
Abstract
A phase 1 clinical trial evaluating the safety of gene therapy for patients with wet age-related macular degeneration (AMD) or retinoblastoma has been completed without problems. The efficacy of gene therapy for Leber's congenital amaurosis (LCA) was reported by three groups. Gene therapy may thus hold promise as a therapeutic method for the treatment of intractable ocular diseases. However, it will first be important to precisely evaluate the efficiency and safety of alternative gene transfer vectors in a preclinical study using large animals. In the present study, we evaluated the acute local (ophthalmic) and systemic toxicity of our simian immunodeficiency virus from African green monkeys (SIVagm)-based lentiviral vectors carrying human pigment epithelium-derived factor (SIV-hPEDF) for transferring genes into nonhuman primate retinas. Transient inflammation and elevation of intraocular pressure were observed in some animals, but these effects were not dose dependent. Electroretinograms (ERGs), including multifocal ERGs, revealed no remarkable change in retinal function. Histopathologically, SIV-hPEDF administration resulted in a certain degree of inflammatory reaction and no apparent structural destruction in retinal tissue. Regarding systemic toxicity, none of the animals died, and none showed any serious side effects during the experimental course. No vector leakage was detected in serum or urine samples. We thus propose that SIVagm-mediated stable gene transfer might be useful and safe for ocular gene transfer in a clinical setting.
Introduction
On the other hand, adverse events have been reported in some gene therapy clinical trials (Marshall, 1999; European Society of Gene Therapy, 2003; Woods et al., 2006; Cavazzana-Calvo and Fischer, 2007). Four of eight patients in a phase 1 trial in France to treat patients with X-linked severe combined immunodeficiency disease (SCID-X1) have developed T cell lymphoma, and the same adverse event was observed in patients in a phase 1 trial for SCID-X1 in the United Kingdom. To prevent adverse events in clinical gene therapy trials, it is important to precisely evaluate the potential safety issues surrounding the gene transfer vectors themselves. Moreover, because the eye is one of the sensory organs, a careful assessment of local toxicity, including potential effects on retinal function, is required. Numerous vectors for ocular gene transfer and gene therapy, including an adenoviral vector (Bennett et al., 1994; Sakamoto et al., 1998), an AAV vector (Ali et al., 1996, 2000; Flannery et al., 1997), a lentiviral vector (Miyoshi et al., 1997; Duisit et al., 2002; Ikeda et al., 2003; Miyazaki et al., 2003), and a Sendai virus vector (Ikeda et al., 2002; Murakami et al., 2008a) have been tested in rodents. However, the anatomical features of the primate retina are different from those of the rodent retina, and thus the retinal gene transfer characteristics may also differ between primates and rodents. To achieve successful translation to a clinical trial, it is important to precisely evaluate the efficiency and potential safety issues using nonhuman primates.
Previously, we demonstrated efficient and stable retinal gene transfer mediated by a nonpathogenic simian immunodeficiency virus from African green monkey (SIVagm)-based lentiviral vector in rodent retinas, and the therapeutic outcome in an animal model of retinal degeneration, using recombinant SIVagm-based lentiviral vectors carrying human pigment epithelium-derived factor (hPEDF) (Ikeda et al., 2003; Miyazaki et al., 2003, 2008; Murakami et al., 2008b). Transgene expression for more than 1 year was seen only in the retinal pigment epithelium via a subretinal injection route (Ikeda et al., 2003), and neuroprotective gene therapy using hPEDF conferred protection against the retinal degeneration and functional defects in Royal College of Surgeons (RCS) rats and retinal degeneration slow (rds) mice (Miyazaki et al., 2003, 2008; Murakami et al., 2008b). Further evaluation in large animals, such as nonhuman primates, must be performed before a clinical application. In the case of lentiviral vectors, there has been only one report demonstrating efficient gene transfer to nonhuman primate retinas (Lotery et al., 2002), despite the many reports of efficient gene transfer using recombinant adeno-associated virus (rAAV) vectors (Bennett et al., 1999; Lotery et al., 2003; Le Meur et al., 2005).
In the present, preclinical toxicity study, we demonstrate the acute local (ophthalmic) and systemic safety of intraocular administration of SIV-hPEDF in Macaca fascicularis. Our results showed that the toxicity of SIV-hPEDF was tolerable at a titer at least 40-fold higher than the clinically available titer. We are currently conducting a long-term safety study including the possibility of carcinogenesis in nonhuman primates.
Materials and Methods
Simian immunodeficiency virus from African green monkey (SIVagm)-based lentiviral vectors
Third-generation recombinant SIVagm-based lentiviral vectors carrying the human pigment epithelium-derived factor (hPEDF) were prepared as previously described (Miyazaki et al., 2003, 2008). Briefly, human embryonic kidney (HEK)-293T cells were transfected with the packaging vector, the gene transfer vectors encoding hPEDF driven by the cytomegalovirus (CMV) promoter, the Rev expression vector, and the envelope vector pVSVG (Clontech Laboratories, Mountain View, CA). A U3 region in the 3′ and 5′ long terminal repeat (3′ and 5′ LTR) of SIVagm was deleted to induce self-inactivation. The viral titer was determined by transduction of the HEK-293T cell line and expressed as transducing units per milliliter (TU/ml), and these viruses were kept at −80°C until just before use. Vector stocks were confirmed to be free from endotoxin, and without extraordinary cytotoxicity by simultaneous transfection testing using HEK-293T cells and human retinal pigment epithelium (RPE) cells (ARPE-19) obtained from the American Type Culture Collection (Manassas, VA).
