
Abstract
The limited efficacy of adenovirus type 5 (Ad5)-based oncolytic viruses seen in the clinic thus far may be attributable in part to variable expression of its receptor on tumor cells. Replacement of the Ad5 fiber knob with the Ad35 fiber knob generated the Ad5/35 chimeric virus, which has previously been demonstrated to have significant antitumor activity in murine tumor models, presumably by virtue of its recognition of the CD46 receptor, which is abundant on many types of tumor cells. In the current study, a CD46 receptor transgenic mouse strain (hCD46Ge) that expresses the CD46 receptor in a pattern closely mirroring that in humans was used to study the in vivo properties of Ad5/Ad35 chimeric viruses. Vector distribution was evaluated after intravenous administration to hCD46Ge mice of an Ad5-based oncolytic adenovirus or an Ad5/35 chimeric oncolytic adenovirus (designated OV-5 and OV-5T35H, respectively), a wild-type Ad5 virus (Ad5wt), or an Ad5-based, E1-deleted adenovirus (Addl312) at 1.25 × 1012 viral particles/kg. The amount of OV-5T35H vector genomes in the liver was at least two orders of magnitude lower than that of Ad5-based viruses. Moreover, animals injected with OV-5T35H virus had significantly lower elevations of serum proinflammatory cytokines and liver enzyme levels. Mice injected with Ad5wt lost more than 20% of their body weight and died or required euthanasia because of poor clinical condition within 4 days of virus administration. Mice treated with OV-5 lost as much as 15% of their body weight over 8–9 days, but recovered within 14 days. Mice that were treated with Addl312 or OV-5T35H exhibited no body weight loss during the study period. These studies suggest that the Ad5/35-based chimeric viruses may have a better safety profile after intravenous injection compared with Ad5-based viruses.
Introduction
The therapeutic profile of the Ad5-based viruses observed thus far has been attributed to the low and variable expression levels of the coxsackievirus–adenovirus receptor (CAR), the cellular receptor used by the Ad5 virus, on tumor cells (Bergelson et al., 1997; Miller et al., 1998; Li et al., 1999; Okegawa et al., 2000; Reddy et al., 2006). CAR expression is also lost with progression of certain tumors (Yamamoto et al., 1997). To overcome this limitation, adenoviruses can be engineered to infect cells by using alternative receptors that are more abundantly expressed (Nilsson et al., 2004; Stone and Lieber, 2006). Because expression of CD46 is relatively high on tumor cells, a retargeted vector that uses CD46 as a cellular receptor should increase the level of tumor cell transduction by the virus. It has been demonstrated that switching the Ad5 fiber knob with the Ad35 knob, which recognizes the highly abundant CD46 receptor on tumor cells, significantly improves viral entry and correlates with enhanced antitumor activity of Ad5/35 chimeric oncolytic viruses (OV-5T35H) compared with Ad5-based oncolytic viruses (OV-5) (Reddy et al., 2006; Ganesh et al., 2007).
Although oncolytic Ad5-based viruses are reasonably well tolerated when injected directly into tumors, toxicity associated with nononcolytic Ad5 administration has been observed at high systemic doses and may include complement activation, cytokine release, and consequent vascular damage leading to systemic inflammatory responses. The acute inflammation occurs within 24 hr of virus administration and is induced primarily by viral capsid interactions with the host cells. The next phase of virus-mediated toxicities begins 3–4 days after Ad5 administration and requires expression of viral gene products. The uptake of systemically delivered Ad5 by its target cells induces elevation of liver transaminases, tissue damage, and production/release of cytokines and chemokines, which in turn play a major role in inflammation and the induction of an antiviral adaptive immune response (Lieber et al., 1997; Muruve et al., 1999; Muruve, 2004; Shayakhmetov et al., 2005). Kupffer cells are thought to be the primary cellular source of these cytokines and chemokines (Lieber et al., 1997).
The Ad5/35 chimeric adenoviruses described previously cannot readily be studied in normal (nontransgenic) mice, because expression of the murine CD46 receptor homolog is restricted to the testis in mice (Cervoni et al., 1992), and there is no homology between the mouse and human cytoplasmic domain sequences of the CD46 receptor (Russell, 2004). In contrast, all human nucleated cells express CD46. However, the CD46 receptor expression pattern in CD46 transgenic mice is similar to that in humans, although there is some variability in expression levels among the three different CD46 transgenic mouse strains that are available (Kemper et al., 2001). After a preliminary evaluation of the three transgenic strains, the hCD46Ge strain, which has CD46 receptor RNA and protein expression levels comparable to those observed in matched human tissues (Mrkic et al., 1998; Kemper et al., 2001), was selected for biodistribution and pilot safety studies of OV-5T35H.
After systemic delivery of a single dose of each virus into hCD46Ge mice, OV-5T35H was found to be significantly less hepatotoxic and to cause less body weight loss than conventional Ad5-based vectors. The amount of OV-5T35H vector genomes in the liver of CD46 transgenic mice was at least two orders of magnitude lower than that of the Ad5-based viruses after systemic administration. Elevation of serum proinflammatory cytokines and liver enzymes was significantly lower in OV-5T35H-treated mice than in mice that received OV-5 or wild-type Ad5 virus (Ad5wt). Body weights of mice that received Ad5wt or OV-5 were decreased by approximately 15–20%, and many treated animals died or were killed during the study period. In contrast, all mice that received OV-5T35H survived for the duration of the study with no body weight loss. These studies demonstrate that the Ad5/35-based chimeric viruses are better tolerated after intravenous injection than are Ad5-based viruses in a murine model that expresses CD46 in a pattern that is similar to the pattern found in humans.
Materials and Methods
Nonreplicating and replicating viruses used in study
Ad5/35GFP is a nonreplicating virus in which a green fluorescent protein (GFP) reporter gene under the control of a strong constitutive cytomegalovirus (CMV) promoter was incorporated into the E1 region of an adenoviral vector that contained the fiber knob derived from Ad35 and the shaft from Ad5 adenovirus, the construction of which was described previously (Reddy et al., 2006). OV-5 and OV-5T35H are conditionally replication-competent adenoviruses also described previously (Reddy et al., 2006). In these viruses, the native E1A promoter is replaced by the tumor-specific E2F-1 promoter to restrict viral replication to cells that have defects in the pRb pathway (Hallenbeck et al., 1999; Jakubczak et al., 2003). In addition, the cDNA encoding human granulocyte-macrophage colony-stimulating factor (hGM-CSF) replaced the open reading frame encoding the 19-kDa glycoprotein of the E3 region (Ramesh et al., 2006). With the aim of improving viral entry and spread throughout the tumor, the Ad5 fiber knob of OV-5 was replaced with the Ad35 knob to generate OV-5T35H. Other adenoviruses used in the study include wild-type Ad5 and Addl312. Addl312 is a replication-defective vector with a deletion in the E1A gene that is used as a negative control (Reddy et al., 2006). High-titer viral stocks of replicating and nonreplicating viruses were generated (7–8 × 1012 viral particles [VP]/ml) in AE1-2a cells with a particle-to-plaque-forming unit (PFU) ratio in the range of 15–20 for all viruses.
CD46 receptor transgenic mouse strain
The CD46 receptor transgenic mouse strain (hCD46Ge) was obtained from the Mayo Clinic (Rochester, MN) (Kemper et al., 2001). The hCD46Ge strain was originally homozygous, but for ease of breeding, the mice were bred with C57BL/6NTac normal mice to generate 100% heterozygous, CD46 receptor transgenic mice. Both homozygous and heterozygous mice were genotyped after the first round of breeding to confirm that they had the correctly sized band when tested by polymerase chain reaction (PCR). These mice were then characterized for CD46 receptor levels on blood cells and these levels were compared with the levels present in human donor blood. The hCD46Ge strain was well characterized previously for CD46 receptor levels on blood cells as well as in various tissues (Kemper et al., 2001).
Isolation of white blood cells and erythrocytes
To isolate mouse or human erythrocytes (E), 4 ml of Histopaque 1119 (Sigma-Aldrich, St. Louis, MO) was overlaid with 4 ml of Histopaque 1077 (Sigma-Aldrich) in a 15-ml centrifuge tube. Heparinized whole mouse or human blood (200 μl) was added onto the upper gradient of the tube and the erythrocytes were isolated according to the manufacturer's protocol. To isolate white blood cells from mouse or human blood, 200 μl of whole blood was lysed for 10 min on ice in 10 ml of erythrocyte lysis buffer. The samples were centrifuged for 5 min at 800 × g at 4°C, washed once with 10 ml of erythrocyte lysis buffer, and then resuspended in the appropriate buffer for subsequent experiments.
