin-labeled goat anti-polyvalent antibody (Lab Vision/Thermo Fisher Scientific) for 20 min. Streptavidin peroxidase (TS-125-HR; Lab Vision/Thermo Fisher Scientific) was added for 20 min. After washing twice with PBS, slides were incubated with chromogen substrate 3,3′-diaminobenzidine tetrahydrochloride solution (DAB; Lab Vision/Thermo Fisher Scientific). Counterstaining was performed by hematoxylin–eosin (H&E) staining. Rabbit anti-human TRAIL antibody (ProSci, Poway, CA) was used for Western blotting.
Glucose-stimulated insulin secretion in isolated rat islets
Freshly isolated islets were infected with AdCMVLacZ or Ad5hTRAIL vector (MOI of 1000) for 36 hr as described previously. Adenovirus-infected and mock-infected islets were washed with PBS and then cultured in low (2.8 mM) and then high (16.7 mM) concentrations of glucose. The static incubation assay was performed in a 24-well plate with 200 islets/0.5 ml per well. Supernatant from each well was collected after 1 hr of incubation. Insulin concentrations were determined by indirect enzyme-linked immunosorbent assay (ELISA). Mouse anti-rat insulin antibody (Abcam, Cambridge, UK) and horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG (Bio-Rad, Hemel Hempstead, UK) were used as primary and secondary antibodies, respectively. Optical density at 450 nm (OD450) was determined 20 min after tetramethyl benzidine (TMB) substrate addition.
Apoptosis detection by annexin V–FITC binding
Freshly isolated islets were infected either with Ad5hTRAIL or AdCMVLacZ vector at the indicated MOI. An annexin V–FITC (fluorescein isothiocyanate) staining procedure was employed to measure apoptosis according to manufacturer's instructions (BD Biosciences, San Jose, CA). As a positive control, a group of islets was treated with 1 mM H2O2 for 1 hr to induce apoptosis. Results were analyzed by fluorescence microscopy.
Generation of STZ-induced type 1 diabetes in Wistar rats
Outbred female Wistar rats were rendered diabetic by a single intraperitoneal injection of streptozotocin (STZ) (Sigma-Aldrich,) that had been freshly dissolved in 0.1 M citrate buffer (pH 4.5). Diabetes was confirmed before transplantation by measuring blood glucose levels and only those rats with blood glucose levels of 300 mg/dl or more were used as recipients. Tail blood was taken from nonfasting rats and whole glucose concentrations were measured with a portable glucose meter (Accu-Chek Go; Roche, Nutley, NJ) at the indicated time points.
Islet transplantation procedure
Groups of 750 Ad5hTRAIL-, AdCMVLacZ-, or mock-infected islets were transplanted into STZ-induced diabetic animals. Islets were aspirated into a 200-μl pipette tip connected with a 0.5-ml syringe (Hamilton, Reno, NV) and then transferred into polyethylene tubing P-50 (Becton Dickinson, Franklin Lakes, NJ). The left kidney of the recipient rat under intraperitoneal ketamine–xylazine anesthesia was exposed through a lumbar incision. A capsulotomy was performed on the caudal outer face of the kidney and the tip of the tube was advanced under the capsule. The tubing was then removed and the capsulotomy was cauterized with a low-temperature cautery pen. The kidney was replaced in the abdominal cavity, and the incision was sutured. The engraftment was considered successful when blood glucose levels dropped below 200 mg/dl after transplantation. The animals were killed after confirmation of diabetes recurrence and then kidney with the transplant was recovered for histological analysis.
Immunohistochemistry and histology of transplanted pancreatic islet allografts
Graft-bearing kidneys were harvested 15, 30, 60, and 90 days after transplantation and fixed in 10% buffered formalin, embedded in paraffin, sectioned (thickness, 5 μm), and placed on poly-
Results
Quality assessment of purified rat pancreatic islets
The successful transplantation of any solid organ is largely dependent on the quality of the donor tissue. Thus, our initial investigation into pancreatic islet transplantation was centered on modifying a previously reported rat pancreatic islet isolation procedure to obtain higher islet yield and quality. We achieved the highest islet recovery rate after occlusion of either side of the small intestine, where the common bile duct drains, leaving a small pocket for infusing Liberase RI solution. Analysis of the islets for cell viability and purity indicated that the freshly isolated and hand-picked islets appeared in regular shapes, and had a size range of 50–500 μm in diameter, with the majority being 100–200 μm (Fig. 1). The purity of the hand-picked islets was approximately 90%, as revealed after dithizone (DTZ) staining. Propidium iodide (PI)–fluorescein diacetate (FDA) staining demonstrated that almost all cells of isolated islets were alive, except for some occasional apoptotic cells within the islet cell cluster. Together, these data show that complete enzyme dispersion in pancreas preisolation leads to successful islet isolation with high quality and yield.
