
Abstract
Conditional elimination of infused gene-modified alloreactive T cells, using suicide gene activation, has been shown to be an efficient strategy to abrogate severe graft-versus-host disease (GvHD) in the context of adoptive immunotherapy. To overcome shortcomings of the most widely used suicide gene, wild-type (splice-corrected) herpes simplex virus thymidine kinase (scHSVtk), we generated two new variants: the codon-optimized coHSVtk and, by introducing an additional mutation (A168H), the novel TK.007. We transduced human hematopoietic cell lines and primary T cells with retroviral “sort-suicide vectors” encoding combinations of selection markers (tCD34 and OuaSelect) with one of three HSVtk variants. In vitro we observed higher expression levels and sustained long-term expression of TK.007, indicating lower nonspecific toxicity. Also, we noted significantly improved kinetics of ganciclovir (GCV)-mediated killing for TK.007-transduced cells. In an experimental (murine) allogeneic transplantation model, TK.007-transduced T cells mediated severe GvHD, which was readily abrogated by application of GCV (10 mg/kg). Last, we established a modified allotransplantation model that allowed quantitative comparison of the in vivo activities of TK.007 versus scHSVtk. We found that TK.007 mediates both significantly faster and higher absolute killing at low GCV concentrations (10 and 25 mg/kg). In summary, we demonstrate that the novel TK.007 suicide gene combines better killing performance with reduced nonspecific toxicity (as compared with the frequently used splice-corrected wild-type scHSVtk gene), thus representing a promising alternative for suicide gene therapy.
Introduction
Alongside its promising performance, there are several shortcomings of the HSVtk/GCV system that have become apparent during its clinical use. First, GCV is relatively toxic at clinically used doses, in particular toward myelopoiesis (Biron, 2006). Consequently, prolonged application may result in profound immunosuppression. Second, the maximal killing efficiency of the HSVtk/GCV system is reached only after about 4–6 days (Fehse et al., 2002; Lemoine et al., 2004) in vitro and >6 days in vivo (Tiberghien et al., 2001). A faster cell-killing rate would often be advantageous, in particular for the elimination of modified allogeneic lymphocytes in order to abrogate severe acute graft-versus-host disease (GvHD), which is associated with high morbidity. Third, the viral HSVtk gene contains an active cryptic splice site, resulting in a fraction of up to 5% of transduced target cells containing an inactive spliced HSVtk variant, rendering these cells insensitive to GCV (Garin et al., 2001). Moreover, translation initiation sites downstream of the start codon have been observed (al-Shawi et al., 1991; Salomon et al., 1995) to generate tissue-dependent truncated HSVtk variants with low GCV sensitivity at low expression levels (Salomon et al., 1995). Fourth, elevated HSVtk expression levels are associated with a certain toxicity in transduced cells. This is, to a great extent, due to the relatively high affinity of HSVtk for endogenous thymidine (K m, 0.5 μM) (Munir et al., 1994; Balzarini et al., 2006). Additional limitations to the HSVtk/GCV system include the preferred use of GCV for treatment of cytomegalovirus reactivation after allogeneic stem cell transplantation (SCT), leading to the unwanted elimination of HSVtk-modified donor lymphocytes in this particular situation and the strong immunogenicity of HSVtk (reviewed in Muul and Candotti, 2007), which might, on the other hand, be of advantage in tumor therapy.
To overcome the described limitations of the HSVtk/GCV system, alternative cell suicide systems have been developed. These include the “classical” suicide gene/prodrug combinations cytosine deaminase/5-fluorocytosine and cytochrome P450/cytochrome P450 reductase, already in clinical trials (Portsmouth et al., 2007). More recent combinations include the human thymidylate monophosphate kinase (TMPK) with the prodrug zidovudine (AZT) (Ostermann et al., 2000), killing from dimerization by the FK506 molecule using either a modified Fas receptor (Thomis et al., 2001) or an inducible caspase-9 (Straathof et al., 2005), and the CD20/rituximab system (Introna et al., 2000). Altogether, the alternative suicide systems are either still under preclinical/early clinical evaluation or do not appear superior to the HSVtk/GCV system regarding the risk/downside-versus-benefit ratio. In particular, the bystander effect of HSVtk/GCV (Moolten, 1986), critical for its efficacy in tumor treatment, is unlikely to be achievable by nonenzymatic approaches.
In view of the previously described limitations, efforts have been made to improve the established HSVtk/GCV system. Black and colleagues (2001) have identified various HSVtk variants with increased GCV sensitivity by semirandom sequence mutagenesis. Among them, the most promising was the SR39 mutant, demonstrating a 14-fold decrease in K m for GCV compared with wild-type (wt) HSVtk. However, the turnover number (k cat) for GCV was also markedly decreased by the SR39 mutations, demonstrated to be 27-fold lower for the SR39 HSVtk mutant than for the wtHSVtk enzyme (Kokoris and Black, 2002). This is likely the reason why conflicting results have been generated in the evaluation of SR39 HSVtk. Some studies reported no enhanced performance of SR39 HSVtk, whereas others confirmed its improved GCV sensitivity (Black et al., 2001; Pantuck et al., 2002; Qasim et al., 2002; Rettig et al., 2003). Other mutants discovered include Q7530, which improved the sensitivity of the enzyme for GCV (Mercer et al., 2002), and TK30, which showed a better bystander effect (Qiao et al., 2000).
Decreasing the affinity of HSVtk for endogenous thymidine represents a novel approach to improve the system. In a targeted mutagenesis study, a novel HSVtk mutant (A168H) with a fully preserved GCV catalytic substrate activity in conjunction with a heavily compromised deoxythymidine (dThd) kinase activity was identified (Balzarini et al., 2006). Interestingly, the enzyme efficiency (V max/K m) of the A168H mutant for GCV was 26 as compared with 6.1 for wtHSVtk, suggesting a superior GCV/dThd catalytic ratio of that mutant compared with wtHSVtk and previously identified mutants (Balzarini et al., 2006). These results were obtained in biochemical assays of purified enzymes. But, to our knowledge, the functional activity of the A168H mutant as a transgene has not yet been reported.