Flow cytometry
To analyze CD46 receptor expression by flow cytometry (FACS), murine or human erythrocytes (5 × 105 cells) were incubated on ice with fluorescein isothiocyanate (FITC)-labeled anti-human CD46 antibody (169-1-E4.3, 1:200 dilution in phosphate-buffered saline [PBS]; Axxora, San Diego, CA) plus 3% fetal bovine serum (FBS) for 1 hr in a total volume of 100 μl. The cells were washed three times with PBS–FBS, resuspended in PBS, and analyzed by flow cytometry (FACScan; BD Biosciences, San Jose, CA) for CD46 receptor expression levels. To evaluate CD46 receptor expression levels on T lymphocytes, 2 × 105 cells were incubated with phycoerythrin (PE)- or allophycocyanin (APC)-labeled mouse/human anti-CD4 or anti-CD8 specific monoclonal antibodies and FITC-labeled human anti-CD46 receptor antibodies for 1 hr on ice in 100 μl of PBS–FBS. Samples were washed three times with PBS–FBS, resuspended in PBS, and analyzed by multichannel flow cytometry.
In vivo biodistribution studies
hCD46Ge mice (homozygous) were bred with C57BL/6NTac mice to generate heterozygous mice that were used for the studies described in this paper. In the first study, 50 heterozygous hCD46Ge mice and 50 C57BL/6NTac mice were randomized into groups (n = 10 per group). Normal and hCD46Ge mice received intravenous injections of PBS, Ad5wt, Addl312, OV-5, or OV-5T35H at a single dose of 1.2 × 1012 VP/kg in a volume of 10 ml/kg, which is the maximal tolerated dose of the Ad5wt virus. Blood samples were drawn 6 hr, 48 hr, 72 hr, and 14 days after virus injection to monitor serum levels of proinflammatory cytokines and liver enzymes and for complete blood count (CBC) analyses. Whole blood was also collected 2 and 48 hr after virus injection to evaluate virus binding to blood cells. Three and 14 days after vector administration, 5 mice from each group were killed and 10 major organs (liver, spleen, lung, kidney, heart, pancreas, brain, lymph nodes, ovary, and colon) were collected from each mouse and samples for DNA analysis were snap frozen in liquid nitrogen.
In the second study, six homozygous hCD46Ge mice and six C57BL/6NTac mice were placed on study (n = 3 per group). Animals were randomized on the basis of individual body weight. Both hCD46Ge mice (n = 3) and normal mice (n = 3) were injected intravenously at 5 × 1012 VP/kg with either nonreplicating Ad5GFP or Ad5GFP-5T35H virus. As stated previously, this is the maximal tolerated dose for the viruses. Mice were bled 2, 6 and 72 hr after virus injection for collection of whole blood and serum. Seventy-two hours after virus administration, the mice were killed and tissues were collected from the same 10 organs. Half the tissue samples were fixed in 4% paraformaldehyde (PFA) to evaluate GFP expression by immunohistochemistry (IHC) and the remaining samples were snap frozen for DNA analysis.
Quantitative PCR for vector genomes
Tissues from mice were harvested at several time points after virus injection and frozen. The genomic DNA was isolated from 10 to 20 mg of tissue or whole blood, using a DNeasy kit (Qiagen, Valencia, CA) according to the manufacturer's protocol. Genomic DNA (∼500 ng) was used for real-time PCR, using vector-specific primers against Ad5 hexon (Hexon For, 5′-CTT CGA TGA TGC CGC AGT G-3′; Hexon Rev, 5′-GGG CTC AGG TAC TCC GAG G-3′; probe, 5′/56-FAM/TTA CAT GCA CAT CTC GGG CCA GGA C/36-TAMAp/-3′) and against the endogenous mouse glyceraldehyde-3-phosphate dehydrogenase (GAPDH) gene as an internal standard to equalize numbers of vector genomes between different tissues. DNA was isolated from whole blood samples, using the DNeasy kit as described previously. The adenoviral DNA content in the blood was determined by real-time PCR as previously described.
Immunohistochemical analyses
Organs were collected and left for 3 hr in 4% PFA in PBS for complete fixation. The samples were transferred to a 30% sucrose solution for cryoprotection and left overnight at 4°C. Samples were rinsed in PBS and blotted dry, and sausage blocks of respective tissues were embedded in O.C.T. medium (Statlab Medical Products, McKinney, TX) before freezing in a methyl butane–dry ice bath. Five-micrometer-thick serial sections were directly analyzed for GFP expression by fluorescence microscopy. Serial sections were collected, air dried, and stored at −20°C for subsequent IHC. Blood vessels were identified with a rat anti-mouse endoglin antibody (CD105) (R&D Systems, Minneapolis, MN).
Blood and serum analyses
A serum chemistry panel and a CBC were performed at IDEXX Laboratories (Westbrook, ME). The level of serum cytokines was measured with a mouse enzyme-linked immunosorbent assay (ELISA) kit specific for mouse IL-6 and tumor necrosis factor (TNF)-α (BD Biosciences).
Results
Characterization of hCD46Ge mice
To effectively study the biological activity of the Ad5/35 fiber-chimeric virus (OV-5T35H), a murine model that expresses the CD46 receptor in a pattern closely mirroring that in humans must be used. The hCD46GE strain that has been selected for this study has previously been shown to express CD46 receptor levels in peripheral blood similar to those found in human blood (Kemper et al., 2001). In addition, the study showed that CD46 receptor RNA and protein expression levels in organs/tissues of hCD46Ge mice are comparable to those found in matched human tissues, a finding that was supported by immunohistochemical analyses (Mrkic et al., 1998; Kemper et al., 2001).
We further evaluated homozygous and heterozygous hCD46Ge mice for CD46 expression levels on blood cells and compared them with CD46 receptor levels found in human donor blood, because virus injected intravenously will bind to CD46 receptor expressed by blood cells and thus CD46 receptor levels on blood cells that differed from the CD46 receptor levels found in human blood could strongly impact biodistribution studies. In homozygous mice, the CD46 receptor levels on CD4+ and CD8+ T lymphocytes were comparable to the levels found on human lymphocytes as shown by fluorescence-activated cell-sorting (FACS) analysis (Fig. 1A and C; mean fluorescence, 54 and 61 and 59 and 62.7, respectively) and the CD46 receptor levels on lymphocytes of heterozygous hCD46Ge mice were, as expected, about half the levels present in homozygous mice (Fig. 1B; mean fluorescence, 37 and 33, respectively). Although the CD46 expression levels in lymphocytes of homozygous mice were close to the levels found in human lymphocytes, they were slightly higher in the red blood cells (RBCs) of homozygous mice than in heterozygous mice or in human donor blood (Fig. 1A; mean fluorescence, 2.5, 1.24, and 0.2, respectively). The slightly higher CD46 receptor levels detected in homozygous compared with heterozygous mice, however, did not make any difference in the hemagglutination affinity for Ad5/35 virus (data not shown). Ad5/35 virus only marginally hemagglutinated the blood from both strains (data not shown). As expected, Ad5 and Ad5/35 viruses did not hemagglutinate normal mouse blood, as these RBCs express neither CAR nor human CD46 (data not shown). Interestingly, the Ad5/35 chimeric virus did not hemagglutinate human blood whereas the Ad5 virus marginally did, because the RBCs in human blood do not express CD46 but CAR (data not shown). Although the difference in CD46 levels found between the two strains led to only a marginal difference in hemagglutination affinity, we used primarily the heterozygous mice in the biodistribution studies to avoid sequestration of incoming virus (if at all present) by the RBCs in homozygous mice in excess of what would be expected in humans.