Retinal gene transfer in nonhuman primates
All nonhuman primates were cared for in accordance with the Association for Research in Vision and Ophthalmology (ARVO, Rockville, MD) guidelines for the use of animals in ophthalmic and vision research. This study was conducted according to the Rules for Animal Care and Management of the Tsukuba Primate Research Center (Honjo, 1985) and the Guiding Principles for Animal Experiments Using Nonhuman Primates, formulated by the Primate Society of Japan (1986). Protocols of experimental procedures were approved by the Animal Welfare and Animal Care Committee of the National Institute of Biomedical Innovation (Osaka, Japan).
Anesthetization was achieved with ketamine (6 mg/kg) and xylazine (1.2 mg/kg). For all subretinal injections, we used an operating microscope to monitor related events. First, approximately 20 μl of balanced salt solution (BSS: 122.2 mM NaCl, 5.08 mM KCl, 1.05 mM CaCl2, 0.98 mM MgCl2, 25.0 mM NaHCO3, 3.0 mM Na2HPO4, 5.11 mM dextrose; pH 7.4) was injected into the subretinal space through the pars plana via a glass capillary tube. The anterior chamber was then tapped with 26-gauge needles to remove the aqueous humor. Finally, approximately 20–50 μl of SIV-hPEDF solution (low titer, 2.5 × 107 TU/ml; high titer, 2.5 × 108 TU/ml; or maximal titer, 1.0 × 109 TU/ml) or BSS was injected into the subretinal space from the previous retinal hole. The total solution volume injected into the subretinal spaces was approximately 40–70 μl.
Acute toxicity study protocol
Eleven Macaca fascicularis (body weight, 2.47–6.15 kg) were enrolled in this toxicity study (see Supplementary Table 1 at

Time schedule of the acute toxicity study.
Color fundus photography and fluorescein angiography
The animals were anesthetized and their pupils were dilated with 2.5% phenylephrine (Santen, Osaka, Japan), and the animals were placed on a bed. A portable fundus camera was used to take color fundus photographs (RC-II; Kowa, Tokyo, Japan), and a portable fundus camera (RC-II) with interference filters (Z6, Z7; Spectrotech, Lincoln, MA) was used for the fluorescein angiography.
Electroretinography
Full-field ERGs (fERGs) and multifocal ERGs (mfERGs) were recorded before and 90 days after the treatments. Each monkey was anesthetized with ketamine (6.0 mg/ml) and xylazine (1.2 mg/ml) and then kept in a dark room for at least 30 min under a dim red light. After this acclimatization period, the pupils were dilated with drops of 2.5% phenylephrine HCl and 1.5% proparacaine HCl and the monkeys were placed on a flat pad throughout the experiment.
fERGs of each eye were recorded with a commercial contact lens electrode with a white/color LED stimulator (LS-W; Mayo, Aichi, Japan), either in the dark or against a white adapting field (25 cd/m2). The corneal electrode was placed on the cornea after the corneal surface had been treated with 1% methylcellulose for protection. A reference needle electrode was placed into the forehead muscle and a ground needle electrode was also inserted into the femoral muscle. The impedances between the corneal and reference electrodes were less than 5 kΩ. Initially, the responses to four successive flashes (0.009 cd·sec/m2) at an interstimulus interval of 2 sec were averaged for the rod response. The responses for the combined ERG and the oscillatory responses were then recorded at flash intensities of 3 and 30 cd·sec/m2. After the recording of dark-adapted (DA) ERGs, the animals were exposed to a white adapting field for at least 10 min and the light-adapted (LA) ERGs elicited by a flash of 3 cd·sec/m2 were recorded. Responses to 16 successive flashes presented at a rate of 1 Hz were averaged for single-flash cone ERG, and 15 successive flashes presented at a rate of 30 Hz were averaged for flicker ERG. In addition, on–off responses to 16 successive flashes (45 cd·sec/m2) were recorded against a white adapting field (40 cd/m2). Responses were differentially amplified and filtered between 0.3 and 200 Hz.
mfERGs of each eye were separately recorded with a bipolar contact lens electrode of the Burian-Allen type (Hansen Ophthalmic Laboratories, Iowa City, IA) or Mayo type (Mayo). The corneal electrode was placed on the cornea after the corneal surface had been treated with 1% methylcellulose for protection. A ground needle electrode was inserted into the left arm muscle. The electrode impedance was maintained at less than 5 kΩ. The mfERG signals were bandpass filtered between 3 and 300 Hz in conjunction with 60-Hz notch filtering. Stimuli were generated on a 21-in. monitor (Radius Intelicolor; Radius, San Jose, CA) controlled by VERIS 1 software and a video driver board (Electro Diagnostic Imaging, San Mateo, CA). A CRT frame frequency of 67 Hz was used. An array of 19 or 37 hexagonal elements of equal size was used for this mfERG recording. The monitor screen subtended approximately 40° × 40° at a viewing distance of 16.5 cm. The mean luminance of dark stimulus elements was 5 cd/m2 and the mean luminance of light stimulus elements was 95 cd/m2 according to a binary m-sequence. The examined eyes of anesthetized animals were fixated on the center of the screen during the recording (2 min).