In this study, we assessed the function of the HSVtk A168H mutant TK.007 in order to evaluate its potential as a suicide gene. Furthermore, to overcome problems associated with the primary sequence of wild-type HSVtk and thus improve its applicability and safety in clinical settings, we constructed a fully codon-optimized HSVtk gene. Codon optimization for gene expression in human cells has previously been shown to improve RNA half-life and translation efficiency, and in addition to disrupt cryptic splice sites and regulatory elements (Wagner et al., 2000; Moreno-Carranza et al., 2009). Here we evaluated a fully codon-optimized HSVtk (coHSVtk) and a fully codon-optimized A168H mutant of HSVtk (TK.007) as compared with the clinically used splice-corrected wild-type HSVtk gene sequence (scHSVtk) (Chalmers et al., 2001). To do so we used gammaretroviral sort-suicide vectors designed for transducing T lymphocytes in various in vitro and in vivo models of adoptive immunotherapy.
Materials and Methods
Vector plasmids
Construction of MP71-tCD34/scHSVtk is described elsewhere (Fehse et al., 2002). Codon-optimized coHSVtk and the analogously codon-optimized HSVtk(A168H) mutant, TK.007, were synthesized by GENEART (Regensburg, Germany). MP71-tCD34/coHSVtk and MP71-tCD34/TK.007 were generated by replacing the initial, splice-corrected scHSVtk gene with the two codon-optimized HSVtk variants. MP71.coHSVtk-OuaS.pre* vector construction was performed by GENEART. OuaS was cloned from SF91-EGFP-OuaS.pre including a codon-optimized OuaSelect gene described previously (Treschow et al., 2007), coHSVtk from MP71-tCD34-coHSVtk and the optimized pre* from pRSF91.EGFP.pre* described previously (Schambach et al., 2006), using PCR and the NotI and NheI sites in MP71-tCD34-coHSVtk. The start codon of OuaSelect was removed to reduce the risk of aberrant translation initiation. MP71.TK.007-OuaS.pre* was generated by replacing the RsrII–SbfI fragment of MP71.coHSVtk-OuaS.pre* with that of MP71-tCD34/TK.007.
Primary cells and cell lines
Peripheral blood mononuclear cells (PBMCs) were isolated from healthy donor blood by centrifugation on a Ficoll gradient (1200 rpm for 20 min). T cells used in the CD34/HSVtk experiments were stimulated with CD3/CD28 DynaBeads (Invitrogen, Carlsbad, Germany) and were cultivated at 2 × 106/ml in the presence of interleukin (IL)-2 (100 U/ml; Cancernova, Reute, Germany) in X-VIVO 10 medium (BioWhittaker, Verviers, Belgium) containing 8% autologous serum (heat inactivated at 56°C for 30 min). In the HSVtk-OuaS experiments, PBMCs were cultivated in X-VIVO 15 (BioWhittaker), supplemented with autologous serum and IL-2 (PeproTech, London, UK) as described previously. All cells were kept at 37°C in a humidified atmosphere in CO2 incubators.
Mice
C57BL/6.Ly5.1 (CD45.1+) mice were obtained from Charles River Laboratories (Sulzfeld, Germany) and BALB/c Rag2−/−/γc −/− mice (Traggiai et al., 2004) from the Laboratory of M. Manz (Bellinzona, Switzerland). Animals were bred and maintained under pathogen-free conditions in the animal facilities of the Georg Speyer Haus (Frankfurt am Main, Germany). Cages were individually ventilated. Animals were killed after anesthesia by cervical dislocation. The experiments were performed in compliance with the local animal experimentation guidelines. Animal experiments were approved by regional council (Regierungspräsidium, Darmstadt, Germany).
Retroviral vector production
Phoenix-GP cells (kindly provided by G. Nolan, Stanford University, Stanford, CA) were employed for transient gammaretroviral vector production as previously described (Li et al., 2003). Producer cells were transfected with a calcium phosphate transfection kit (Sigma-Aldrich, St. Louis, MO) in accordance with the manufacturer's instructions. Medium was changed 6–10 hr posttransfection. Virus-containing supernatant was harvested every 12 hr. MP71 vectors were pseudotyped with Env proteins from gibbon ape leukemia virus (GALV-env) (Fehse et al., 2002) or ecotropic murine leukemia virus (Eco-env) (Newrzela et al., 2008).
Retroviral transduction and selection of transduced cells
K562 cells, PM1 cells, CEM cells, and Jurkat cells were transduced with retroviral vectors by centrifugation at 1000 × g for 1 hr at room temperature in the presence of protamine sulfate (4 μg/ml). Primary human T cells were transduced with tCD34/HSVtk vectors at a density of 1 × 106/ml in 3 ml of medium in 6-well tissue culture plates. Fibronectin-coated plates (BD Biosciences, San Jose, CA) were used for HSVtk-OuaS vector transduction. Multiplicities of infection (MOIs) were adjusted to obtain transduction efficiencies below 30%, corresponding to one vector copy per cell (Kustikova et al., 2003). Cells were kept in culture for at least 2 days before the analysis of gene transfer efficiency by fluorescence-activated cell sorting (FACS) and 3 days before copy number assessment by real-time PCR. Expression of tCD34/HSVtk fusion proteins was monitored with CD34 antibodies (QBEnd10; Beckman Coulter, Krefeld, Germany). K562, PM1, and primary human T cells transduced with either MP71-tCD34-HSVtk vector were selected by magnetic cell sorting (MACS), using a CD34 MicroBead kit, and MiniMACS columns (both from Miltenyi Biotec, Bergisch Gladbach, Germany) in accordance with the manufacturer's instructions. MP71.coHSVtk-OuaS.pre*- or MP71.TK.007-OuaS.pre*-transduced cells (CEM, Jurkat, and primary T cells) were selected in 1 μM ouabain for 48 hr as described previously (Treschow et al., 2007).
In vitro ganciclovir sensitivity assay
To evaluate killing kinetics in hematopoietic cells, nontransduced control cells and cells transduced with CD34 vectors were seeded at a density of 0.5 × 106/ml (cell lines) or 1 × 106/ml (primary human T cells) in duplicate and incubated without or with 1- and 4-μg/ml concentrations of ganciclovir (GCV, Cymeven; Roche, Basel, Switzerland). Cells were replated at the same density every counting day. MP71.coHSVtk-OuaS.pre*- or MP71.TK.007-OuaS.pre*-transduced and selected CEM and Jurkat cells and untransduced controls were seeded at 0.2 × 106 cells/ml and were exposed to GCV (1 or 3 μg/ml) in triplicate. OuaS vector-transduced and selected primary T cells were seeded at a density of 1 × 106/ml with or without GCV (1 μg/ml). Absolute numbers of viable cells were determined every second/third day on the basis of trypan blue exclusion staining over a total time of 6 days to 1 week (Fehse et al., 2002).