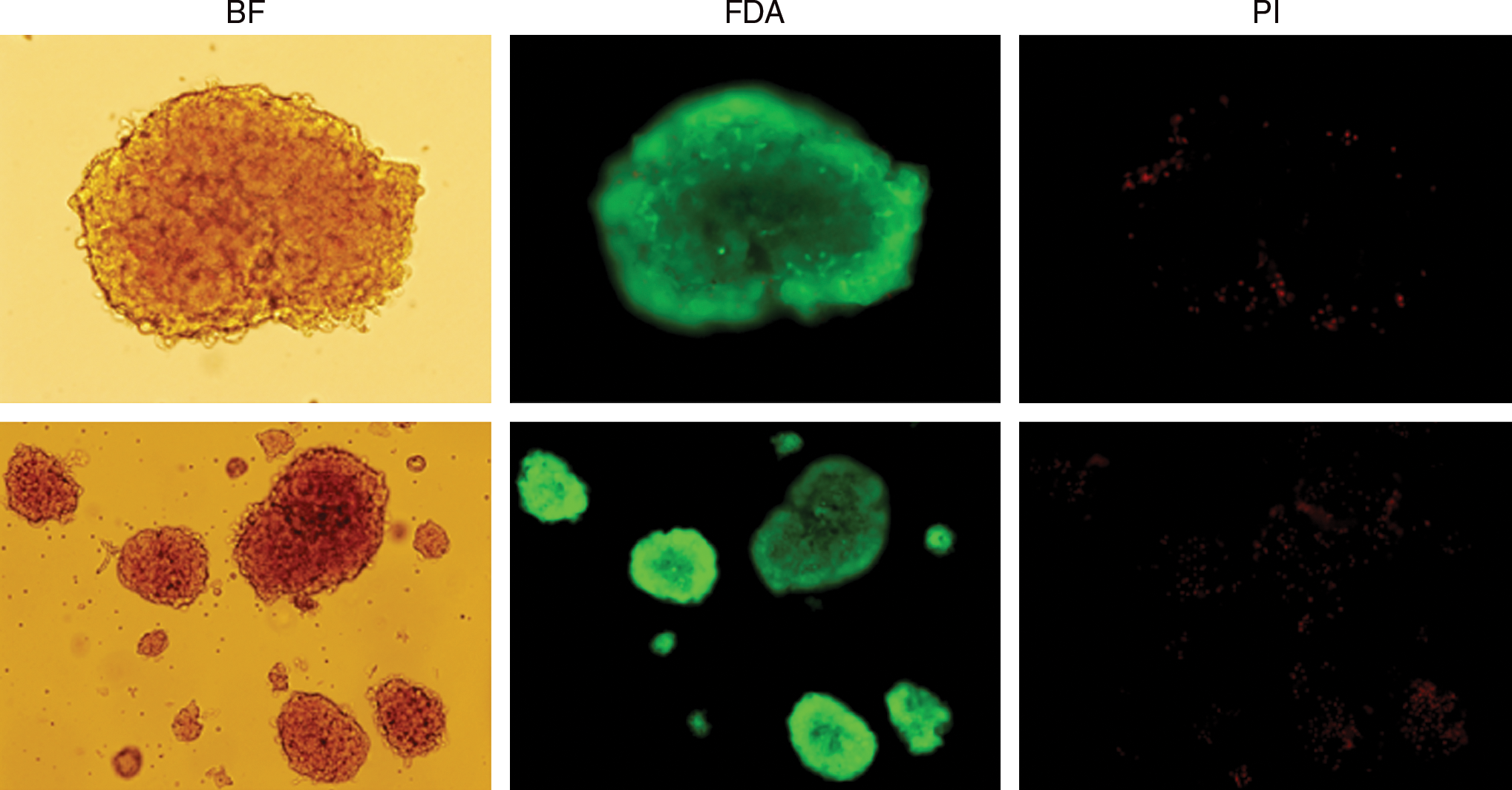
Cell viability of isolated pancreatic islets. Propidium iodide (PI)–fluorescein diacetate (FDA) staining was performed on isolated rat pancreatic islets (original magnification: top lane, × 400; bottom lane, × 200). Bright-field (left), FDA-stained (middle), and PI-stained (right) islets are shown by fluorescence microscopy. Color images available online at
Adenovirus-mediated transduction of pancreatic islets
We next determined the efficacy of transduction of isolated rat pancreatic islets by first-generation adenoviral vectors. Freshly isolated islet cells were infected with increasing MOIs of adenovirus encoding enhanced green fluorescent protein (AdEGFP). The transduction efficiency was determined by fluorescence microscopy (Fig. 2A) and then analyzed by fluorometry 48 hr after infection (Fig. 2B). No EGFP signal was detected in mock-infected islet samples, but AdEGFP-infected samples expressed significant EGFP on the basis of the viral dose used. Whereas an MOI of 200 DNA particles/cell transduced a measureable number of islets, optimal transduction efficiency was attained with an MOI of 1000 DNA particles/cell. Together, these results demonstrate that pancreatic islets can be successfully transduced with recombinant adenoviral vectors, and the number of EGFP-expressing islets was proportional to the viral dose used.

AdEGFP transduction of rat pancreatic islets. Freshly isolated rat pancreatic islets were infected with AdEGFP (MOIs of 0, 10, 50, 200, and 1000) virus as described in Materials and Methods. (

On the basis of the preceding data, we next transduced rat pancreatic islets with Ad5hTRAIL at an MOI of either 200 or 1000 for 48 hr to achieve TRAIL overexpression. Subsequently, TRAIL transgene expression was evaluated by immunohistochemistry and Western Blotting. As shown in Fig. 3A and B, neither mock-infected nor AdCMVLacZ-infected islets displayed measurable TRAIL expression. However, infection with Ad5hTRAIL resulted in significant TRAIL production in rat pancreatic islets, which was proportional to the vector dose used. Importantly, there was no observable difference in glucose-stimulated insulin release between mock-infected or adenovirus-infected (AdCMVLacZ/Ad5hTRAIL) pancreatic islets (Fig. 3C). Collectively, these data suggest that neither adenovirus infection nor transgene expression interferes with glucose-stimulated insulin release from pancreatic islets.

Immunocytochemical analysis of TRAIL expression on rat pancreatic islets. (
Assessment of islet cell death after adenoviral infection
TRAIL is well known for its cytotoxic activity against tumor cells, whereas normal cells/tissues are unaffected. Thus, we next evaluated the viability of islet cells after forced TRAIL expression. Isolated pancreatic islets were infected with either AdCMVLacZ or Ad5hTRAIL at various MOIs for 48 hr before measuring apoptosis by annexin V–FITC staining (Fig. 4). Neither AdCMVLacZ nor Ad5hTRAIL-infected islets displayed the cell surface markers of apoptosis at an MOI of 1000 DNA particles/cell. However, increasing the MOI to 5000 DNA particles/cell induced significant apoptosis for both AdCMVLacZ- and Ad5hTRAIL-infected samples. This finding suggests that apoptosis induction was not due to the transgene expressed (TRAIL or LacZ), but rather was due to adenoviral infection because both vectors generated the same result.