Immunohistochemistry
Immunohistochemical analysis was performed to evaluate the therapeutic transgene expression. The eyes were enucleated and fixed with ice-cooled 4% paraformaldehyde in 0.1 M phosphate-buffered saline (PBS) for 1 day at 4°C. They were then mounted in paraffin, and 5-μm-thick sections were made and allowed to dry overnight. The specimens were incubated overnight at 4°C with mouse monoclonal anti-human PEDF IgG antibody (Chemicon International, Temecula, CA) diluted 1:400. Signals were then developed with EnVision/HRP (Dako, Carpinteria, CA) and a Vector VIP peroxidase substrate kit (Vector Laboratories, Burlingame, CA).
Reverse transcription-polymerase chain reaction assay
To detect the vector sequence in serum or urine samples, we performed a reverse transcription-polymerase chain reaction (RT-PCR) assay. Sixty-microliter RNA solutions were extracted from 140-μl serum or urine samples, using a QIAamp viral RNA mini kit (Qiagen, Valencia, CA) according to the manufacturer's protocol. The SIVagm-based lentiviral vector sequence was amplified with 22 μl of the 60-μl RNA solutions by RT-PCR, using a One-Step RT-PCR system (Invitrogen, Carlsbad, CA) according to the manufacturer's protocol. As a positive control, SIV-hPEDF vector solutions were step-diluted (100 to 3 viral genomes [VG] per 140 μl of vector solution) and amplified. The minimal sensitivity of this analysis was 6 VG per 140 μl of vector solution. The primers used for the SIV vector sequences were 5′-CGGAGGGCTTAAAAAGTCTGTTC-3′ and 5′-ATAGGGCTTGAAACATGGGTACT-3′, with an expected PCR product of 330 bp.
PCR assay
PCR analysis for the presence of integrated SIV-based lentiviral vector (SIVagm proviral genome) sequences was performed. Genomic DNAs from whole blood samples were extracted with a GenElute mammalian genomic DNA kit (Sigma, St. Louis, MO). The SIVagm-based lentiviral vector sequence was amplified with 500 ng of genomic DNA by PCR. The same primers as described previously were used for the SIV vector sequences. As a positive control, gene transfer vector plasmids for SIV-hPEDF vector were step-diluted (100 to 6 copies of plasmid per tube). The minimal sensitivity of this analysis was 12 plasmids per tube. Monkey β-actin was simultaneously amplified with 15 ng of genomic DNA. The primers used for monkey β-actin were 5′-TGACGGGGTCACCCACACTGTGCCCATCTA-3′ and 5′-CTAGAAGCATTTGCGGTGGACGATGGAGGG-3′, with an expected PCR product of 661 bp.
Results
Evaluation of ophthalmic toxicity
Slit lamp biomicroscopy
Slit lamp biomicroscopy was performed to assess the status of the wound and cornea, conjunctival erythema, anterior chamber (AC) cells, AC flare, lens opacity, and anterior vitreous cells. The clinical findings of all animals until 30 days after gene transfer are listed in Table 1. Corneal opacity (edema) and anterior segment inflammation were observed in some animals in the early phase (day 1–day 8); however, these effects had almost entirely disappeared by 30 days after treatment. Only one eye (the right eye of animal 10, treated with the maximal titer) showed an inflammatory reaction in the anterior segment (keratic precipitate and anterior vitreous cells) on day 30, and there was no apparent inflammatory response in the retina and choroid by funduscopic examination. These findings were diminished by the use of topical steroid. There were no apparent clinical findings in any of the animals on day 90.

Abbreviations: a.c., anterior chamber; a.v., anterior vitreous; BSS, balanced salt solution; dis, discharge; hyper, hyperemia; KP, keratic precipitate; L, left; max, maximum; opa, opacity; pig, pigment; R, right; SPK, superficial punctate keratopathy.