Transduction and transplantation of murine cells
Murine T cells were transduced and transplanted in accordance with our published protocol (Newrzela et al., 2008). In brief, murine mononuclear cells were isolated from spleen and lymph nodes of C57BL/6.Ly5.1 mice and stimulated with anti-CD3 (clone 145-2C11), anti-CD28 monoclonal antibody (mAb, clone 37.51; BD Biosciences) paramagnetic beads (Invitrogen) for 3 days to obtain stimulated T cells. On day 4 of stimulation cells were transduced with retroviral vectors by transfer onto vector-preloaded, RetroNectin-coated 6-well non-tissue culture plates (BD Biosciences). Plates were preloaded with viral particles by centrifugation (1000 × g) at 4°C for 90 min. Throughout the entire cultivation time, cells were kept in RPMI 1640 supplemented with 10% fetal calf serum, 2%
Flow cytometry
Flow cytometry was carried out on cell lines, primary human T cells, and blood of mice (blood was drawn from the tail vein) as previously described (Fehse et al., 2002; Newrzela et al., 2008). The following antibodies were used for staining: phycoerythrin (PE)-conjugated anti-human CD34 (Beckman Coulter), cyanine 5 (Cy5)-conjugated anti-mouse CD3 (Invitrogen), and fluorescein isothiocyanate (FITC)-conjugated anti-mouse CD45.1 (A20; BD Biosciences).
Administration of GCV in vivo
In the acute GvHD (aGvHD) model, mice were examined every day for symptoms of severe aGvHD (see Fig. 4A). Criteria included >10% weight loss, scrubby fur, diarrhea, and loss of activity. If an animal met three of the four criteria, treatment was initiated: GCV was administered at 10 mg/kg for three consecutive days by intraperitoneal injection once per day. In the “quantitative mouse model” (see Fig. 5A), GCV was administered by intraperitoneal injections at various doses (50, 25, and 10 mg/kg) once daily over five consecutive days.
Real-time PCR
Transduction rates of OuaS vectors were assessed by real-time PCR and the ΔC t method (Kröger et al., 2007). DNA from samples of 3 × 106 unselected cells was prepared with a QIAamp DNA mini kit (Qiagen, Hilden, Germany) according to the manufacturer's description. For the quantitative analysis of proviral DNA copies, the 7500 fast real-time system (Applied Biosystems, Foster City, CA) was used. Primers and probes for coHSVtk were designed with Primer Express software (Applied Biosystems) and synthesized by Applied Biosystems: coHSVtk forward primer, 5′-GACATCGTGTACGTGCCCG-3′; coHSVtk reverse primer, 5′-TGTGTGGTGTAGATGTTGGCG-3′; coHSVtk probe, 5′-FAM-CCTACTGGCAGGTGCTGGGCGC-TAMRA-3′. Probes and primers used for detection of the glyceraldehyde-3-phosphate dehydrogenase (GAPDH) gene (DNA content control) have been described previously (Alizadeh et al., 2002). C t values obtained for GAPDH were subtracted from the C t values for coHSVtk. ΔC t values were plotted against a standard curve as previously described (Kröger et al., 2007). Samples of primary T cells of known ratios of coHSVtk-OuaS-transduced and selected cells were prepared so as to create standard curves for quantification of unknown samples. Each reaction was made with 10 μl of ready-to-use Taq mix (Applied Biosystems), 100 ng of DNA, 100 nM (GAPDH) or 50 nM (coHSVtk) primers, and 100 nM probe, which were mixed and adjusted to 20-μl volume with deionized H2O. Each sample was tested in triplicate. Standard conditions were used for the PCR: 3 min at 95°C, and then 45 cycles of 15 sec at 95°C and 1 min at 60°C.
Results
Construction of gammaretroviral suicide vectors
Altogether, we used five different vectors in our preclinical adoptive immunotherapy studies in vitro and in vivo (Fig. 1A and B). All vectors are based on the gammaretroviral MP71 backbone (Baum et al., 1995) and express combinations of two different marker genes, tCD34 (Fehse et al., 2000) and OuaSelect (Treschow et al., 2007) (OuaS), and three HSVtk variants. In experiments with tCD34 vectors, MP71-tCD34/scHSVtk (Fehse et al., 2002), containing a splice-corrected, wild-type HSVtk (Chalmers et al., 2001), was used as a control. The new vectors MP71-tCD34/coHSVtk and MP71.coHSVtk-OuaS.pre* contain a novel, fully codon-optimized (co) HSVtk. The A168H mutation was introduced into coHSVtk in MP71-tCD34/TK.007 and MP71.TK.007-OuaS.pre* vectors.

Retroviral herpes simplex virus thymidine kinase (HSVtk) vectors used in this study. (
Improved expression of tCD34/TK.007, as compared with both tCD34/scHSVtk and tCD34/coHSVtk, indicates lower toxicity of TK.007
To assess expression levels of the novel suicide genes, various hematopoietic cell lines, as well as primary human T cells, were transduced with MP71 vectors encoding the different HSVtk variants (fused to tCD34) and analyzed. To allow direct gene comparison, low MOIs were applied to ensure a mean copy number of one per transduced cell (Fehse et al., 2004b). Notably, in several independent experiments we reproducibly found higher tCD34/HSVtk expression levels per cell (as indicated by increased mean fluorescence intensities, MFIs) in K562 cells (Fig. 2A) as well as in PM1 cells (data not shown) transduced with MP71-tCD34/TK.007 compared with cells transduced with either MP71-tCD34/coHSVtk or MP71-tCD34/scHSVtk. As shown in Fig. 2B, summarizing data from 12 independent experiments with K562 cells, the differences were significant in relation to both tCD34/coHSVtk and tCD34/scHSVtk. It is notable that there was essentially no difference in expression between tCD34/coHSVtk and tCD34/scHSVtk, indicating that codon optimization was not sufficient to mediate higher expression levels.