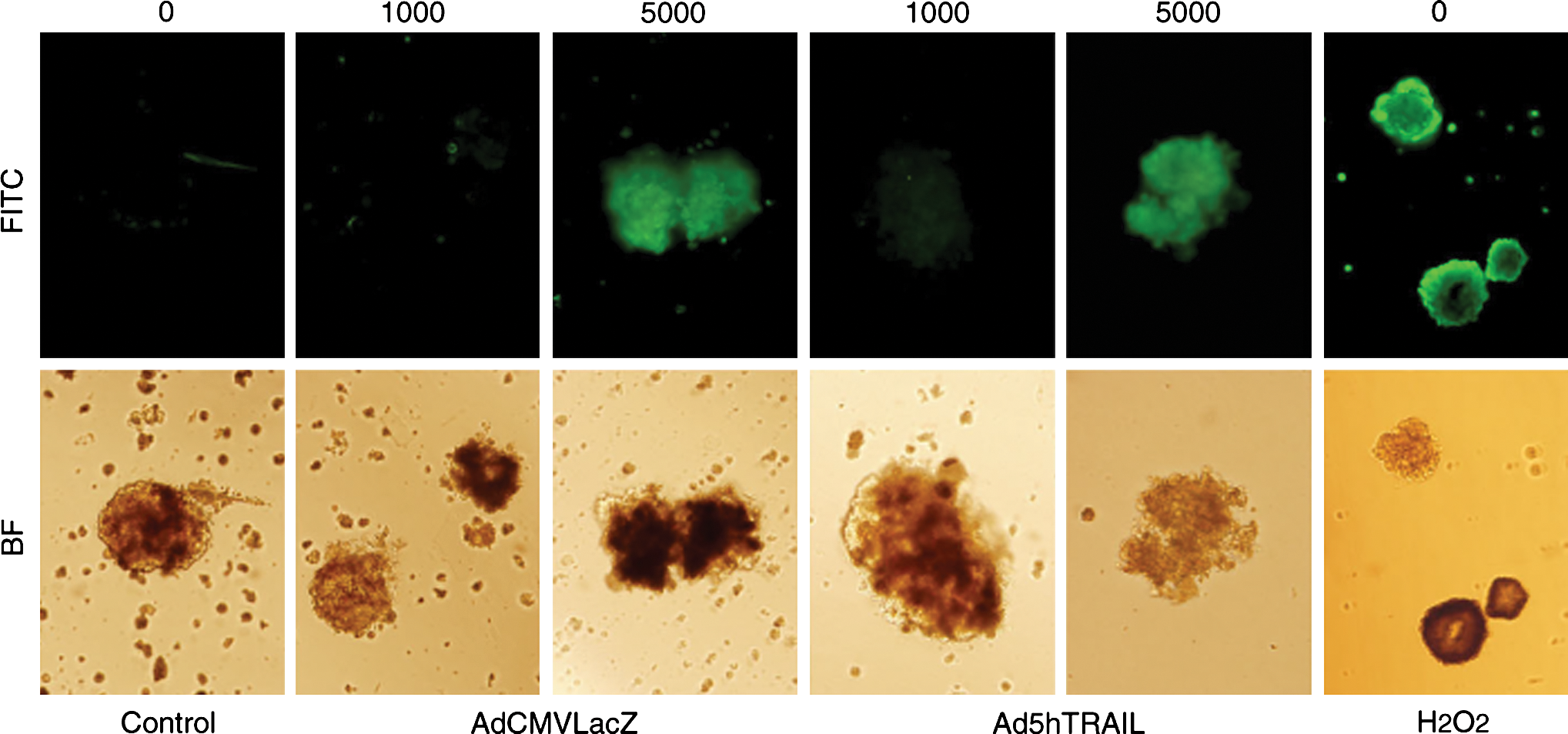
Cytotoxic effects of adenoviral infection on pancreatic islets. Mock-transduced, AdCMVLacZ-transduced, or Ad5hTRAIL-transduced islets (MOIs of 1000 and 5000) were stained with annexin V–FITC 48 hr after infection and then analyzed by fluorescence microscopy. As a positive control, islets were treated with 1 mM H2O2 for 1 hr. Images were taken at an original magnification of × 200. Color images available online at
Generation of STZ-induced type 1 diabetes in rats
Diabetogenic agents such as STZ, which selectively destroy pancreatic beta cells, have been widely used to experimentally induce T1D in rats. To find the optimal dose of STZ to induce T1D, a single intraperitoneal injection of STZ at 30, 40, or 50 mg/kg was administered to three groups of rats and these animals were monitored for 100 days for survival (Fig. 5A). Survival decreased to 16.6% within 4 days of STZ administration with an intraperitoneal dose of 50 mg/kg and therefore this group was terminated on day 35, because of the unfavorable effects of STZ. The survival rate of animals that received STZ at 40 mg/kg decreased to 70% within 4 days of STZ administration, but no additional deaths occurred during the rest of the assessment period. The best survival rate (91%), however, was observed in the animals that received STZ at 30 mg/kg.
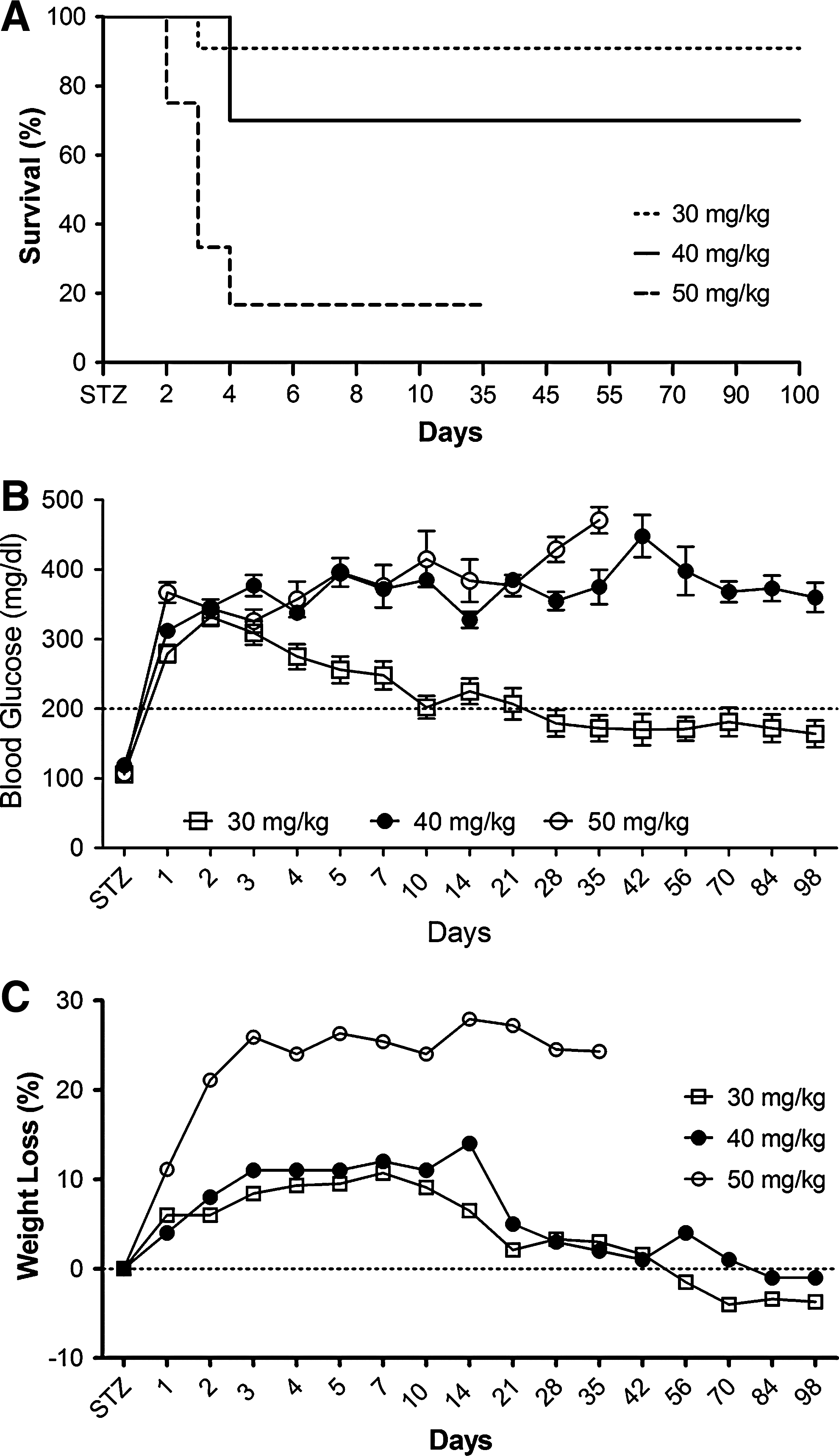
Streptozotocin (STZ) induction of type 1 diabetes (T1D) in rats. (
Along with survival rates, we simultaneously monitored blood glucose levels. As shown in Fig. 5B, all three groups displayed blood glucose levels above 200 mg/dl 24 hr after STZ injection. Whereas the blood glucose level remained elevated during the entire assessment period in the rats that received STZ at either 40 or 50 mg/kg, those animals that received STZ at 30 mg/kg began to show a decrease in blood glucose levels on day 3 and displayed normoglycemia by day 10. One-way analysis of variance (ANOVA) followed by Bonferroni's multiple comparison test indicated that although no significant difference in blood glucose levels was observed in animals injected with STZ at either 40 or 50 mg/kg, animals injected with STZ at 30 mg/kg displayed a significant reversion in blood glucose compared with the rest of the groups (p < 0.001). Although the blood glucose levels of this group never returned to original levels and stayed slightly below 200 mg/dl (our criterion for hyperglycemia) during the entire follow-up period, we considered these animals to be normoglycemic. We also measured the body weight of the three groups of animals (Fig. 5C). Animal injected with STZ at 30 and 40 mg/kg showed nearly 10% body weight loss until day 10. Whereas the 30-mg/kg STZ group started to gain weight, the 40-mg/kg STZ group continued