Intraocular pressure measurement
We performed IOP measurement by Tono-Pen tonometry at various time points after gene transfer (days 1, 8, 30, and 90). As shown in Fig. 2, two eyes (the right eye of animal 14, treated with a high titer and the left eye of animal 15, treated with the maximal titer) showed a mild IOP elevation (animal 14, right eye: 23 mmHg; animal 15, left eye: 27.3 mmHg). There were no apparent clinical findings, including AC inflammation, by slit lamp biomicroscopic assessment. Moreover, there was no apparent histopathological change in the iridocorneal angle (see Supplementary Fig. 1 at
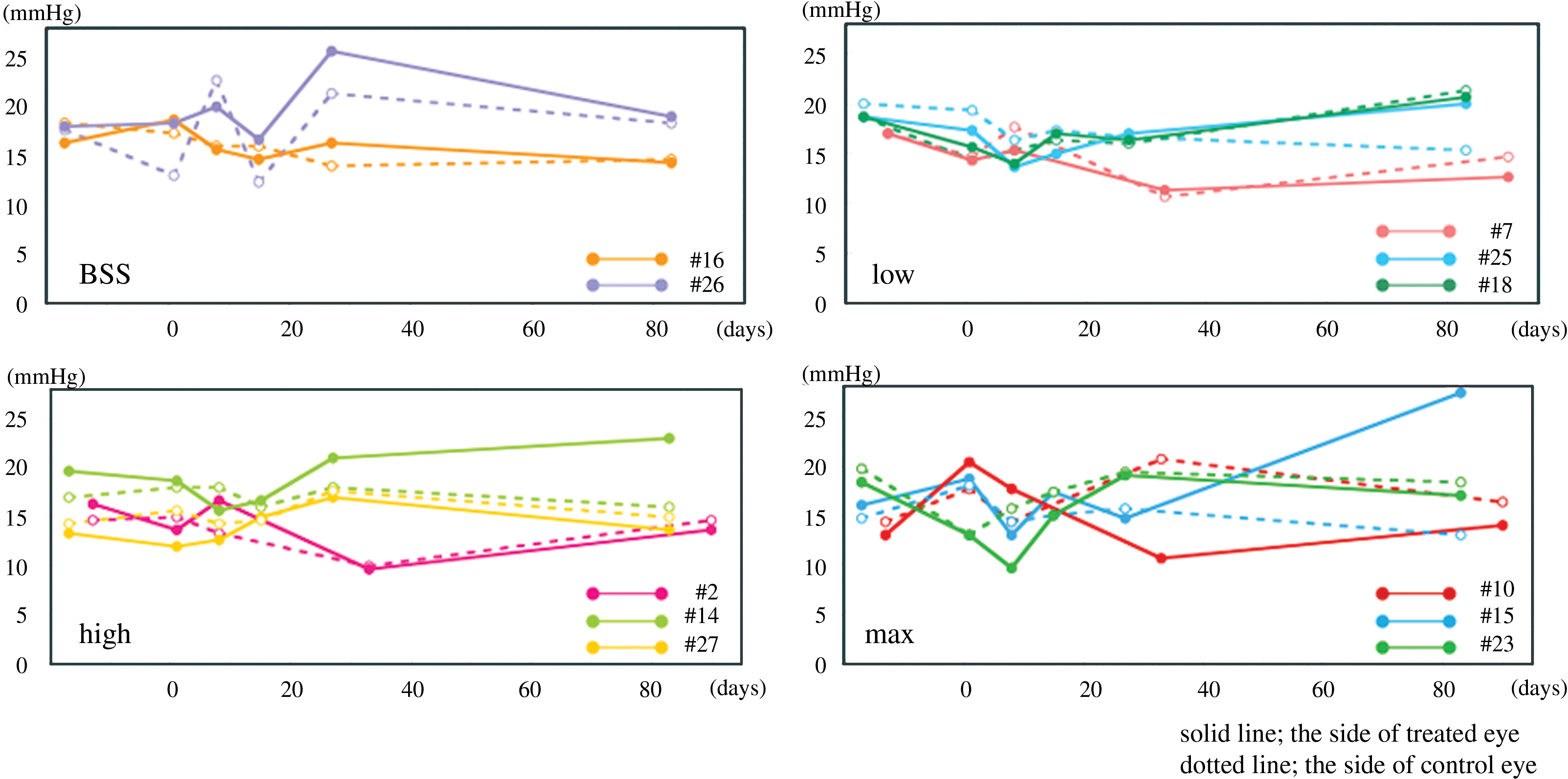
Intraocular pressure (IOP) history over the course of study for all animal eyes. On day 90, two eyes showed slight to mild IOP. The solid line and solid circles indicate treated eyes, and the dotted lines and open circles indicate control eyes. BSS, balanced salt solution. Color images available online at
Indirect ophthalmoscopy
Indirect ophthalmoscopy was performed to assess the status of the optic disk, retina, and choroid. Examination by indirect ophthalmoscopy at all time points revealed an absence of retinal detachment and an absence of apparent inflammatory reaction of retina and choroid. The optic disk color was good and there was no apparent disk edema. At the area of vector injection the retina showed hyper- or depigmentation. A focal retinal pigment epithelium (RPE) defect and retinal degeneration caused by needle damage were seen in all treated eyes (Fig. 3; and see Supplementary Fig. 2 at