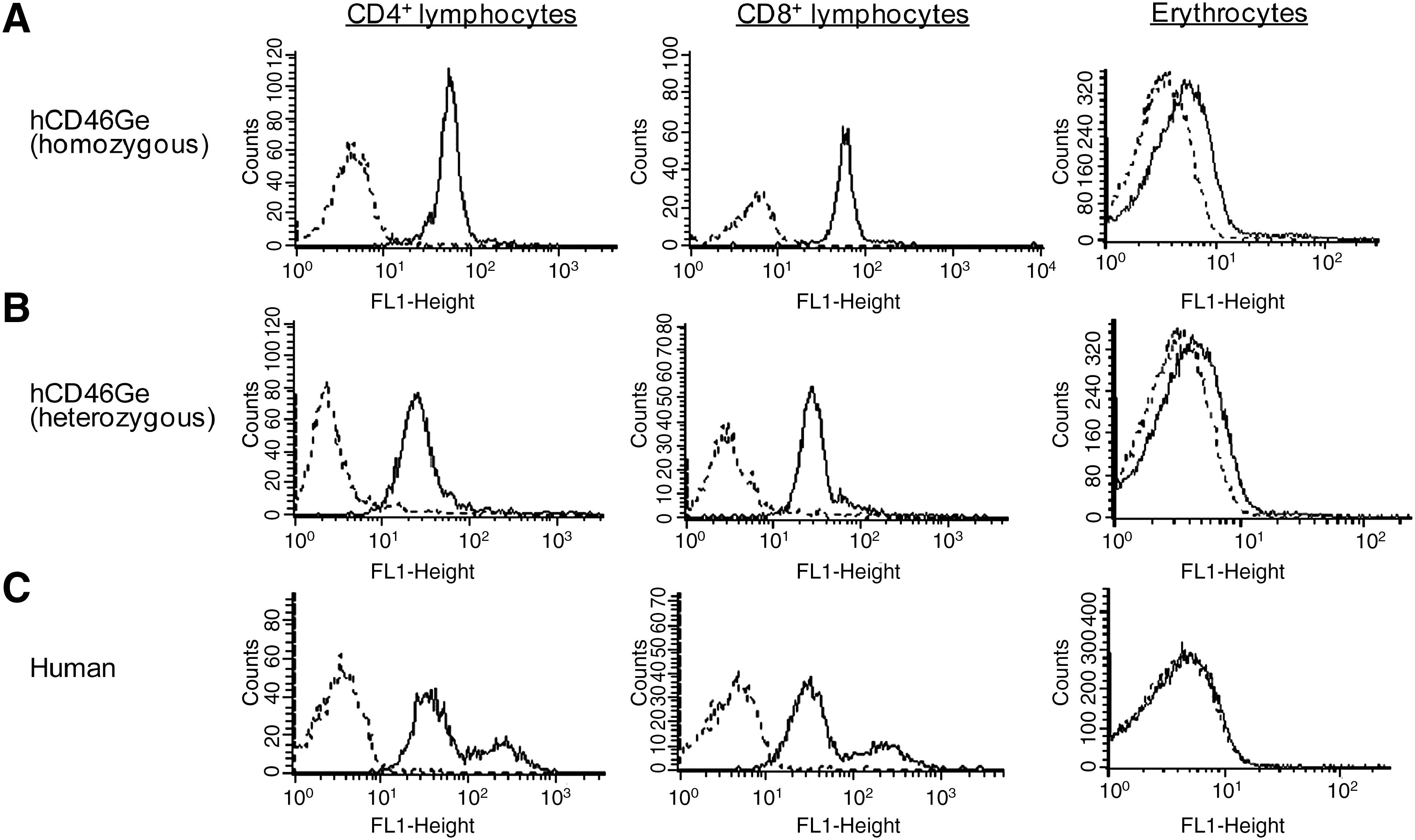
CD46 receptor levels on peripheral blood cells (CD4+, CD8+ T lymphocytes and erythrocytes) of CD46-receptor transgenic mice (hCD46Ge) or human donor blood. White blood cells and erythrocytes were purified from whole blood of either (
The chimeric oncolytic virus, OV-5T35H, demonstrated lower liver transduction and less overall toxicity in hCD46Ge (heterozygous) mice than did Ad5-based viruses. To understand the biodistribution and safety profile of the Ad5/35 fiber-chimeric virus, OV-5T35H, OV-5, Ad5wt, and Addl312 viruses were tested in normal and heterozygous hCD46Ge mice. Ad5wt virus was included in the study to show that the selectively replicating oncolytic adenovirus (in which the tumor-selective promoters control the E1A expression that is required for replication only in tumor cells)-mediated toxicity is less than that of wild-type virus. Addl312 was included in the study not to be compared side by side with the oncolytic viruses but to serve as a “negative control for toxicity” for the selectively replicating oncolytic viruses.
Normal and hCD46Ge mice (n = 10) each received one of the viruses at 1.25 × 1012 VP/kg, and body weights and overall condition were monitored over 14 days. Three days after virus injection, liver, spleen, lung, kidney, heart, pancreas, brain, lymph nodes, ovary, and colon from a selected set of mice were harvested and tissue samples were snap frozen for quantitative PCR (qPCR) analyses to determine the viral genome distribution. Treatment of both normal and hCD46Ge mice with Ad5wt caused a body weight reduction of more than 20%, and all treated mice in both groups died or were euthanized due to poor clinical conditions (Fig. 2A and B) within 4 days of treatment. Normal and hCD46Ge mice treated with OV-5 lost up to 15% of their body weight in the 8 days after viral injection, but most recovered to their start weight by day 14 after virus injection (21.1 ± 2.5 g). One normal mouse treated with OV-5 was euthanized due to extreme weight loss 4 days postinjection. In contrast, mice injected with OV-5T35H or Addl312, and the control mice injected with PBS, had no significant body weight loss during the 14-day duration of the study (Fig. 2A and B), suggesting that these viruses are better tolerated in normal and hCD46Ge mice.
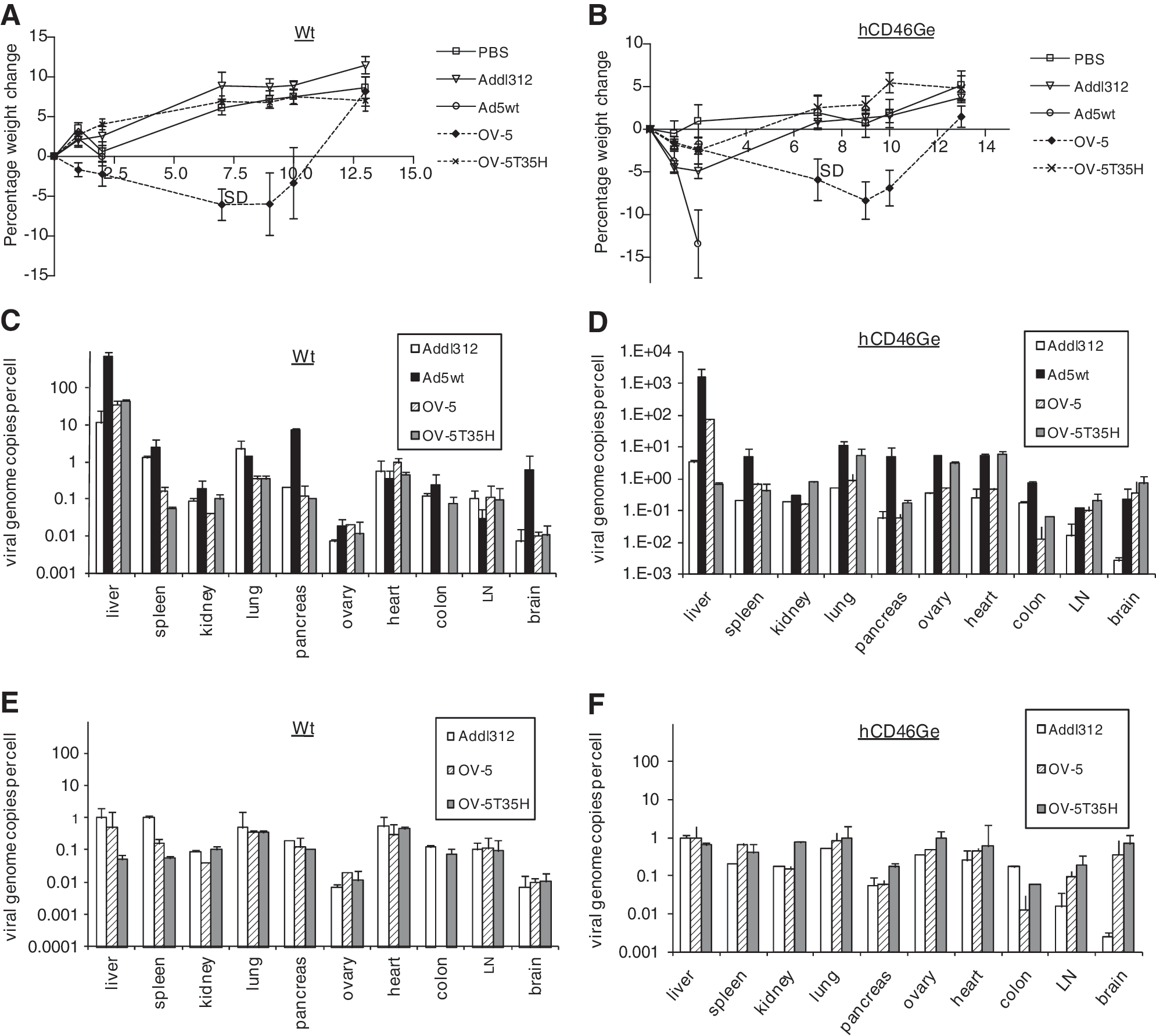
Biodistribution of adenoviral variants in normal and heterozygous hCD46Ge mice. Mice (n = 5) were injected intravenously with PBS, an E1-deleted Ad-5-based control virus (Addl312), Ad5 wild-type virus (Ad5wt), Ad5-based oncolytic virus (OV-5), or Ad5/35 fiber-chimeric oncolytic virus (OV-5T35H) at a dose of 1.2 VP/kg. (
qPCR analysis showed that heterozygous hCD46Ge mice injected with Ad5wt had about 1000 viral genomes per hepatocyte 3 days after virus injection (Fig. 2D). Liver of hCD46Ge mice injected with OV-5 had only 100 viral genomes per cell on day 3, and mice treated with OV-5T35H had < 1 viral genome per hepatocyte (Fig. 2D), suggesting that the chimeric oncolytic virus transduced liver of hCD46Ge mice poorly. Liver of Addl312-treated hCD46Ge mice had about 2 or 3 viral genome copies per cell, emphasizing again the low transduction efficiency of hepatocytes by the chimeric virus. Although Ad5wt and OV-5 use the same receptor for cell entry, about 10-fold more viral genomes were detected in the hepatocytes of Ad5wt-injected hCD46Ge mice than in hepatocytes of OV-5-injected mice, possibly reflecting DNA replication of the Ad5wt genome without forming infectious Ad5wt viral particles. Normal mice injected with OV-5 or OV-5T35H had similar viral copy numbers in their hepatocytes, suggesting that fiber-chimeric and parental Ad5-based oncolytic viruses enter liver cells equally well in the absence of the CD46 receptor (Fig. 2C). A similar number of viral genome copies was found in normal and hCD46Ge mice injected with Addl312 (2 or 3 copies per cell). Normal mice injected with Ad5wt had, as observed in