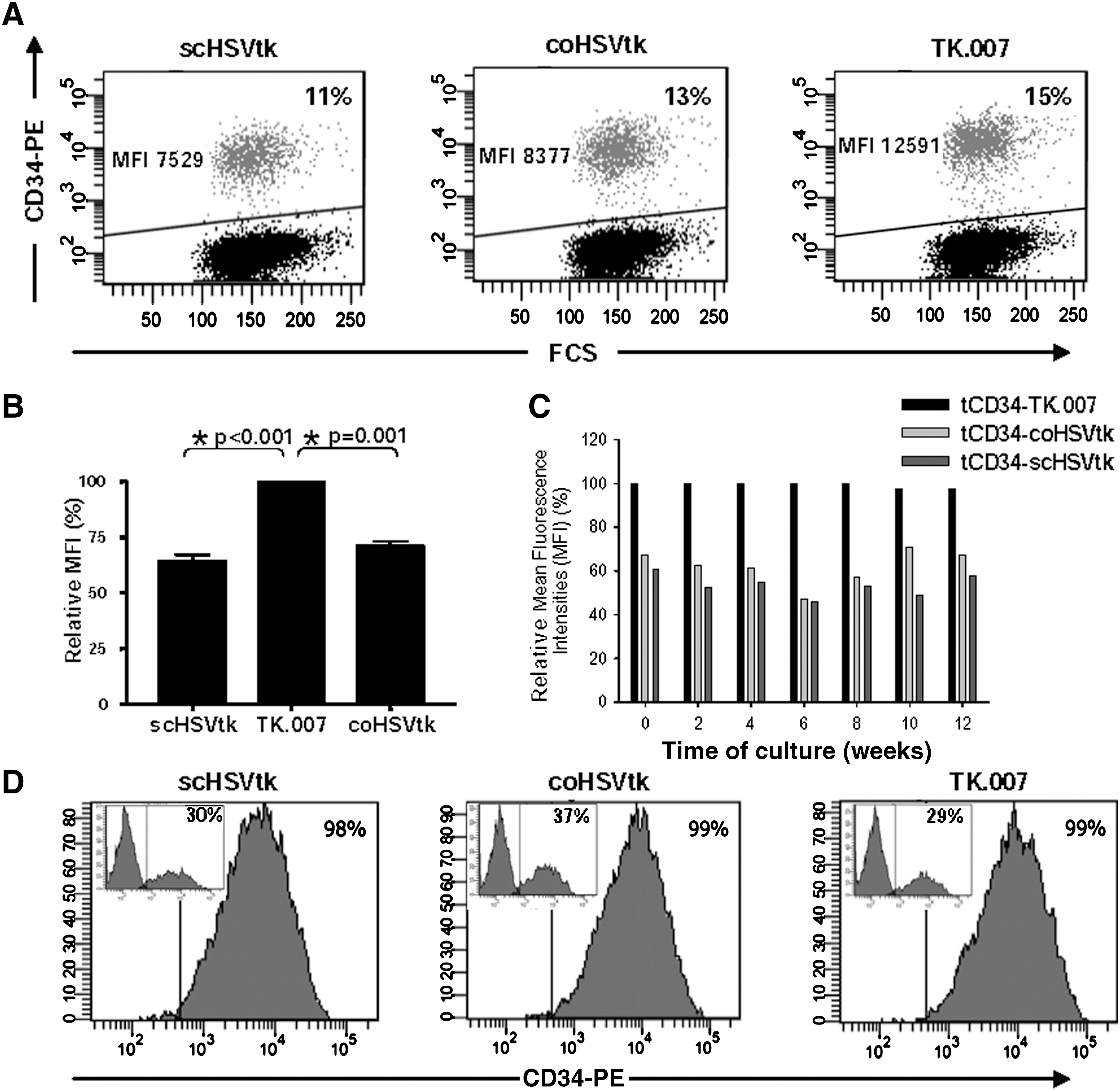
Improved and stable long-term expression of novel HSVtk variants. (
To assess stability of expression over time for the different HSVtk variants, K562 and PM1 cells transduced with one copy per cell were kept in culture for an observation period of 12 weeks. Expression levels were measured every other week by CD34 staining and flow cytometric analysis. Over the 12-week period we observed essentially stable protein expression (measured on the basis of MFIs), independent of the HSVtk gene used. Consequently, the initially established advantage in protein expression for the A168H mutant (see above, Fig. 2A and B) was conserved over time for both K562 cells (Fig. 2C; p < 0.0002) and PM1 cells (data not shown). These data were confirmed in two independent long-term experiments for both cell lines. It again emphasizes that the A168H mutation into HSVtk was the main factor for the higher expression of the fusion gene tCD34/TK.007 as compared with tCD34/coHSVtk and tCD34/scHSVtk.
Last, we evaluated whether the expression levels of the three fusion genes were high enough to facilitate CD34-directed magnetic cell sorting (MACS). We reached efficient enrichment (>95%) of both PM1 and K562 cells transduced with any of the HSVtk variants (data not shown). In accordance with this, primary T lymphocytes transduced with any of the three tCD34/HSVtk vectors were efficiently enriched by MACS to high purities (>98%; Fig. 2D).
TK.007 facilitates faster killing of cultured hematopoietic cells independently of the coexpressed selection marker
Next, we tested the GCV sensitivity of highly enriched hematopoietic cells transduced with any of the three gammaretroviral tCD34/HSVtk constructs described previously (Fig. 1A). Transduced and magnetically sorted cells (K562 and PM1 cells, described previously) were cultured in various concentrations of GCV (1 and 4 μg/ml) for 1 week. Absolute cell numbers were determined on days 2, 4, and 7. As shown in Fig. 3A, at a low GCV concentration (1 μg/ml) cell killing occurred significantly faster for cells transduced with MP71-tCD34/TK.007 than for cells transduced with the standard vector MP71-tCD34/scHSVtk. In fact, most cells (60–80%, depending on the cell type) transduced with the vector encoding TK.007 had already been eliminated after 2 days of GCV application, independent of the investigated cell type (Fig. 3A).