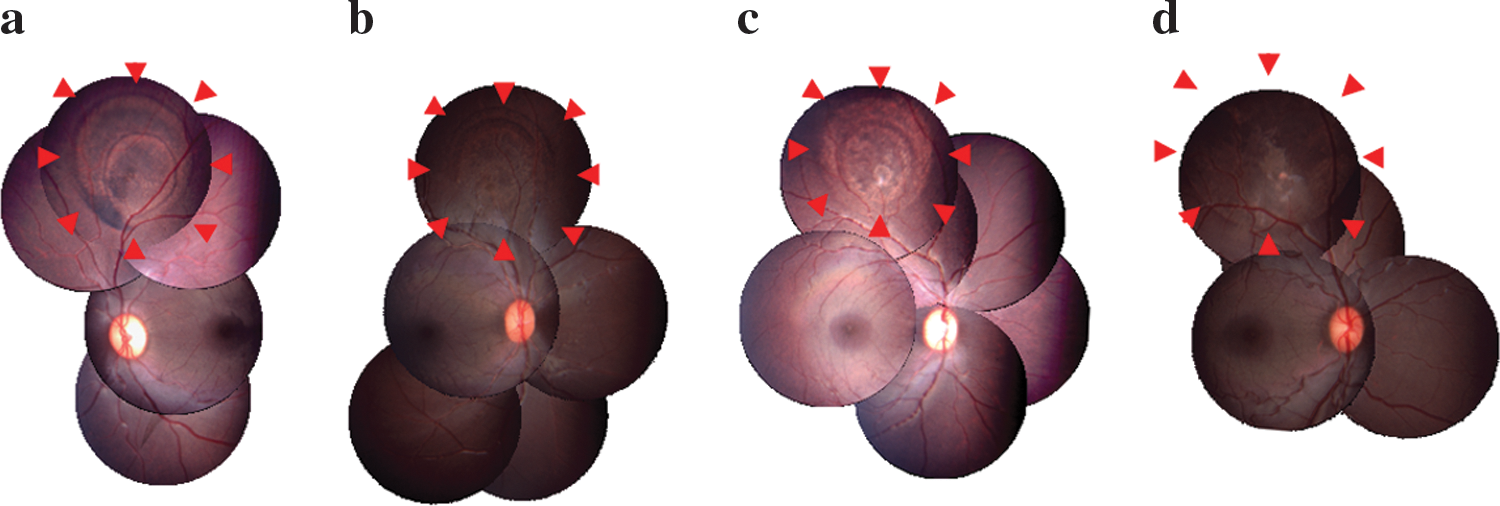
Color fundus photography of (
Fluorescein angiography
Fluorescein angiography showed a window defect of the treatment lesion in all treated eyes (Fig. 4a and c). Except for one eye treated with the maximal titer, neither late fluorescein leakage nor staining was seen on day 90 (Fig. 4b). Only one eye treated with the maximal titer (animal 23, right eye) showed focal late fluorescein staining, caused by the focal subretinal granulation tissue due to retinal paracentesis (Fig. 4d).

Fluorescein angiograms of eyes injected with a high titer (animal 2, right eye;
Electroretinograms
The full-field ERGs (fERGs: rod and cone responses) on day 90 from untreated controls, BSS-treated eyes, and SIV-hPEDF-treated eyes (low titer, high titer, and maximal titer) are illustrated in Fig. 5. A summary of the fERG responses (amplitude and implication time) is shown in Supplementary Table 2 (see at
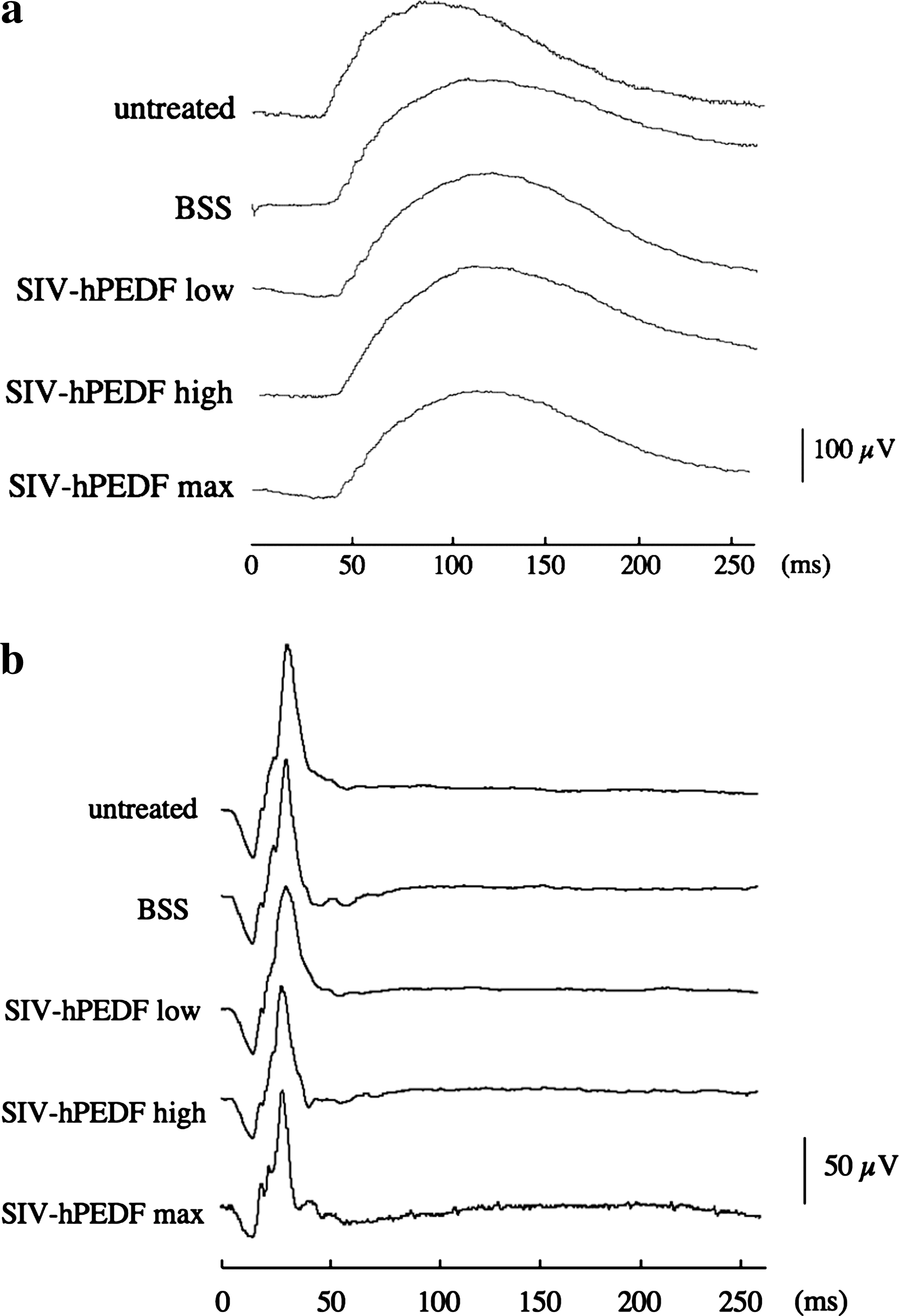
Full-field ERG (fERG) responses obtained from untreated control eyes, BSS-treated eyes, and SIV-hPEDF-treated eyes on day 90. (
Each data of an array of 19 or 37 hexagonal elements of the multifocal ERGs (mfERGs) in untreated controls, BSS-treated eyes, and SIV-hPEDF-treated eyes were analyzed separately. For each eye, the mfERG data of the vector- or BSS-injected area were compared with the data for the uninjected area of the opposite hemisphere of the same eye. As shown in Supplementary Tables 3 and 4 (see at

Multifocal ERG (mfERG) response (37 hexagonal elements) and injected and uninjected area responses obtained from untreated control eyes, BSS-treated eyes (
Histopathological findings of retina
All retinas treated with either SIV-hPEDF or BSS showed a certain degree of inflammatory infiltration. Except for the retinal paracentesis site for vector injection, no retinal structural destruction, such as was previously reported after administration of a high titer of SIVagm-based lentiviral vector to rodent retinas (Ikeda et al., 2003), was apparent even in eyes treated with the maximal titer of SIV-hPEDF (Fig. 7a and b), or in retinas treated with a low or high titer of SIV-hPEDF or with BSS (data not shown). The RPE appeared to be repopulated with variable pigmentation and microvilli, and with pigment-laden cells that overlaid the renewed RPE. Around the retinal paracentesis site, focal retina tissue showed gliosis and formation of subretinal granulation tissue, suggesting operation-related change (data not shown). As shown in Fig. 7c and d, an abundant amount of secreted hPEDF protein (purple staining) was accumulated in the rods and cones layer, just as in human retinal tissue. There were no apparent changes in the optic nerve, retina, or RPE outside of the vector-injected area, including macular changes.