hCD46Ge mice, an approximately 10-fold higher number of viral genomes in the liver compared with the liver of normal mice injected with either OV-5 or OV-5T35H (Fig. 2C), again suggesting that Ad5wt might be replicating in the liver or that DNA replication of the Ad5wt genome occurred without forming infectious viral particles. OV-5 viral genome copies present in liver of both normal and hCD46Ge mice were also similar. Although the number of OV-5T35H genome copies present in the liver of normal mice is similar to the number of OV-5 genome copies found in normal and hCD46Ge mice, a significantly lower level of OV-5T35H viral genome copies was observed in the liver of hCD46Ge mice postinjection, suggesting that the number of chimeric viral particles that enter the liver of animals expressing the CD46 receptor at levels similar to those in human liver is lower than the number of chimeric viral particles entering the liver of normal mice. By study day 14, OV-5-treated normal or hCD46Ge mice had similar amounts of vector DNA in the liver as mice injected with Addl312 (∼2 copies per cell), and the viral genomes present in the liver of OV-5T35H-injected mice (in both mouse strains) were below 1 copy per cell, indicating that the majority of viral genomes in the hepatocytes were cleared by day 14 (Fig. 2E and F).
The qPCR data also showed that the number of viral genome copies was low (0.0001–10 copies per cell) for all the viruses in the other organs evaluated, including spleen, kidney, lung pancreas, ovary, heart, colon, lymph node (LN), and brain, compared with the number of copies in the liver (Fig. 2D). On study day 3, a 2- to 10-fold increase in the number of viral genome copies was detected in heart, lung, brain, kidney, and ovary of hCD46Ge mice treated with OV-5T35H compared with hCD46Ge mice treated with OV-5 (0.01–0.8 and 0.06–3.0 copies per cell, respectively, in OV-5- and OV-5T35H-treated mice). At the same time point Ad5wt-injected hCD46Ge mice had approximately 2- to 25-fold higher levels of viral genome copies compared with the OV-5T35H-treated mice. By day 14 after virus administration, the viral genome copy numbers for Addl312-, OV-5-, and OV-5T35H-treated hCD46Ge mice were reduced to less than 1 copy per cell in all organs tested, including liver, showing that the viral DNA had been cleared by that time. In contrast, normal mice injected with OV-5 and OV-5T35H had similar numbers of viral genome copies in all organs tested, and the Ad5wt-injected mice had a 10- to 50-fold greater number of genome copies (Fig. 2C). Increased viral genome copies were observed in all other organs besides liver in hCD46Ge mice injected with OV-5T35H when compared with normal mice injected with the same virus.
To evaluate the extent of virus binding to blood cells, whole blood was collected 2 and 48 hr after treatment for qPCR analyses of genomic DNA extracted from the blood cells. The number of viral genomes associated with the blood of heterozygous hCD46Ge or normal mice 2 and 48 hr after treatment was <1% of the virus input dose (data not shown), suggesting similar clearance of all viruses in both mouse strains and that no significant binding to erythrocytes occurred.
The efficiency of virus uptake by hepatocytes correlated with elevations in liver enzymes and serum cytokines. Six hours after virus administration, IL-6 levels in normal and hCD46Ge mice injected with Addl312, Ad5wt, or OV-5 were elevated and were 2- to 3-fold higher than in OV-5T35H-injected animals. This difference may reflect an acute inflammatory response to the Ad5-based viruses (Fig. 3A). Forty-eight hours after virus injection, IL-6 serum levels in mice injected with Addl312, OV-5, or OV-5T35H virus dropped by 3- to 15-fold whereas IL-6 levels in Ad5wt-injected mice remained elevated, further suggesting active DNA replication in the liver (Fig. 3B). TNF-α levels of mice injected with any of the viruses remained within the normal range at both time points (data not shown).