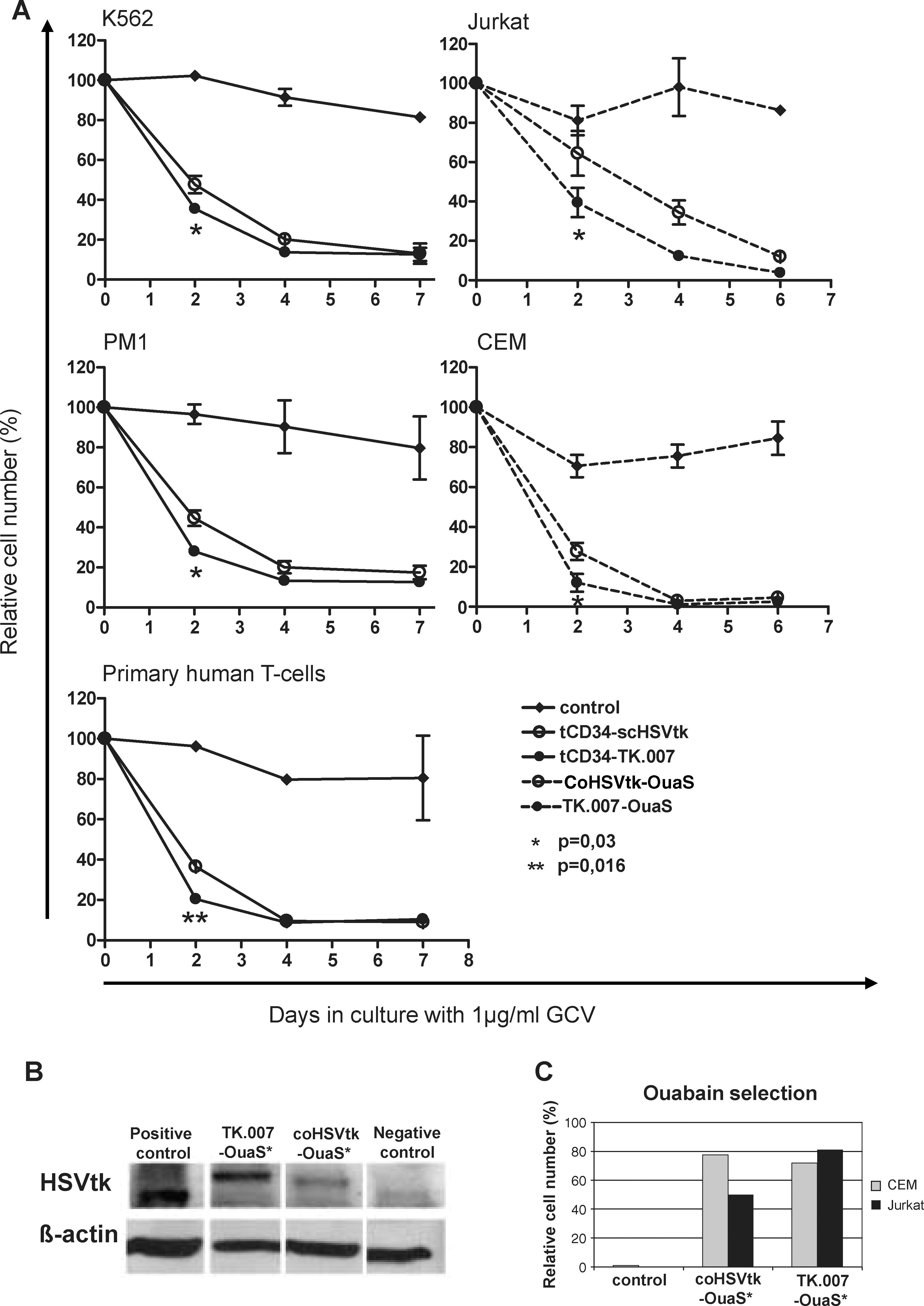
Superior functional performance of TK.007 in hematopoietic cells. (
In a separate set of experiments, the novel codon-optimized HSVtk variants were tested as fusion genes with the OuaSelect marker. The same MP71 gammaretroviral backbone was used in these studies (Fig. 1B), with the inclusion of the optimized woodchuck hepatitis posttranscriptional regulatory element (wPRE*) (Schambach et al., 2006). Lymphoid cells (CEM and Jurkat) were transduced at low MOIs. Transduction rates were estimated by real-time PCR as 22–29% for CEM cells and 11–14% for Jurkat cells. Transduced cells were selected for 48 hr in the presence of 1 μM ouabain. The purity after cell selection reached >99% because less than 1% of untransduced cells survived selection whereas good recovery rates were found for gene-modified cells (Fig. 3C). Notably, transduction efficiencies estimated on the basis of quantitative PCR (qPCR) for integrated proviral copy number are likely to overestimate functional transduction rates (e.g., protein expression by flow cytometry), which might explain the observed noncomplete recovery rates after selection. Differences in transduction rates as estimated on the basis of copy number versus protein expression have been shown to vary 10- to 60-fold, depending on protein expression level (Sastry et al., 2002; Lizee et al., 2003). Western blots demonstrated that the TK.007-OuaS protein was expressed to higher levels than coHSVtk-OuaS in transduced and selected CEM cells (Fig. 3B). Jurkat cells expressed lower levels of all transgenes, resulting in weaker bands, with the TK.007 signal still being most prominent (data not shown).
Functional activity of HSVtk in the various vectors was determined by GCV exposure. The rate of cell death induced by exposure to GCV was more rapid for TK.007- than for coHSVtk-expressing cells (Fig. 3A), which was in accordance with results obtained for the CD34 vectors (see previously). On the basis of summarized data obtained with the various cell lines the faster (i.e., more efficient) killing of cells transduced with vectors expressing TK.007 as compared with scHSVtk was significant (day 2: p = 0.03; Wilcoxon matched pairs test).
To assess transgene functionality after long-term cultivation, magnetically sorted K562 and PM1 cells as well as ouabain-selected CEM and Jurkat cells were cultured for approximately 12 weeks, before they were exposed to GCV. Again, in two independent experiments lymphoid PM1 cells transduced with the codon-optimized A168H mutant TK.007 showed superior killing kinetics compared with cells transduced with scHSVtk and coHSVtk (data not shown). In line with this, faster killing was also seen for long-term cultivated CEM and Jurkat cells transduced with TK.007-OuaS as compared with coHSVtk-OuaS (data not shown). No significant differences were seen between the three HSVtk variants in K562 cells cultivated for 12 weeks (data not shown).
Rapid killing of primary T cells transduced with retroviral TK.007 vectors
To assess the functionality of TK.007 in primary T cells we once more made use of the two different sort/selection vectors available in the participating laboratories. As shown in Fig. 2D, primary T cells transduced with any of the three tCD34/HSVtk vectors at approximately one vector copy per cell were readily sorted by MACS to high purity. Again, treatment of those enriched cells with GCV at 1 μg/ml resulted in faster killing of TK.007-transduced cells as compared with both scHSVtk- (Fig. 3A) and coHSVtk-transduced primary T lymphocytes.