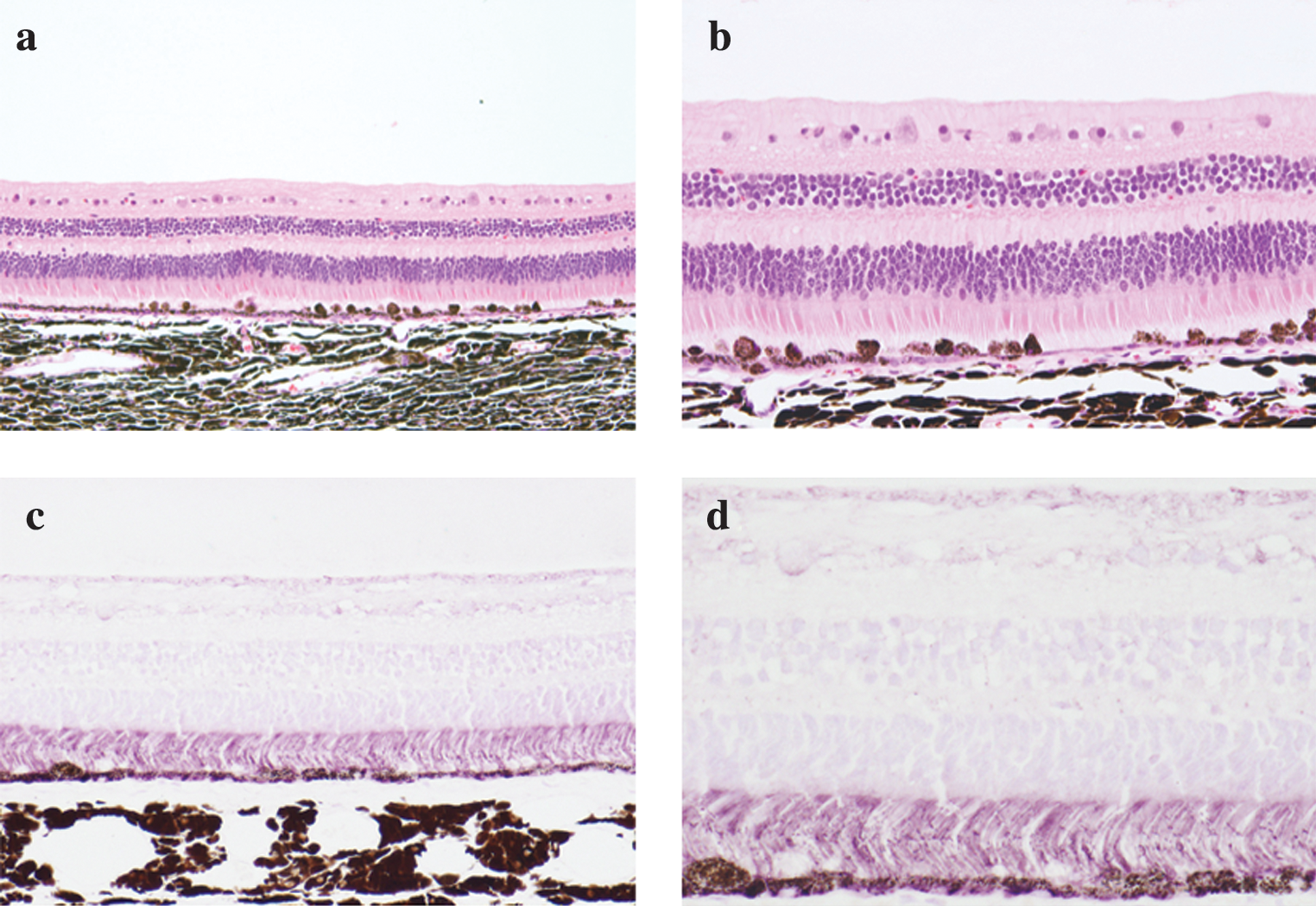
(
Subretinal injection of SIV-hPEDF results in a well-tolerated level of systemic toxicity in the acute phase
General body conditions
None of the animals died or experienced serious side effects during the experimental course. General conditions including activity, appetite, and body weight showed no apparent changes in any of the animals. Some animals (2 and 26) suffered from diarrhea for a short period of time.
Vital signs
Body temperature changed in all treated animals within 14 days of treatment, suggesting an operation-related event. There was no apparent change in heart rate or either systolic or diastolic blood pressure in any of the animals during the experimental course.
Hematology
A mild and transient increase in the number of white blood cells was observed on days 1 and 8 in 8 of 11 animals, including both BSS- and vector-treated ones (see Supplementary Table 5 at
Blood chemistry
(See Supplementary Table 6 at
Urine chemistry
There were no changes in any of the parameters measured in urine throughout the course of the experiment.
Vector sequence in serum and urine samples
To detect the vector sequence in serum or urine samples, we performed an RT-PCR assay. At the level of sensitivity of our assay (the minimal sensitivity of this analysis was 6 VG/140-μl sample; Fig. 8), no vector sequence was detected at any of the measurement time points.

RT-PCR assay for the SIV-hPEDF vector sequence. We performed an RT-PCR assay to detect the vector sequence in serum or urine samples at various time points (preoperation and on days 1, 8, 30, and 90). No vector sequence was detected at the level of sensitivity of this analysis (the minimal sensitivity of this analysis was 6 VG/140-μl sample).
Integrated vector sequence in whole blood samples
PCR analysis for the presence of integrated SIVagm-based lentiviral vector (SIVagm proviral genome) sequences was performed. At the level of sensitivity of our assay (the minimal sensitivity of this analysis was 12 plasmids per 500 ng of genome DNA; Fig. 9), no integrated proviral sequence was detected at any of the measurement time points.

PCR assay for the SIVagm proviral genome. We performed a PCR assay to detect the presence of an integrated SIVagm-based lentiviral vector sequence in whole blood samples at various time points (preoperation and on days 1, 8, 30, and 90). No integrated proviral sequence was detected at the level of sensitivity of this analysis (the minimal sensitivity of this analysis was 12 plasmids per 500 ng of genome DNA).
Results of autopsy
Gross and microscopic examination showed no apparent serious change, including carcinogenesis, in major organs (brain, heart, lung, liver, spleen, pancreas, kidney, adrenal gland, intestine, and reproductive organ). Lymphoid follicles were observed in spleen and/or lymphoid tissue of nine animals, including a BSS-treated one. No vector-related event was detected.