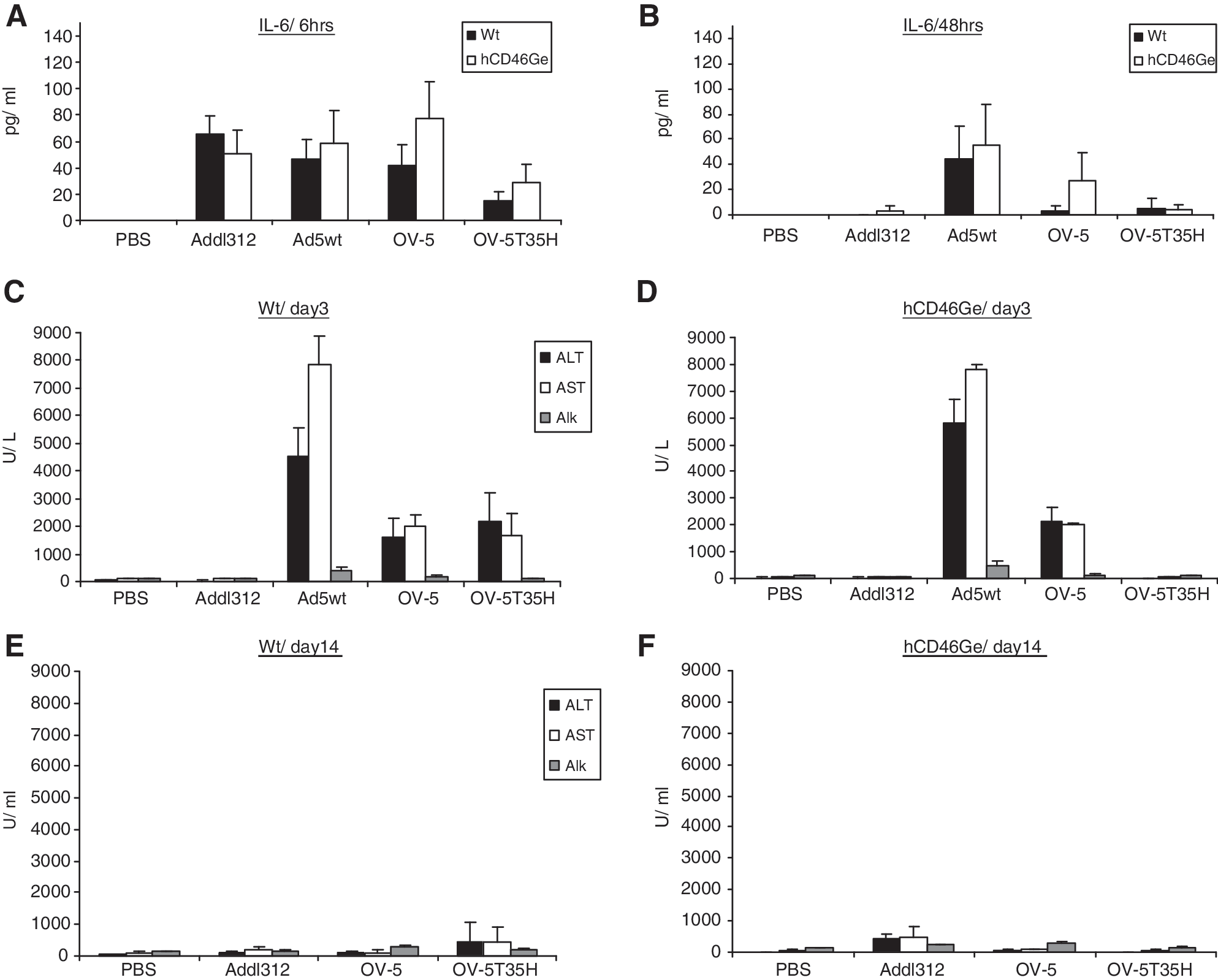
Analysis of selected clinical chemistry markers including ALT and AST, and markers of the innate immune response (IL-6), after injection of PBS (control), Addl312, Ad5wt, OV-5, or OV-5T35H into normal and hCD46Ge (heterozygous) mice at various time points. PBS, or Addl312, Ad5wt, OV-5, or OV-5T35H virus (1.25 VP/kg), was injected intravenously into normal or heterozygous hCD46Ge mice. IL-6 plasma levels were determined (
Serum chemistry analysis on day 3 in Ad5wt-injected hCD46Ge mice revealed significantly increased aspartate aminotransferase (AST) and alanine aminotransferase (ALT) levels (∼3- to 4-fold) compared with mice injected with OV-5 (p < 0.0001; Fig. 3C and D). However, ALT and AST levels in hCD46Ge mice injected with PBS, Addl312, or OV-5T35H were 25- to 100-fold lower than those in OV-5-injected mice (p < 0.0001) and 100- to 300-fold lower than those in Ad5wt-injected mice, suggesting that the OV-5T35H virus may be less hepatotoxic than the Ad5wt or OV-5 virus in transgenic hCD46Ge mice. Alkaline phosphatase (ALP) levels in OV-5T35H-injected hCD46Ge mice remained close to background levels (80 U/liter), whereas 2- to 5-fold higher levels were seen in mice treated with OV-5 (140 U/liter) and Ad5wt (477 U/liter) (Fig. 3D). Similarly elevated levels of AST, ALT, and ALP were seen in normal mice that received either OV-5 or OV-5T35H and were about 2- to 4-fold lower than those in mice injected with Ad5wt virus. Liver enzyme levels were elevated in both normal and hCD46Ge mice injected with Ad5wt or OV-5, although to a lesser extent with the latter virus. Normal mice treated with OV-5T35H had elevated liver enzyme levels similar to OV-5-injected mice, but liver enzyme levels remained at background levels in hCD46Ge mice treated with the fiber-chimeric virus. This observation correlates with the viral genome copy numbers found in the liver of these animals, suggesting a different liver uptake mechanism of fiber-chimeric viruses in the absence of CD46 receptors (Fig. 3C).
By study day 14, serum levels of AST, ALT, and ALP in mice injected with PBS, Addl312, OV-5, or OV-5T35H had returned to background levels (Fig 3E and F). No Ad5wt-treated mice survived to study day 14. On day 3, total bilirubins (TBili), direct bilirubins (DBili), hemoglobin, hematocrit (HCT), and RBC, neutrophil, and monocyte levels were elevated in both hCD46Ge and normal mice injected with Ad5wt compared with either mouse strain treated with the other viruses (Table 1). In contrast, lymphocyte and eosinophil levels were slightly decreased in both mouse strains treated with Ad5wt (at the same time point) compared with all other treatment groups (Table 1). However, the levels were within the control range of mice treated with OV-5 and OV-5T35H. Overall, these findings suggest that systemically administered fiber-chimeric oncolytic OV-5T35H virus may be less hepatotoxic in transgenic hCD46Ge mice than the in-class competitor Ad5-based oncolytic OV-5 virus as determined by changes in body weight, whole serum panel chemistries, and liver transduction efficiency.
Abbreviations: Abs, absolute; Alk, alkaline phosphatase; ALT, alanine aminotransferase; AST, aspartate aminotransferase; bili, bilirubin; HCT, hematocrit; HGB, hemoglobin; NRBCs, nucleated red blood cells; RBCs, red blood cells; WBCs, white blood cells.
PBS, or Addl312, Ad5wt, OV-5, or OV-5T35H virus (1.25 VP/kg), was injected intravenously into normal or heterozygous hCD46Ge mice. Three days later, whole blood was collected and complete blood count analysis was performed.
Ad5/35GFP virus-mediated transduction of blood endothelial cells in homozygous hCD46Ge mice
Vector biodistribution studies often make use of qPCR methodology to yield information about viral genome copy numbers. These assays are quantitative, but they do not provide information about the identity or location of transduced cells within the positive organs. To determine the location and identity of the virally transduced cells, GFP-expressing Ad5 parental and Ad5/35 chimeric viruses were injected intravenously into homozygous hCD46Ge and normal mice (5 × 1012 VP/kg). Three days after virus administration, liver, spleen, lung, kidney, heart, pancreas, brain, lymph nodes, ovary, and colon were collected and fixed with paraformaldehyde (PFA) to evaluate the distribution and fraction of GFP-positive cells. As expected from the preceding biodistribution data, the liver of Ad5GFP-injected hCD46Ge mice had higher numbers of GFP-expressing cells compared with the liver of Ad5/35GFP-injected hCD46Ge mice (Fig. 4A), but comparable numbers to those in the liver of normal mice. To determine the type of cells that were transduced by the chimeric virus, tissue sections of liver, heart, lung, brain, and ovary were stained for the endothelial-specific marker CD105. Endothelial cells (ECs) lining the blood vessels have previously been shown to express high levels of the CD46 receptor, suggesting that they are a likely cellular site for virus uptake (Stephen and Louise, 2002). GFP-expressing cells in the heart, lung, and ovary overlapped with the cells that stained positive for CD105, confirming that endothelial cells were transduced with the chimeric virus (Fig. 4B). In contrast, in the liver, the majority of cells that were transduced with Ad5 or the chimeric virus were hepatocytes in both normal and hCD46Ge mice (Fig. 4A). All the measurable transduction for Ad5 was detected mainly in liver and to a lesser extent in spleen. GFP signals were rather weak in other organs of mice transduced with Ad5GFP.