Transduction of primary T cells with TK.007-OuaS or coHSVtk-OuaS vectors resulted in 30–56% gene transfer as estimated by real-time PCR (four independent experiments with cells from different donors). As noted previously, these data were possibly overestimating the capacity of transduced cells to express the transgene (see previously; and see Sastry et al., 2002; Lizee et al., 2003). Also, at early time points nonintegrated vector copies might influence PCR-based assessment of transduction efficiencies. In line with this, recovery rates of transduced primary T cells after ouabain selection were 7.7% (range, 1.2–17.6%) for coHSVtk-OuaS and 9.2% (range, 1.6–18.2%) for TK.007-OuaS positive cells.
Transduced and selected T cells were exposed to GCV (1 μg/ml) for 7 days and a rapid elimination of transduced cells was observed. As with tCD34 vectors the TK.007 gene was superior to the wild-type protein for OuaS vectors regarding the rapidity of cell killing. In fact, on the basis of seven independent experiments with primary T cells transduced with either tCD34 vectors or OuaS vectors the difference in TK.007- versus coHSVtk-mediated killing rates was highly significant (p < 0.016, Wilcoxon matched pairs test) after 2 days of treatment with GCV at 1 μg/ml. Large interassay variations could be attributed to different T cell growth kinetics seen for individual cell donors.
TK.007-transduced murine T cells mediate severe GvHD in vivo, which can be abrogated by GCV treatment
We next asked whether the novel tCD34/TK.007 gene also allows abrogation of severe GvHD in vivo, in the mouse model. To do so, we made use of Rag2−/−/γc −/− mice (Traggiai et al., 2004), thus ensuring efficient T cell engraftment. We modified our previously established transplantation model (Newrzela et al., 2008) by using allogeneic donor T cells (Fig. 4A), which would be expected to cause acute GvHD. In fact, in small-scale preliminary dose-finding experiments we found severe GvHD to be caused by 1 × 106 nonmodified fresh T cells in the described model (data not shown).

In an allogeneic transplantation model TK.007-transduced murine T cells cause severe, potentially lethal graft-versus-host disease (GvHD) that can be abrogated by ganciclovir treatment. (
We transplanted mice with 1 × 106 allogeneic T cells transduced with either MP71-tCD34/TK.007 (n = 7) or MP71-tCD34/scHSVtk (n = 7; control). Efficient engraftment of tCD34-positive allogeneic T cells was confirmed for both groups by FACS analysis of peripheral blood (Fig. 4B).
A tight regimen was implemented (with daily physicals, including weighing and analysis of activity, fur, feeding responses, and feces) in the follow-up period to promptly diagnose severe aGvHD in accordance with the four criteria listed in Materials and Methods. Importantly, between 20 and 40 days after infusion of gene-modified allogeneic T cells all transplanted mice in both groups developed severe GvHD, proving that the transgenes used had no specific deleterious effect on T cell functionality. In both groups, one mouse was not treated and died within about 1 week. An autopsy was performed, confirming severe gut GvHD (data not shown). In both groups (TK.007 vs. scHSVtk) all other mice (n = 6) were subjected to treatment with GCV (10 mg/kg) for three consecutive days. In the TK.007 group, all mice quickly recovered, which was reflected by fast weight gain (Fig. 4C). Similarly, five of six mice in the scHSVtk control group reached complete remission; one mouse could not be rescued and died of severe GvHD (Fig. 4D). In the follow-up, four of six mice in the TK.007 group and three of six in the scHSVtk group developed relapses of their aGvHD and eventually had to be killed because of the severity of disease. This indicates that in most animals the 3-day low-dose GCV treatment regimen, although effective in abrogating the acute disease, was not sufficient to deplete all alloreactive T cell clones. On the other hand, in both groups two of six mice remained in continuous complete remission during the further observation (up to 3 weeks), emphasizing the high efficacy of the suicide mechanism.
Altogether, data from this transplant experiment confirmed the functionality of tCD34/TK.007-transduced T cells in vivo as well as the effectiveness of the suicide mechanism in abrogating aGvHD. At the same time, although we found some difference in performance between TK.007 and scHSVtk, it was impossible to assess them quantitatively in the given model. Therefore, we pursued this issue with a modified model, which was intended to detect quantitative differences between the two HSVtk variants, if they existed.
Higher sensitivity of TK.007-transduced T cells toward GCV in vivo
To compare the functional performance of TK.007 and scHSVtk in vivo in a quantitative manner, we introduced two modifications in the allogeneic T cell transplantation model. Instead of infusing highly enriched populations of gene-modified cells (after MACS), we transplanted a mixture of TK-transduced and nontransduced cells (Fig. 5A). To exclude copy number effects we used low MOIs for both vectors (MP71-tCD34/TK.007 and MP71-tCD34/scHSVtk), aiming at gene transfer rates of approximately 30%. For each vector, altogether 12 mice were transplanted and showed efficient engraftment of allogeneic T cells. In the treatment groups, mean in vivo gene-marking rates (based on CD34 FACS analysis) reached 28.4% (range, 18.7–46.2%) for TK.007 and 23.3% (range, 16.3–30.5%) for scHSVtk.