to lose weight until day 14. Despite this difference, both groups eventually returned to preinjection body weights. In contrast, a 25% decrease in body weight was observed in the animals that received STZ at 50 mg/kg within the first 3 days of STZ administration. Although no additional weight loss occurred in this group, the weight loss was permanent. One-way ANOVA followed by Bonferroni's multiple comparison test demonstrated that body weight loss was statistically significant in 50-mg/kg STZ-injected animals compared with rest of the groups (p < 0.0001), but the difference in weight loss was not significant in animals injected with STZ at either 30 or 40 mg/kg. Together, when we considered survival rates, blood glucose levels, and morbidity, we decided that 40-mg/kg STZ was the optimal dose to induce T1D in rats for our subsequent experiments.
Ad5hTRAIL-transduced pancreatic islets prolong normoglycemia in STZ-induced diabetic rats
On the basis of our previous experience, a minimum of 750 islet equivalents (IE) was required to restore normoglycemia in STZ-induced diabetic rats. For this reason, 750 IE was used in our transplantation experiments. Rat pancreatic islets were infected with either AdCMVLacZ or Ad5hTRAIL (MOI of 1000) for 48 hr, and then transplanted under the left kidney capsule of STZ-induced diabetic outbred Wistar rats (blood glucose, ≥300 mg/dl). After transplantation, blood glucose levels were monitored daily for the first 10 days and then at 1-week intervals thereafter. Although normoglycemia was achieved 14 days after transplantation when using mock-infected islets, this protection was short-lived and the animals became hyperglycemic 32 days later (Fig. 6). Diabetic animals transplanted with pancreatic islets infected with AdCMVLacZ did not regain normoglycemia during the entire follow-up period. In contrast, diabetic animals transplanted with Ad5hTRAIL-infected islets regained normoglycemia 7 days after transplantation and this status was maintained for 60 days. One-way ANOVA followed by Bonferroni's multiple comparison test revealed that Ad5hTRAIL-infected islet grafts lowered blood glucose significantly compared with that of AdCMVLacZ-infected islets (p < 0.01). However, the decline in blood glucose was not significant between TRAIL-expressing islets and mock-infected islets (p > 0.05). In addition, no significant difference was noted in survival rates and body weight among the three groups. To confirm that normoglycemia was dependent on the function of the transplanted islets, rats were nephrectomized on day 32 after transplantation and became hyperglycemic (blood glucose, ≥300 mg/dl; data not shown) within 24 hr. In summary, Ad5hTRAIL-infected islets indisputably prolonged normoglycemia compared with mock-infected islets.