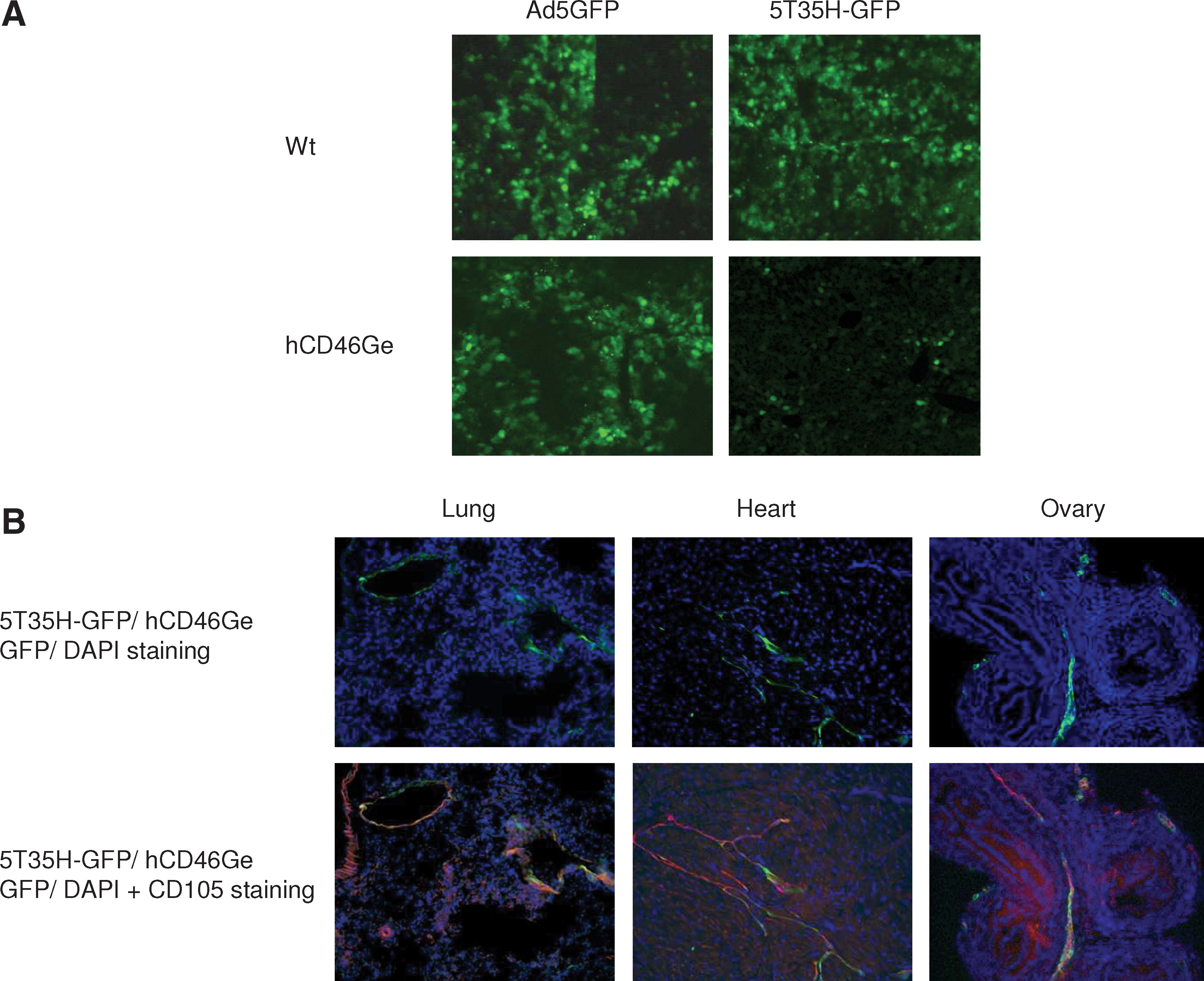
Biodistribution analyses of E1-deleted Ad5- and Ad5/35-based viruses expressing GFP in normal and homozygous hCD46Ge mice. Ad5GFP or 5T35H-GFP (5 × 1012 VP/kg) was injected intravenously into normal or hCD46Ge mice and selected organs were collected 3 days later. Liver (
Ad5/35 chimeric virus is better tolerated than Ad5-based virus in homozygous hCD46Ge mice
To determine whether the Ad5/35 chimeric virus would cause increased inflammatory cytokine or liver enzyme levels in homozygous CD46 transgenic mice, which express 2-fold higher levels of the CD46 receptor than do heterozygous mice, these mice (n = 6) and normal mice (n = 6) were injected intravenously at 5 × 1012 VP/kg with the E1-deleted viruses Ad5GFP and Ad5/35GFP. Blood was collected 2 hr after treatment to evaluate the blood clearance rate of the virus and 6 hr after virus administration to determine cytokine levels. Three days after treatment, serum was collected to measure liver enzyme levels and the same set of organs as listed previously were collected to determine distribution of viral genomes, using qPCR analyses.
IL-6 levels in hCD46Ge (homozygous) and normal mice treated with Ad5GFP were approximately 2.5- to 3-fold higher than those in Ad5/35GFP-injected mice 6 hr after treatment (Fig. 5A). TNF-α levels were elevated 20-fold in Ad5GFP-treated hCD46Ge mice compared with Ad5/35GFP-injected mice, but only about 2-fold in normal mice (Fig. 5B).
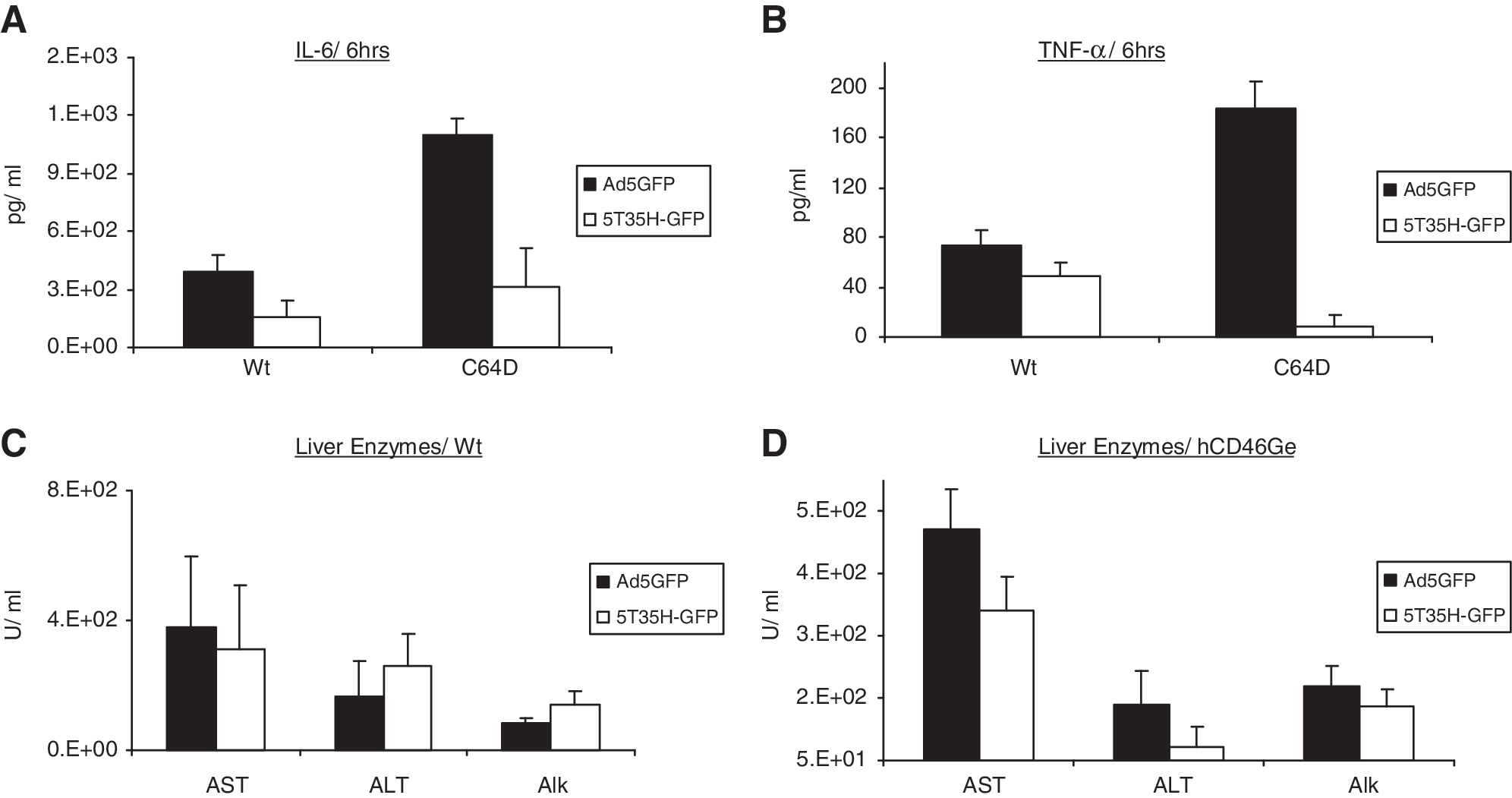
Evaluation of innate immune responses and liver enzymes after treatment with Ad5- and Ad5/35-based E1-deleted viruses expressing GFP. Ad5-GFP or 5T35H-GFP virus (5 × 1012 VP/kg) was injected intravenously into normal or homozygous hCD46Ge mice and plasma levels of (
Serum transaminase levels were 1.5- to 2-fold higher in hCD46Ge mice treated with Ad5GFP than in those that received Ad5/35GFP (Fig. 5D). The liver enzyme levels were similar in normal mice that received either Ad5GFP or Ad5/35GFP virus (Fig. 5C), as also seen in heterozygous hCD46Ge mice. The data suggest that the toxicity observed in heterozygous and homozygous hCD46Ge mice transduced with the various viruses was similar.