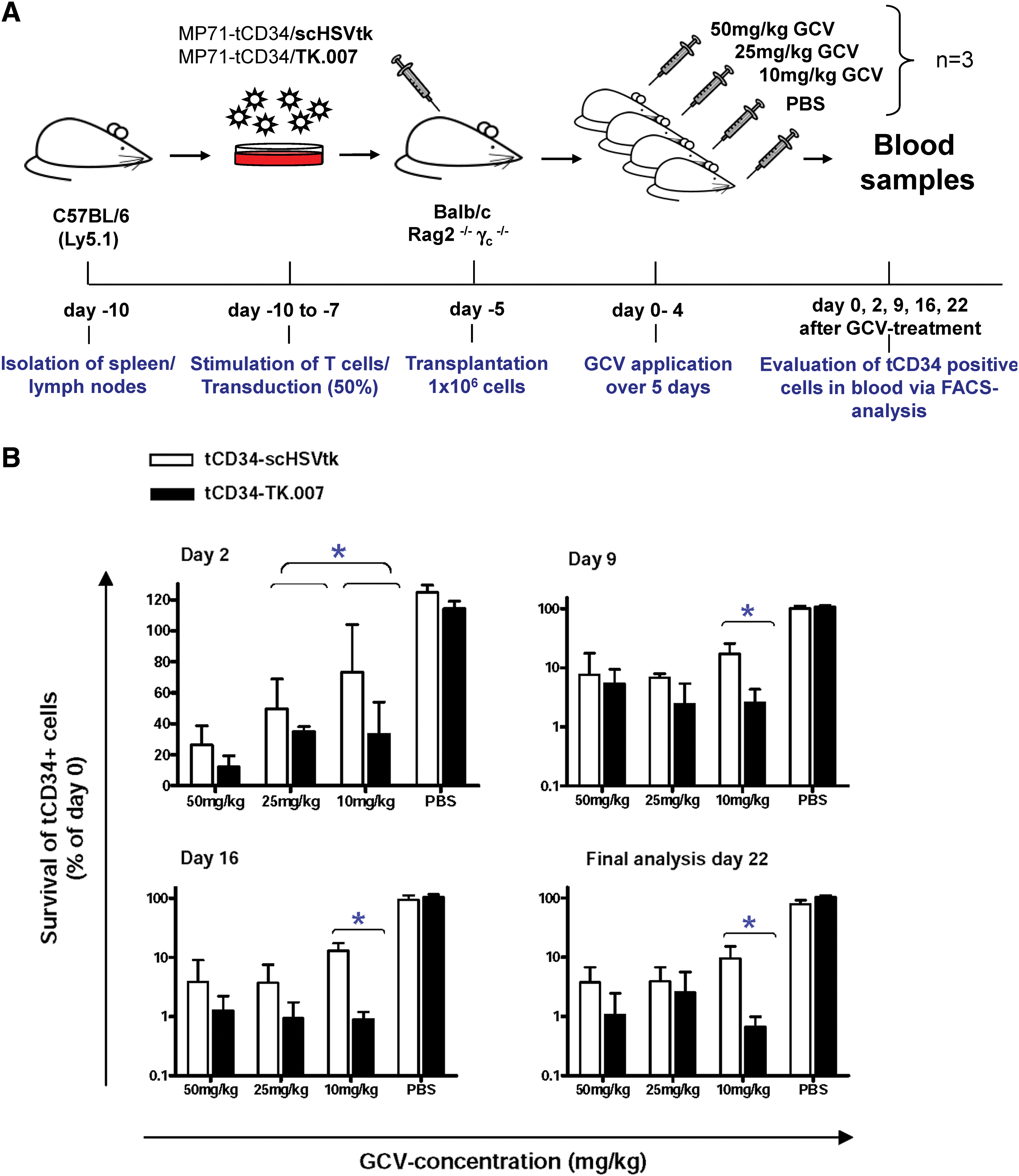
TK.007 shows superior performance as compared with scHSVtk in a quantitative mouse model in vivo. (
After engraftment, mice transplanted with either of the TK variants were subdivided into three treatment groups (three animals per group): group 1, GCV at 50 mg/kg daily; group 2, GCV at 25 mg/kg daily; and group 3, GCV at 10 mg/kg daily. A potential influence of the treatment regimen or the vectors used on long-term engraftment was excluded by performing a control experiment. Here, mice transplanted in parallel with cells transduced with either MP71-tCD34/TK.007 or MP71-tCD34/scHSVtk (n = 3 per vector) were treated with PBS instead of GCV.
For the three GCV groups and the one control group (per vector), treatment was initiated 5 days after T cell infusion. GCV (in the control group, PBS) was administered on five consecutive days. To measure the effects of the various GCV doses in relation to the two tested HSVtk variants, numbers of CD34+ cells in the peripheral blood of each individual mouse were determined at various time points (Fig. 5A). On the basis thereof, relative survival rates of CD34+ cells were estimated for the two suicide genes at all available time points (Fig. 5B).
As already seen in vitro, TK.007 conferred faster killing kinetics as evident from higher killing rates 2 days after GCV initiation (Fig. 5B). Notably, despite the limited numbers of animals, for the two lower GCV concentrations the observed differences reached significance. Moreover, subsequent analyses at various time points after GCV treatment indicate efficient elimination (approximately 99%) of TK.007-transduced cells independently of the applied GCV dose (Fig. 5B). In striking contrast, at low GCV (10 mg/kg), a considerable proportion (>10%) of scHSVtk-transduced cells escaped GCV-mediated depletion. As shown in Fig 5B, in each single analysis after GCV treatment (independent of the applied dose) we found higher absolute killing rates for TK.007-transduced versus scHSVtk-transduced cells. Importantly, for GCV at 10 mg/kg the observed differences were always significant (Fig. 5B). Taken together, these data emphasize that TK.007 mediates not only faster killing, but also higher absolute sensitivity toward GCV, particularly at lower drug doses.
Discussion
Suicide genes that kill cells on external activation have become useful tools in gene and cell therapy. They are introduced into cells (1) with the aim of immediate killing of the targeted cells in vivo, most commonly cancer or cancer-supporting stromal cells or (2) as a safety measure providing the opportunity to eliminate infused cells in case they would cause harmful side effects. Moreover, given the occurrence of severe side effects in clinical gene therapy studies aiming at the treatment of monogenic inherited diseases, inclusion of suicide genes into integrating vectors is being discussed as a potential safety measure (Baum and Fehse, 2003).
HSVtk is one of the most extensively evaluated transgenes in gene therapy. This broad application of HSVtk as a suicide gene has various reasons: (1) the gene has already proven its general safety in vivo; (2) GCV is a commonly used antiviral drug with a well-established safety profile even in the transplant setting; (3) the mechanism of GCV toxicity results in the almost exclusive killing of dividing cells, thus providing an additional specificity for alloreactive T cells (in the case of the adoptive immunotherapy approach) or tumor cells when surrounded by nondividing tissues (such as the brain); and (4) in solid tumors, the HSVtk/GCV system is characterized by a pronounced bystander effect (Dilber et al., 1997).
At the same time, the use of HSVtk as a suicide gene is associated with several drawbacks (see the Introduction) that have led to numerous efforts to improve the system or to introduce alternative suicide/prodrug combinations. In this study, we evaluated novel HSVtk gene variants. Therefore we synthesized a fully novel gene variant with optimized codon usage to overcome gene expression problems and potential toxicity associated with its viral gene sequence. In a second step we introduced into this coHSVtk a mutation (A168H) previously described by Karlsson and colleagues (Balzarini et al., 2006), which results in strongly reduced affinity for endogenous thymidine. The resulting TK variant represented the seventh TK gene tested in our laboratories and was therefore named TK.007.