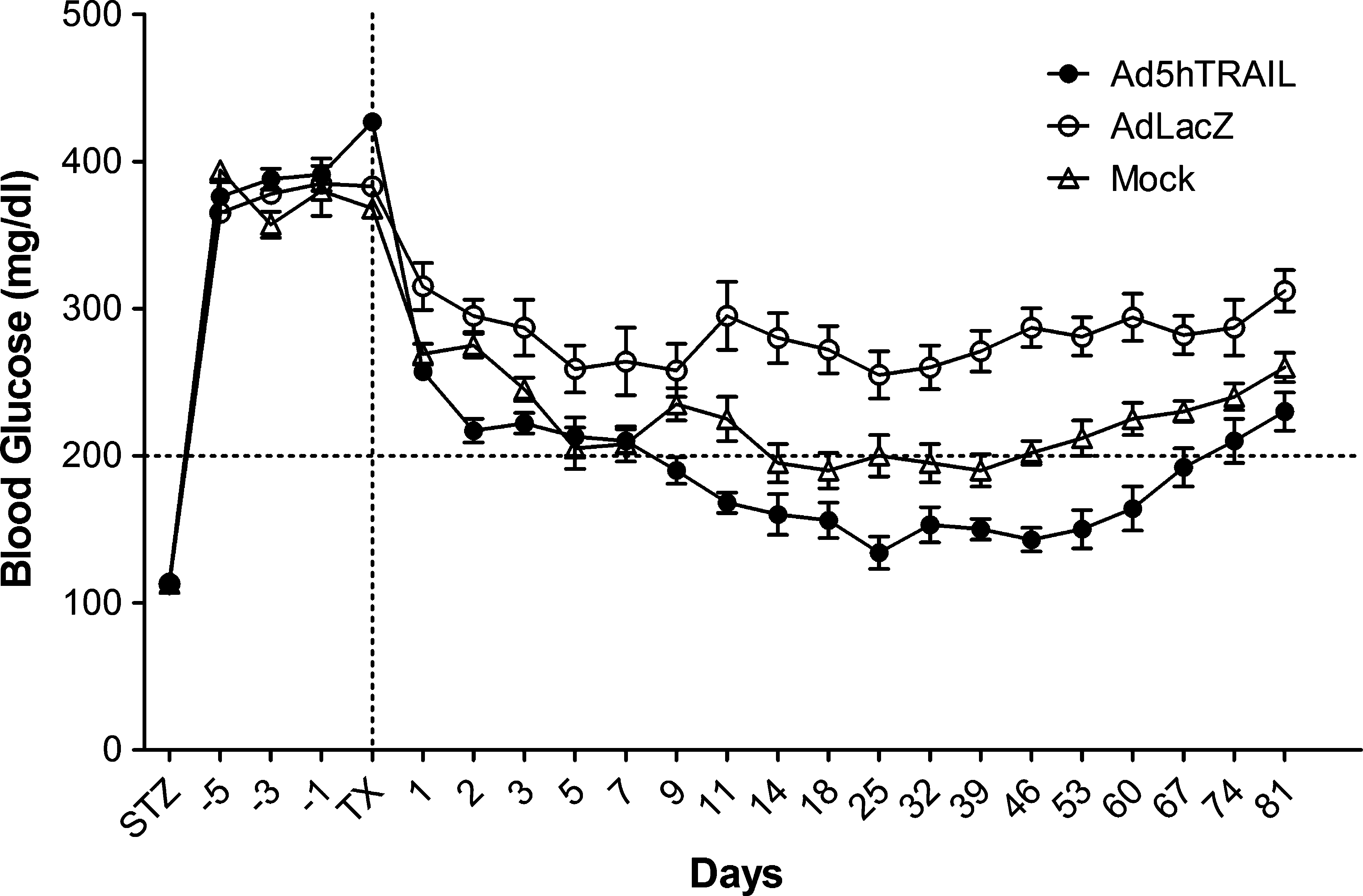
Therapeutic outcome of diabetic rats given genetically engineered pancreatic islet transplants. Diabetic rats were grafted with AdCMVLacZ (open circles; n = 10), Ad5hTRAIL (solid circles; n = 10), or mock-transduced (open triangles; n = 10) at 750 islet equivalents (IE) 7 days after STZ injection. The dotted line on the x axis represents the transplantation time, and the dotted line on the y axis displays normoglycemic animals (defined as blood glucose levels ≤200 mg/dl on two consecutive readings). Data represent means ± SEM of at least two different experiments.
Exogenous TRAIL expression reduces mononuclear cell infiltrates after transplantation of genetically engineered islets into STZ-induced diabetic rats
Because Ad5hTRAIL-infected pancreatic islet allografts prolonged normoglycemia in STZ-induced diabetic rats, we were interested in examining the degree of inflammation in the transplant. Thus, kidneys from STZ-induced diabetic rats were harvested at various time points after the transfer of adenovirus-infected islets. As shown in Fig. 7, mononuclear cell infiltration was first observed on day 15 after the transplantation of AdCMVLacZ-infected or mock-infected islets. Significant mononuclear cell infiltrates were detected after day 30 in these two groups of animals. Interestingly, mononuclear cells appeared as dense aggregates inside the islet allografts in the mock-infected and AdCMVLacZ-infected islets. By comparison, there was no detectable cell infiltrate on day 15 in animals transplanted with Ad5hTRAIL-infected islet allografts, and diabetic rats transplanted with Ad5hTRAIL-infected islets displayed reduced mononuclear cell infiltrates throughout the assay period. We also quantified the inflammation in each group, and the animals that received Ad5hTRAIL-infected islets had significantly lower insulitis compared with those that received either AdCMVLacZ- or