It was also noted that the liver enzyme levels in both wild-type and CD46 mice injected with OV-5 were about 4-fold higher than in wild-type and CD46 mice injected with a five times higher dose of nonreplicating Ad5GFP virus, suggesting that the oncolytic nature of the virus contributes to the overall toxicity. The same trend was observed in wild-type mice injected with OV-5T35H compared with Ad5/35GFP.
qPCR analyses showed that liver of homozygous hCD46Ge mice injected with Ad5/35GFP had approximately 20-fold less adenoviral DNA than liver of Ad5GFP-treated hCD46Ge mice (Fig. 6). A slight drop in the amount of viral DNA was observed in the spleen of Ad5/35GFP-injected hCD46Ge mice compared with spleen from mice injected with Ad5GFP. The number of viral genomes was elevated ∼2.5-fold in lung, ∼15-fold in heart, ∼2-fold in brain, and ∼4-fold in kidneys of hCD46Ge mice injected with Ad5/35GFP compared with mice injected with Ad5GFP virus. As described previously for the oncolytic viruses, both normal and hCD46Ge mice treated with Ad5GFP had viral genome copy numbers similar to those of normal mice injected with Ad5/35GFP. In contrast, homozygous hCD46Ge mice injected with Ad5/35GFP had significantly lower viral genome copies in liver compared with the liver of normal mice injected with the same virus.
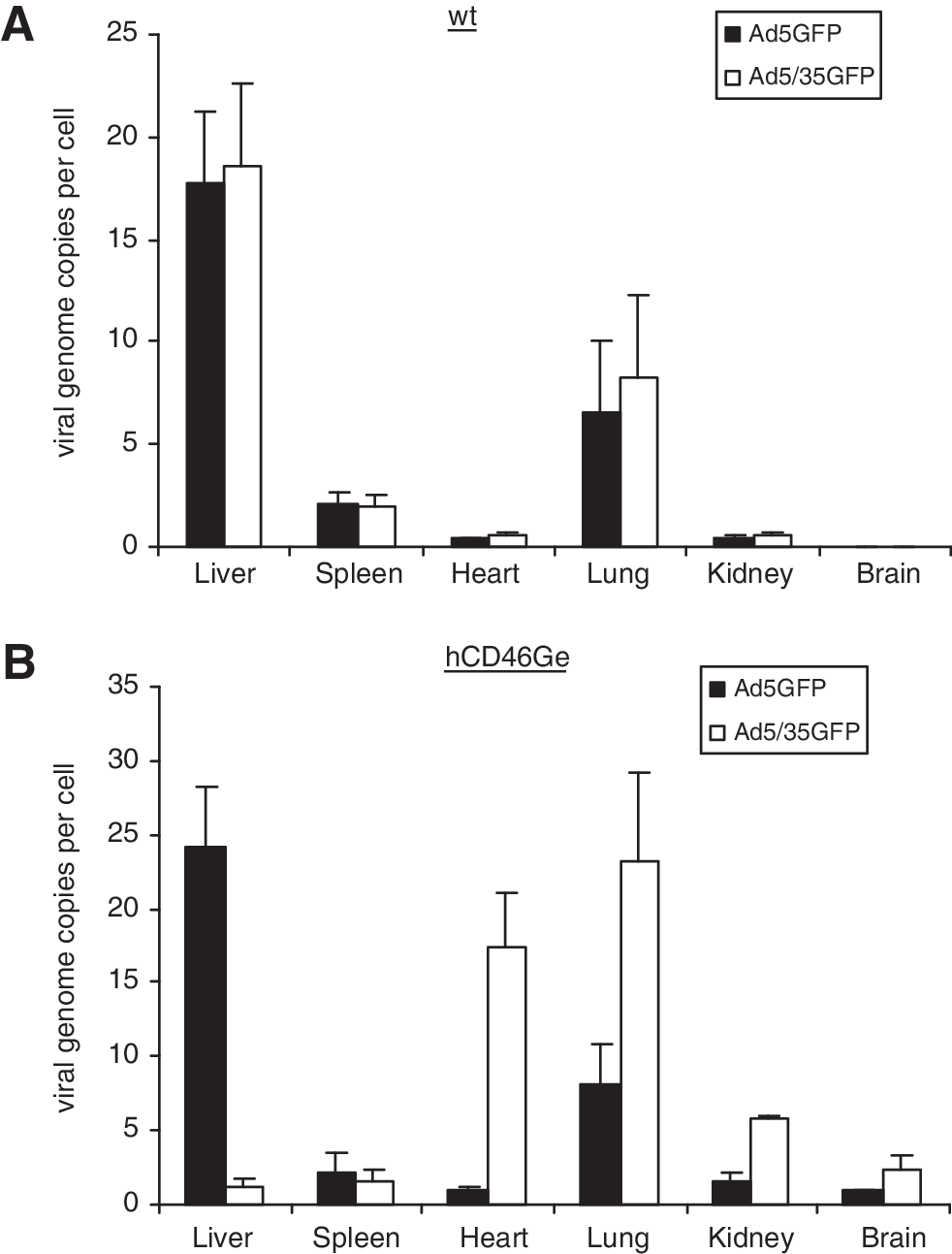
Biodistribution analyses of Ad5- and Ad5/35-based E1-deleted viruses expressing GFP in (
Thus, as was observed in heterozygous hCD46Ge mice, relatively low uptake of chimeric virus in the liver of homozygous hCD46Ge mice correlated with more modest increases in the levels of liver enzymes, IL-6, and TNF-α, confirming that the Ad5/35GFP virus was less hepatotoxic than the Ad5-based Ad5GFP virus in homozygous hCD46Ge mice.
The persistence of each vector in the blood after systemic delivery was determined in homozygous hCD46Ge and normal mice by measuring the adenoviral DNA content in whole blood. The levels of Ad5/35 and Ad5 viral genomes in the blood of homozygous hCD46Ge mice 2 hr after vector delivery were 1.5–2 and 0.1%, respectively, of the theoretical starting level compared with 0.1% of both viruses in normal mice (data not shown), suggesting a slight delay of virus clearance in Ad5/35-transduced hCD46Ge mice compared with normal mice. Overall, the viral genome distribution and inflammatory response were similar in heterozygous and homozygous CD46Ge mice transduced with the fiber-chimeric virus, despite the fact that the latter has 2-fold higher CD46 receptor levels.
In summary, the data demonstrated that after systemic delivery of a single dose of Ad5 or Ad5/35-based virus in hCD46Ge mice, which have a CD46 receptor pattern resembling that of humans, the fiber-chimeric Ad5/35-based virus (Ad5/35GFP or OV-5T35H) was significantly less hepatotoxic than Ad5-based viruses.
Discussion
The therapeutic profile reported to date, using Ad5-based oncolytic viruses as treatment for solid tumors in humans, has been attributed in part to the low and variable expression of CAR, the Ad5 virus receptor, and to the loss of its expression on progression of some tumors (Kim et al., 2002; Mathis et al., 2006). Ad5-based vectors also display acute vector-mediated toxicity associated with complement activation and the release of cytokines and chemokines, and the elevation of liver transaminases (Lieber et al., 1997; Muruve et al., 1999; Muruve, 2004; Shayakhmetov et al., 2005). Adenoviral vectors incorporating fiber proteins from different serotypes have been generated in the hope that transduction efficiency will be increased and toxicities reduced. Several of these modified viruses have shown promise in various applications (Ni et al., 2005).
Viral entry into tumor cells is significantly improved and the antitumor efficacy of chimeric oncolytic viruses is enhanced when the Ad5 fiber knob is replaced with the Ad35 knob, the latter of which recognizes the CD46 receptor that is abundant and widely expressed on tumor cells (Reddy et al., 2006). Biodistribution and safety studies with these fiber-chimeric viruses conducted in normal mice are, however, of limited value because the mouse analog of the CD46 receptor is expressed only in testis, whereas CD46 is expressed in all nucleated cells in humans (Cervoni et al., 1992; Gagger et al., 2003; Segerman et al., 2003). Accordingly, a murine CD46 receptor transgenic mouse strain (hCD46Ge) that expresses the CD46 receptor in a pattern closely mirroring that in humans was chosen for biodistribution studies to evaluate Ad5/Ad35 chimeric viruses in vivo (Mrkic et al., 1998; Kemper et al., 2001). Although the CD46 receptor levels in lymphocytes of the heterozygous mouse strain were slightly lower than in human lymphocytes, the receptor levels on RBCs of heterozygous mice and humans were similar and heterozygous animals were thus used for the primary biodistribution studies. Homozygous mice, which express slightly higher levels of CD46 on RBCs, were used to identify the type of infected cells in the virus-transduced organs, using GFP-expressing, nonreplicating viruses. We have shown previously the superior antitumor efficacy of the Ad5/35 chimeric virus compared with Ad5-based viruses in various xenograft models performed in nude mice (Reddy et al., 2006). Because human tumors do not grow in immune-competent mice, it was unfortunately not possible to repeat these efficacy studies in the immune-competent CD46 receptor transgenic mice used here for the biodistribution studies.