We extensively tested the novel TK variants in the setting of hematopoietic cells in general and lymphocytes in particular, regarding their potential use in adoptive immunotherapy. We found strongly increased expression of the novel codon-optimized A168H mutant TK.007 in both myeloid and lymphoid cells as compared with splice-corrected wild-type HSVtk (Chalmers et al., 2001). Codon optimization alone resulted in an intermediate performance. Notably, higher expression of TK.007 as compared with wild-type HSVtk was conserved during long-term observation (12 weeks) in nonselected cell cultures, which indicates lower toxicity of the A168H mutant in comparison with wild-type HSVtk. This apparently lower toxicity may be important for in vivo applications, because it should reduce the selective pressure against cells expressing functional TK. Indeed, preferential survival of clones bearing nonfunctional HSVtk copies has been described in the long-term follow-up of gene therapy patients (Deschamps et al., 2007). The latter has been attributed to immune-mediated rejection of TK-expressing clones (Deschamps et al., 2007). However, a direct toxic impact of HSVtk expression could not be excluded, in particular because immune responses were not detected in all patients (Deschamps et al., 2007). Moreover, given our observation that high HSVtk expression is much better tolerated in the case of the novel TK.007 gene, one might well speculate that the use of the latter will also allow the achievement of better vector titers in permanent vector producer cell lines.
Using different in vitro readout systems, we next showed that TK.007 was also functionally superior to both the splice-corrected and codon-optimized HSVtk proteins. In fact, primary T lymphocytes as well as several T cell lines and one myeloid cell line transduced with TK.007 showed improved killing kinetics after GCV administration compared with cells transduced with the control vector(s). Notably, nearly identical results were independently obtained in different laboratories using gammaretroviral vectors encoding two different sort-suicide genes—tCD34/HSVtk (Fehse et al., 2002) and HSVtk-OuaS, a novel fusion protein of HSVtk with the OuaSelect marker (Treschow et al., 2007). This emphasizes that the better functionality of TK.007 is not context or vector dependent. More rapid elimination of alloreactive T cells is highly relevant in the adoptive immunotherapy setting, because the time point of GCV administration would be at the manifestation of severe GvHD.
In a next step we investigated the potential of TK.007-transduced cells to mediate severe allo-GvHD in vivo, in the mouse model. Genetic modification of T lymphocytes was shown to significantly impair their functional capabilities (Robinet et al., 2005). Moreover, a potential impact of the tCD34/TK.007 fusion suicide gene used on T cell activity in vivo could not per se be excluded. However, we found induction of severe, lethal GvHD after infusion of T cells transduced with either MP71-tCD34/TK.007 or MP71-tCD34/scHSVtk. GvHD was readily abrogated in six of six TK.007 mice and in five of six scHSVtk mice by short-term (3 days) application of ganciclovir at a dose (10 mg/kg) that was five times lower than the dose most often used in murine studies. The high frequency of severe GvHD relapses indicates that in two thirds of the animals the 3-day treatment course at such a low dose was not sufficient to eliminate all alloreactive T cells. On the other hand, continuous complete remissions were found with this regimen in two of six animals (at least within the observation time), emphasizing the principal possibility to decrease both GCV dose and treatment duration. Given the reported myelotoxicity of GCV (Biron, 2006) the possibility to reduce GCV doses might be particularly important in clinical hematopoietic stem cell transplantation (HSCT) settings.
Although the previously described model proved the potential of TK.007 in abrogating GvHD, it did not allow quantitative comparison of the novel versus the well-established scHSVtk variant. Therefore we established a modified T cell transplantation model that facilitated direct in vivo comparison of killing kinetics and absolute killing rates mediated by the various suicide genes. On the basis thereon we were able to provide strong in vivo evidence that TK.007 shows significantly improved characteristics regarding (1) GCV sensitivity, (2) kinetics, and (3) absolute efficacy of GCV-mediated killing. All three aspects are of the utmost importance for the adoptive immunotherapy field, because severe acute GvHD requires the fastest and most comprehensive effectiveness of any treatment regimen (Bonini et al., 1997; Tiberghien et al., 2001), whereas a potential dose reduction and/or a shorter application time of GCV would decrease unwanted side effects resulting from its myelotoxicity (Biron, 2006).
The molecular basis for the superior performance of the codon-optimized A168H HSVtk mutant TK.007 as a suicide gene needs to be investigated in greater detail. The higher expression levels in cells observed in this study could be interpreted as resulting from lower unspecific toxicity, although it is in principle possible that the A168H amino acid substitution affects mRNA or protein stability. The higher enzyme levels achieved with the A168H mutation would facilitate higher GCV turnover. In addition to the higher catalytic efficiency of the enzyme (Balzarini et al., 2006), the required threshold for apoptosis initiation may therefore be reached more rapidly and at lower GCV concentrations. The decreased affinity of TK.007 for endogenous thymidine would also result in lower substrate competition between ganciclovir and cellular nucleosides. Moreover, higher unspecific toxicity of wild-type HSVtk may negatively influence cell cycle rates and metabolic activities of transduced cells, thereby indirectly decreasing their sensitivity toward GCV toxicity.
In conclusion, we have shown that the novel suicide gene TK.007 shows better functionality (improved GCV sensitivity and killing rate) both in vitro and in vivo. Better performance of TK.007 may at least partly be due to its lower unspecific toxicity, allowing higher intracellular enzyme levels and sustained long-term expression. On the basis of these data we believe that TK.007 represents an attractive candidate suicide gene. In particular, the two new sort-suicide genes combining advantages of TK.007 with tCD34 or OuaSelect should be well suited for genetic modification of lymphocytes in adoptive immunotherapy settings. At the same time, our preliminary data (not shown) indicates that TK.007, on the basis of its superior performance, also represents a promising alternative for cancer suicide gene therapy.
Footnotes
Acknowledgments
This work is part of the doctoral theses of Ellen Preuß and Alexandra Treschow. The authors are grateful for the kind support provided by the Frankfurter Stiftung für krebskranke Kinder, the Gutermuth-Stiftung, the Deutsche Forschungsgemeinschaft (FE 568/11-1), Clinigene-NoE EU grant LSHB-CT-2006-018933, and the Swedish Childhood Cancer Foundation. The authors also thank Mehmet Uzunel for generously providing GAPDH primers and probes and Rolf Marschalek for critical reading of the manuscript.
Author Disclosure Statement
No competing financial interests exist.