mock-infected islets (Fig. 8). To disclose the insulin profile of transplanted islets, the sections used to quantify inflammation were immunostained for insulin expression. As shown in Fig. 9, sustained insulin expression was achieved only in animals transplanted with Ad5hTRAIL-infected islets 90 days after transplantation. Intriguingly, low levels of TRAIL expression were still detectable in these islets (Fig. 10). Although animals transplanted with Ad5hTRAIL-infected islets displayed low blood glucose levels compared with those transplanted with AdCMVLacZ-infected islets on day 90, neither TRAIL nor insulin expression was sufficient to maintain normoglycemia in STZ-induced diabetic rats.
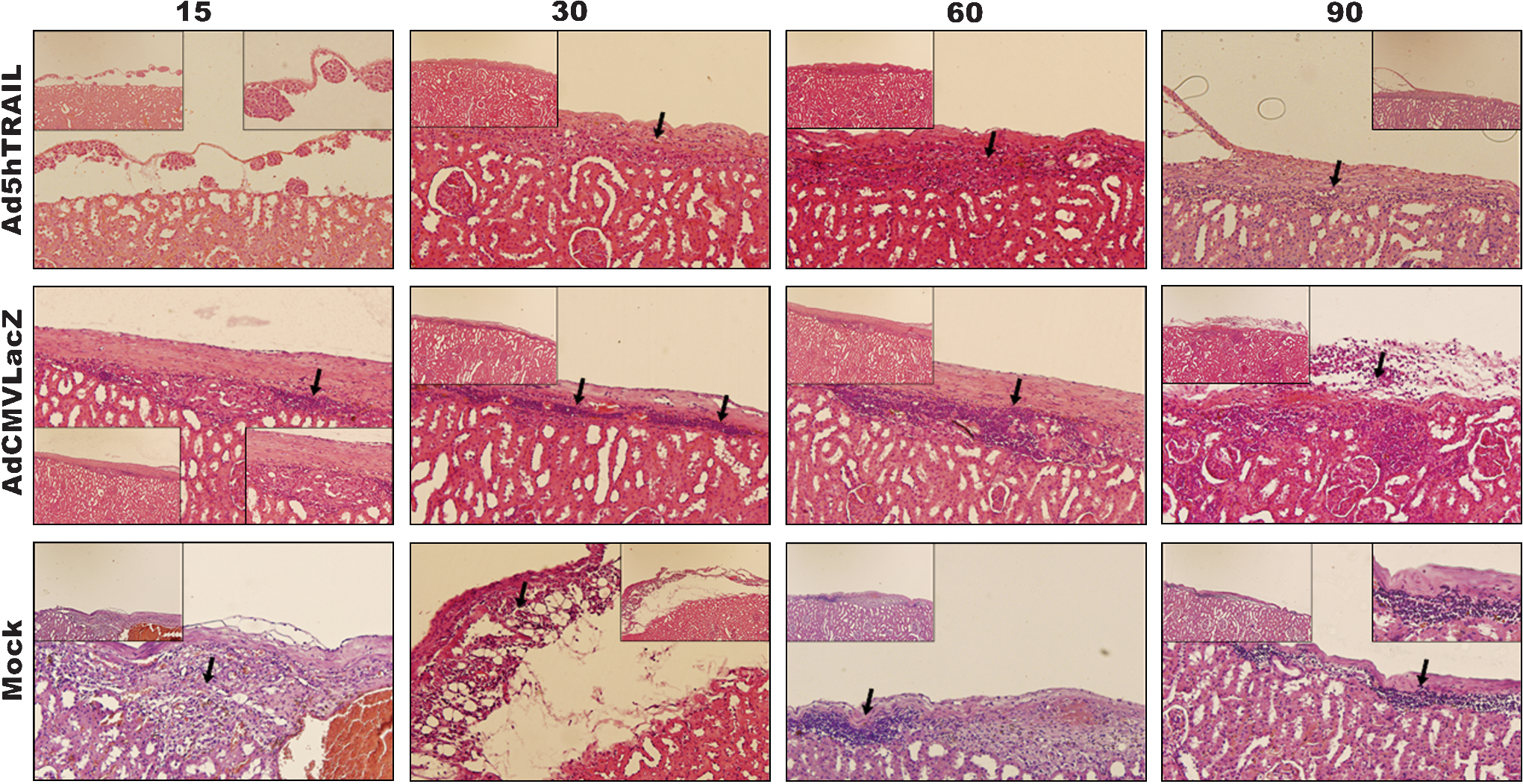
Islet inflammation after transplantation of genetically engineered pancreatic islets. Ad5hTRAIL-, AdCMVLacZ-, or mock-infected rat pancreatic islets were transplanted into STZ-induced diabetic rats as described in Materials and Methods. Grafts were harvested 15, 30, 60, and 90 days after transplantation, and analyzed for mononuclear cell infiltration by H&E staining. Arrows show mononuclear cell deposition sites. The large images were taken at an original magnification of × 200, and × 100 magnifications are also provided for clarity inside each image. Color images available online at
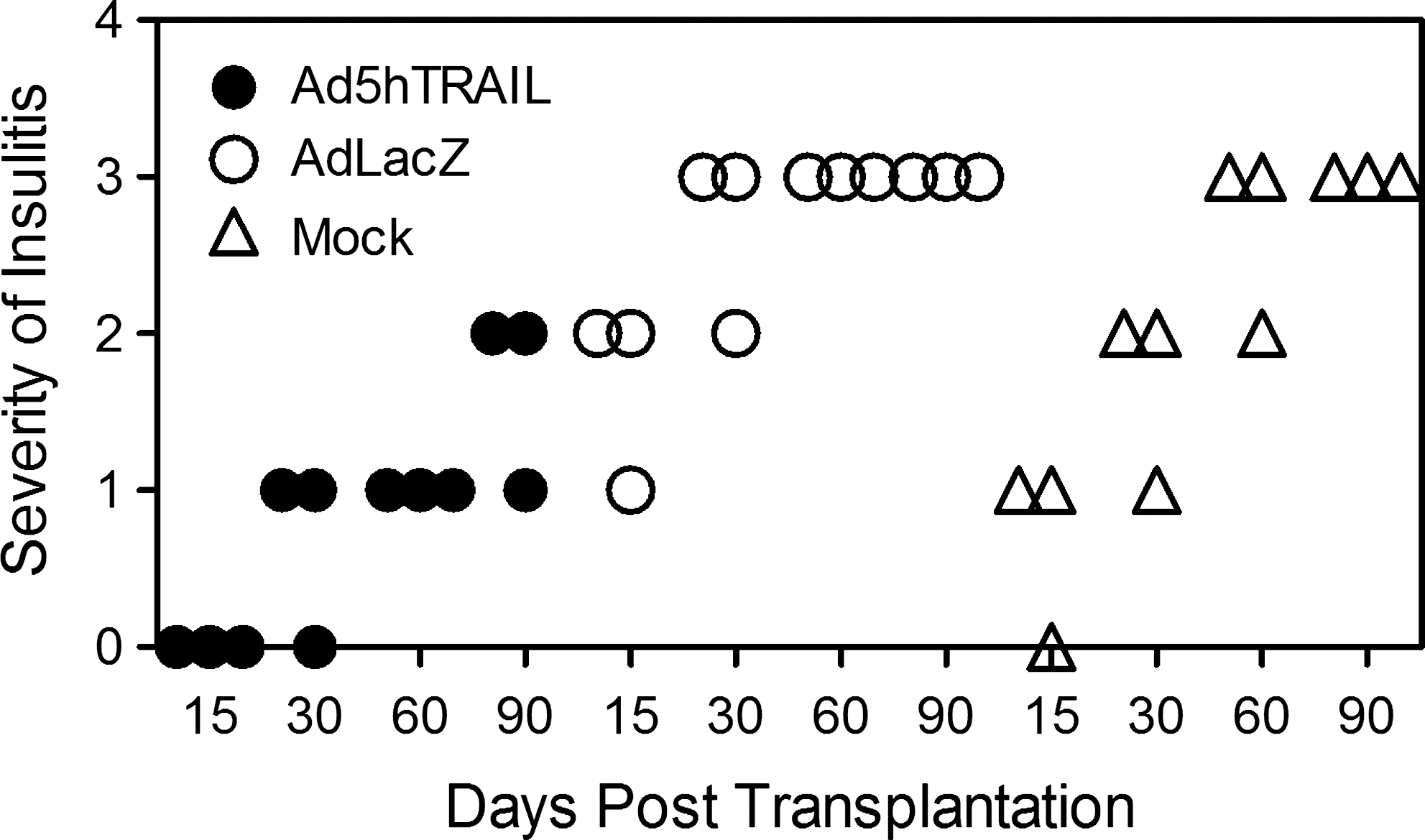
Dot-plot analysis of insulitis scoring. Scores were obtained by histochemical analysis, as described in Materials and Methods, from animals transplanted with Ad5hTRAIL-infected (solid circles), AdCMVLacZ-infected (open circles), or mock-infected (open triangles) islets. Three animals representing each condition were killed on day 15 (TRAIL vs. mock, p > 0.05; TRAIL vs. LacZ, p < 0.001), day 30 (TRAIL vs. mock, p < 0.05; TRAIL vs. LacZ, p < 0.001), day 60 (TRAIL vs. mock, p < 0.001; TRAIL vs. LacZ, p < 0.001), and day 90 (TRAIL vs. mock, p < 0.01; TRAIL vs. LacZ, p < 0.01).