The biodistribution studies showed that the number of Ad5/35 viral genomes in the liver of homozygous and heterozygous hCD46Ge mice on day 3 after virus injection was 20- to 100-old lower than the number of Ad5 viral genomes. Many of the intravenously injected viral particles are taken up by the Kupffer cells, and a dose-dependent increase in hepatocyte transduction occurs only when the Kupffer cells are saturated (Lieber et al., 1997, Tao et al., 2001). This has been shown by temporarily depleting Kupffer cells with reagents such as GdCl2 or clodronate before virus administration, which increased virus transduction of hepatocytes by several fold (data not shown). These data also indirectly show that Kupffer cells take up the majority of intravenously injected virus.
Although it is possible that a portion of the viral genomes detected by qPCR were taken up by Kupffer cells rather than hepatocytes, GFP-positive cells within the liver of homozygous mice were identified by their phenotype as predominantly hepatocytes (Fig. 4A). The lower number of chimeric viral genomes in the liver may have been due to the widespread distribution of CD46 receptors in hCD46Ge transgenic mice and the consequently increased uptake of the virus by the other tissues. It is also possible that the chimeric virus uses a different mechanism independent of the CD46 receptor to enter hepatocytes (Shayakhmetov et al., 2004).
Interestingly, although Ad5wt and OV-5 use the same receptors to enter cells, both transgenic and normal mice injected with Ad5wt consistently had significantly higher levels of vector genomes in the liver compared with OV-5-injected mice. This suggests that the wild-type virus might be replicating in the liver or, alternatively, that DNA replication occurs without forming infectious viral particles, leading to higher numbers of viral genome copies that could ultimately result in higher toxicity, an observation that has previously been seen with Ad5wt in SCID mice (our unpublished data).
Body weight changes seen in mice injected with Ad5wt support this suggestion. Mice that received Ad5wt lost more than 20% of their initial body weight and died within 4 days, compared with about 15% body weight loss and one death among OV-5-treated mice, and no body weight loss in chimeric virus-treated mice or in mice receiving the nonreplicating negative control virus.
The distribution of Ad5wt and OV-5 in hCD46Ge mice was similar to that seen in normal mice. Interestingly, the number of genome copies found in the liver of normal mice after OV-5T35H or OV-5 administration was ∼20- to 100-fold higher than in CD46 transgenic mice (heterozygous or homozygous), suggesting that the Ad5/35-based virus may enter hepatocytes by a receptor-independent mechanism in the absence of CD46 receptors. In contrast, Sakurai and colleagues reported that the transduction level in heart, spleen, lung, and kidney as well as in liver by their Ad35L vector was higher in CD46 receptor transgenic mice (both homozygous and heterozygous) than in normal mice (Sakurai et al., 2006). Although Ad5/35 virus transduction in several organs in the CD46 receptor transgenic mice was higher than in normal mice in our study, it was orders of magnitude lower in the liver of CD46 receptor transgenic mice. This discrepancy seems likely to be due to the significant differences between the viruses. The Ad35L virus used by Sakurai and colleagues consists of the Ad35 capsid, an Ad35 knob, and a fiber with a long shaft whereas the Ad5/35 chimeric virus used in our studies consists of the Ad5 capsid with the Ad5 knob replaced by the Ad35 fiber knob. Although the adenoviral fiber, which is believed to be driving viral entry into cells, is the same in both viruses, virus distribution in the liver of CD46 receptor transgenic mice appears to be significantly different. In any event, these contrasting findings suggest that transduction of liver by the two chimeric viruses may occur in a CD46 receptor-independent manner, or the viral capsid, which differs between the two viruses, is impacting viral entry. It has been shown that the adenovirus serotype 5 hexon is critical for virus infection of hepatocytes (Kalyuzhniy et al., 2008). According to Kalyuzhniy and colleagues, Ad5 hexon binds with high affinity to factor X (FX), leading to efficient Ad5 entry into liver cells in vivo. The authors also showed that both human and mouse FX bind to Ad5 particles as well as to purified Ad5 hexon with identical kinetics and affinity but not to Ad5 knob. This observation correlates with our data demonstrating that Ad5 viral particles and/or Ad5 hexon-carrying vectors seem to transduce liver cells of wild-type mice to the same extent. However, the transduction efficiencies of these vectors in CD46 mice were significantly different irrespective of hexon, suggesting that the CD46 expression in the liver of these mice impacts virus infection.
The virus distribution found in the liver of normal and hCD46Ge (heterozygous) mice correlated with the level of proinflammatory serum cytokines and liver enzymes. Ad5wt, OV-5, and Addl312 vectors resulted in 2- to 3-fold elevated IL-6 serum levels compared with OV-5T35H. Although the IL-6 serum levels observed in mice injected with OV-5, OV-5T35H, or Addl312 decreased at a later point (48 hr), the IL-6 serum levels of Ad5wt-injected mice remained at elevated levels even 48 hr after injection, again suggesting potential virus replication or DNA replication of viral genomes without forming infectious particles in the liver of these animals. hCD46Ge (heterozygous) mice that received Ad5wt showed a significant increase in AST and ALT levels on day 3 after viral infusion. Mice injected with OV-5 also showed increased levels of transaminases. In contrast, the hCD46Ge mice injected with OV-5T35H, Addl312, or PBS had background levels of liver enzymes, suggesting that the chimeric virus may be less hepatotoxic than the OV-5 or Ad5wt virus in CD46 transgenic mice. The normal mice that received either OV-5 or OV-5T35H had similar increases in AST and ALT levels, supporting the observation that these viruses behave differently in normal and hCD46Ge mice. The similar proinflammatory serum cytokine and liver enzyme induction pattern as well as the relatively unaffected whole serum panel chemistries observed in the hCD46Ge (homozygous) mice injected with parental Ad5GFP or chimeric Ad5/35GFP suggest that the fiber-chimeric virus is less toxic in a transgenic mouse strain that expresses the CD46 receptor in a pattern that closely mirrors the expression pattern found in humans.
Although the number of genome copies was lower in the liver of hCD46Ge mice transduced with the fiber-chimeric virus compared to the Ad5 virus (both replicating and nonreplicating viruses), the number of chimeric viral genome copies was higher in other organs. Interestingly, the higher number of chimeric viral genome copies detected in selected organs by qPCR appeared, on the basis of CD105 staining, to be in the endothelial cells (ECs) lining the blood vessels of the organs. CD46 receptors, besides being expressed in endothelial cells lining blood vessels, have also been shown to be expressed in epithelial cells and tissue macrophages in a wide range of peripheral tissues, as well as in Langerhans and squamous cells in the skin in human tissues (Stephen and Louise, 2002). Besides transducing ECs lining the blood vessels of organs, the Ad5/35 chimeric viruses are known to efficiently transduce tumor ECs that also have been shown to overexpress the CD46 receptor (Shinozaki et al., 2006). Targeted destruction of the EC population in tumor vessels is considered an attractive approach for halting tumor growth, as ECs are easily accessible through the bloodstream (Shinozaki et al., 2006). Thus, the Ad5/35 vectors may be a useful agent for targeted antiangiogenic gene therapy to treat cancer if a similar virus distribution pattern occurs in humans. It is worth noting that the low level of the CAR receptor in ECs limits the ability of Ad5-based vectors to target tumor ECs.
In summary, biodistribution studies showed less uptake by the liver of adenoviruses carrying the Ad35 knob than Ad5-based vectors. The lower level of liver transduction observed with the fiber-chimeric viruses correlated with reduced induction of inflammatory responses and elevation of liver transaminases than after treatment with Ad5-based viruses. These findings show that after systemic delivery of a single dose of virus, OV-5T35H may be less hepatotoxic and is better tolerated than the conventional Ad5-based vector, OV-5, in a mouse model that expresses CD46 in a pattern that is similar to that found in humans.
Footnotes
Acknowledgments
The authors thank B. Batiste, S. Ulufatu, J. Ho, and T. Langer (animal facility) for excellent work and Dr. Peter Working for critically reviewing the manuscript. The authors also thank Dr. Roberto Cattaneo (Mayo Clinic) for providing the hCD46Ge mouse strain for breeding.
Author Disclosure Statement
No competing financial interests exist.