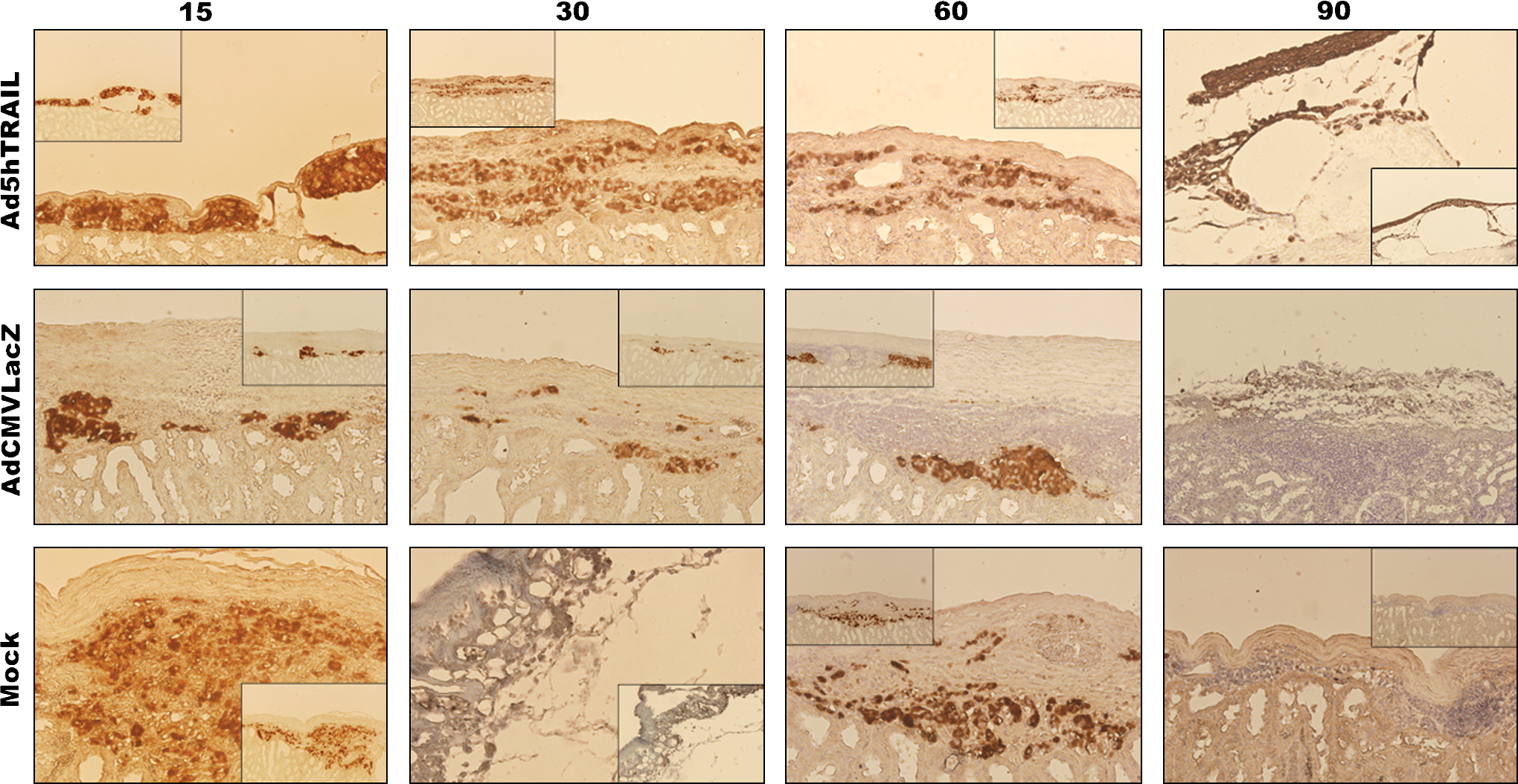
Immunohistochemical analysis of islet allografts. Photomicrographs show insulin in rat islet allografts 15, 30, 60, and 90 days posttransplantation as indicated above each panel. Infection conditions (mock, AdCMVLacZ, or Ad5hTRAIL) are given to the left of each panel. The large images were taken at an original magnification of × 200, and × 100 magnifications are also provided for clarity inside each image. Color images available online at
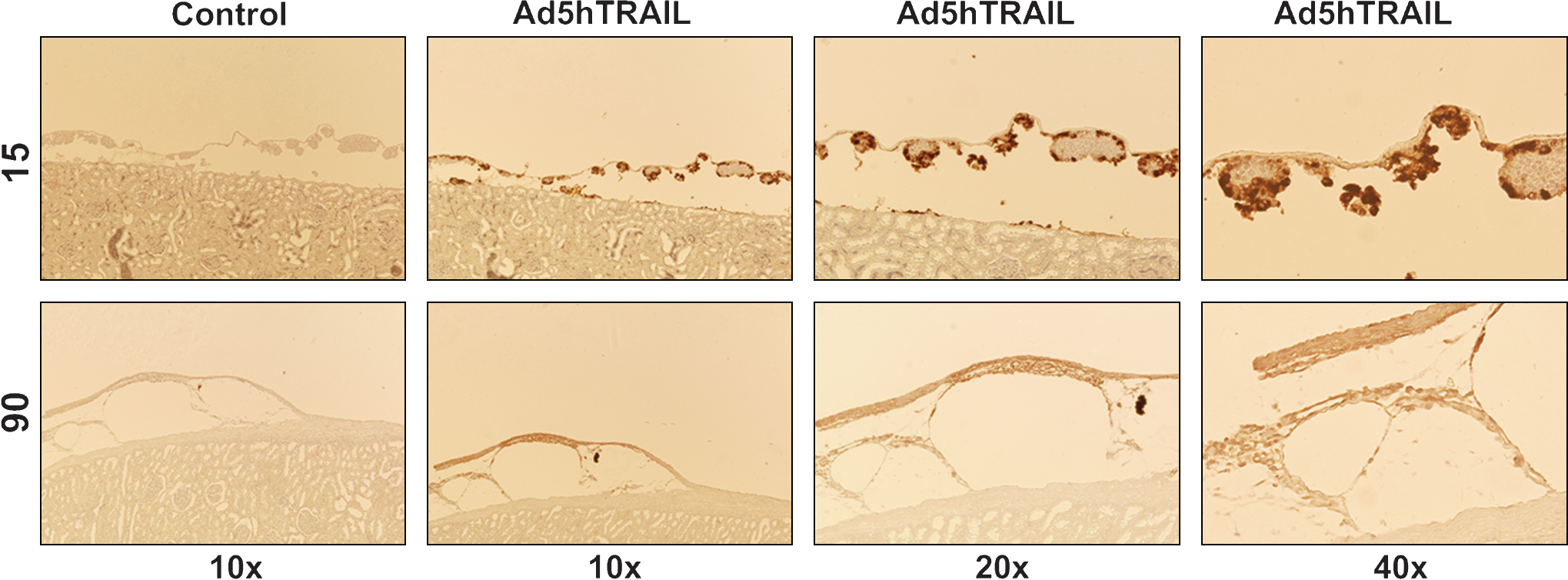
TRAIL expression in pancreatic islet allografts after transplantation. Photomicrographs were taken 15 and 90 days posttransplantation. Control indicates staining performed in the absence of primary antibody. Color images available online at
Together, these data indicate that TRAIL expression in the transplanted islets resulted in decreased inflammation, which ultimately led to prolonged normoglycemia and decreased morbidity in STZ-induced diabetic mice.
Discussion
The ultimate goal of islet transplantation is to completely correct the diabetic syndrome without the need for chronic immunosuppressive drug therapy. By this token, both autoimmune and alloimmune barriers must be overcome to maintain long-term graft function. Thus, tolerance induction clearly represents one of the main objectives in islet transplantation (Chuang et al., 2008). Although many approaches have been devised to protect beta cells from immune-mediated destruction (encapsulation, etc.) (Omer et al., 2003), these methodologies could only delay the eventual failure of the transplanted beta cells. Thus, modification of beta cells by gene therapy arose as a complementary treatment modality to increase the success rate of islet transplantation for the treatment of patients with T1D (D'anneo et al., 2006). The envisioned scenario relies on the premise that pancreatic islet grafts protected by therapeutic gene transfer from immune-mediated cytotoxic T cell attack are expected to function longer after transplantation (Sanlioglu et al., 2008b). For this purpose, engineered expression of members of the TNF family, such as Fas ligand (FasL) (Lau et al., 1996) and TNF (Sanlioglu et al., 2001; Dajani et al., 2007), in pancreatic islets have been tested. As reported previously, adenovirus-mediated expression of FasL induced apoptosis of pancreatic islet allografts and isografts infected ex vivo (Kang et al., 1997; Muruve et al., 1997).
TRAIL is a potent inducer of tumor cell apoptosis, but is nontoxic to normal cells and tissues (Wiley et al., 1995; Terzioglu et al., 2007). This distinctive effect of TRAIL on cell survival/death clearly distinguishes it from other members of the TNF family. However, TRAIL-mediated killing is also important in other immunological settings besides tumor immune surveillance. As previously shown, the absence of TRAIL expression or function increased the degree of islet inflammation and the incidence of autoimmune diabetes (Lamhamedi-Cherradi et al., 2003). And because TRAIL-transduced dendritic cells suppressed antigen-specific T cell activity and protected mice from acute graft-versus-host disease and leukemia relapse (Sato et al., 2005), we hypothesized that increased TRAIL expression would provide immune protection to Langerhans islets by virtue of its potential to protect these cells from CTL attack.
To test this hypothesis, TRAIL was overexpressed in rat pancreatic islets via Ad5hTRAIL infection, and the cells were then transplanted into STZ-induced diabetic outbred rats. Whereas animals transplanted with AdCMVLacZ-infected islets did not regain normoglycemia, those that received Ad5hTRAIL-infected islets experienced prolonged normoglycemia. Intriguingly, AdCMVLacZ-infected islets could also lower blood glucose, but the contribution was not sufficient to establish normoglycemia as observed with Ad5hTRAIL- or mock-infected islets. Because AdCMVLacZ infection did not alter glucose-stimulated insulin secretion of pancreatic islets in vitro, failure to establish normoglycemia can be attributed to the immunity to adenoviral vectors or transgene expression ultimately constraining cell survival/function of insulin-producing pancreatic islet grafts in vivo. The fact that Ad5hTRAIL-infected islet grafts restored normoglycemia more rapidly than mock-infected islets suggests that exogenous TRAIL expression facilitated pancreatic islet transplantation in STZ-induced diabetic rats. Whether or not revascularization of islet grafts is influenced in this process remains to be tested.
Examination of the transplanted islets from the animals transplanted with TRAIL-expressing islets revealed reduced mononuclear cell infiltration into the graft compared with animals transplanted with either AdCMVLacZ-infected or mock-infected islets. It is interesting to note that animals transplanted with mock-infected islets were normoglycemic on day 14 through 45 posttransplantation, despite the presence of mononuclear cell infiltration. Because TRAIL has previously been claimed to be an immune modulator, it is reasonable to propose that TRAIL is suppressing or delaying the mononuclear cell infiltration in this model. Intriguingly, transgenic expression of TNF in islets led to severe insulitis but not diabetes in NOD mice (Higuchi et al., 1992; Picarella et al., 1993). These findings suggest that insulitis might be necessary, but not sufficient, to induce diabetes. In this scenario, the inhibition of autoreactive islet-specific T cells is held responsible for the observed phenotype (Grewal et al., 1996). Conversely, islet-specific neonatal expression of TNF accelerated the onset and penetrance of diabetes by enhancing presentation of islet antigens in transgenic NOD mice (Green et al., 1998). On the basis of these conflicting results, it was reasoned that the late islet-specific expression of TNF, but not early neonatal expression, could abrogate ongoing autoimmune process in T1D (Christen et al., 2001). Therefore, the consequence of islet-specific TNF expression varies depending on the timing of expression. TRAIL, in contrast, can prevent the activation and expansion of lymphocytes in vivo, but does not delete them from the system (Song et al., 2000). Thus, contrary to TNF or FasL, TRAIL may be vital in downregulating autoimmune responses. Our findings also substantiate a previous report suggesting that adenovirus overexpression of interleukin receptor antagonist protein-1 (IRAP-1) in islet grafts reduced lymphocyte infiltration (Tellez et al., 2005). It is worth mentioning that, contrary to AdCMVLacZ-infected or mock-infected islets, only Ad5hTRAIL-infected islets maintained insulin expression 90 days after transplantation.
Although eventual rejection of the allograft remains a problem to be solved, we have clearly demonstrated in this study that prolongation of islet allograft survival is achievable by ex vivo adenoviral-mediated TRAIL gene transfer. By this token, the use of gutless adenoviral or adeno-associated viral (AAV) vectors is expected to yield a longer therapeutic benefit in extending allograft survival and function (Flotte et al., 2001; Kapturczak et al., 2001).
Footnotes
Acknowledgments
This work is supported by grants from the Scientific and Technological Research Council of Turkey (TUBITAK-104S224) and Akdeniz University Scientific Research Project Administration Division (2007.03.0122.003). The authors thank Fatma Zehra Hapil for technical assistance.
Author Disclosure Statement
No competing financial interests exist.