Discussion
In this work, we assessed acute local (ophthalmic) and systemic toxicity after intraocular administration of SIV-hPEDF in Macaca fascicularis as a preclinical toxicity study. The key observations obtained in this study were that (1) SIVagm-mediated gene transfer neither affected retinal function nor damaged retinal architecture; (2) in the systemic evaluation, a mild and transient inflammatory reaction was seen in all animals, and was suggested to be an operation-related event; and (3) at all time points, neither vector particles nor integrated proviral vector sequences were detected in serum or urine samples. The current study indicated that these toxicities were tolerable at an at least 40-times higher titer than that clinically available.
Regarding acute toxicity at the anterior segment of eyes in this study, ocular inflammation was milder than in the previous safety study using an rAAV vector (rAAV2.RPE65), which resulted in frequent ocular inflammation (75%) (Narfström et al., 2003). Only one eye (the right eye of animal 10, treated with the maximal titer) showed an inflammatory reaction (keratic precipitate and anterior vitreous cells) on day 30. The inflammatory response was well controlled by topical steroid, but the reason for the late-phase inflammatory reaction was unclear. We checked serum immunoglobulin G against the vector envelope protein (VSV-G protein) and human PEDF protein 30 days after vector administration. We could not detect the specific immune reaction using a dot-blot assay (data not shown). One study using an rAAV vector reported a delayed-onset intraocular inflammatory response either to the presence of a contaminant in the vector preparation or to an immune reaction to the expressed foreign protein (Bainbridge et al., 2003). In our clinical trial, it will be crucial to use good manufacturing practice (GMP)-grade vectors. In addition, we will need to administer local or systemic steroid to reduce the possibility of intraocular inflammation.
Next, we observed a mild IOP increase in two eyes (the right eye of animal 14, treated with a high titer; and the left eye of animal 15, treated with the maximal titer). The exact reason for this finding was unclear, because there was no AC inflammation or other clinical findings by slit lamp biomicroscopic assessment and no apparent histopathological change at the iridocorneal angle (Supplementary Fig. 1). Thus, we should carefully check up on IOP change in our long-term safety study using nonhuman primates; there was no evidence of aberrant IOP elevation through a 3-year observation period (data not shown).
We previously observed retinal damage at about day 90 after administration of a high titer of SIV-nls-lacZ and SIV-EGFP (enhanced green fluorescent protein) to the eyes of rats (Ikeda et al., 2003), and similar findings of a reduction in reporter gene expression have been observed in ocular tissue and other organs (van Ginkel et al., 1997; Reichel et al., 1998; Stripecke et al., 1999). In all of these studies, a clear association was observed between the decline in reporter gene expression and the host immune response. Reports have also suggested that the gradual reduction in GFP expression might be due to a host immune response against GFP itself when using the lentiviral vector (Doi et al., 2002, 2004). Doi and colleagues demonstrated that no retinal damage was seen in eyes treated with an empty HIV-based lentiviral vector, and transient immunosuppression resulted in prolongation of the transgene expression. We therefore attempted further evaluation using an empty vector without transgene (empty-SIV) in rodent retinas. Most of the empty-SIV-treated retinas showed no degeneration and no damage to retinal function on day 90 (data not shown). Moreover, the most critical distinction between the previous study and this study concerns vector generation. The SIV vectors used in the previous study were older and immature second-generation vectors, whereas our more recent studies using third-generation vectors revealed no apparent retinal damage (Miyazaki et al., 2003, 2008; Murakami et al., 2008b). In this preclinical toxicity study using large animals, there was neither histopathological destruction nor functional failure in any of the treated retinas (Figs. 5 –7). Only one eye treated with the maximal titer (animal 23, right eye) showed local late fluorescein staining on day 90 (Fig. 4d). Histopathologically, focal subretinal granulation tissue only around the retinal paracentesis site was observed, suggesting operation-related change. However, we should carefully check up on retinal function in our long-term safety study using nonhuman primates.
Regarding acute systemic toxicity, a mild and transient inflammatory reaction was observed in the hematology and blood chemistry data. These findings were seen in both BSS- and vector-treated animals, and thus were suggested to be related to the operation, not to vector-related events. However, it is important to carefully monitor acute systemic immune reactions, such as the systemic inflammatory response syndrome, or so-called SIRS, which is recognized as an important adverse effect of adenoviral gene therapy (Marshall, 1999). A previous study has reported systemic vector dissemination after subretinal injection of an rAAV vector (Weber et al., 2003). Systemic vector leakage is one of the most important problems in gene therapy, and causes germ line transmission or carcinogenesis in other organs. In this study, we could detect neither vector particles nor integrated proviral vector sequences in serum, urine, or whole blood samples at the level of sensitivity employed (Figs. 8 and 9). A previous report showed that VSV-G-pseudotyped lentiviral vectors were rapidly inactivated by human serum (Higashikawa and Chang, 2001). If vector leakage into the circulation occurred in the case of our vectors, it would be difficult for the vectors to disseminate to other organs, even by germ line transmission. However, we should carefully check up on the vector dissemination, including a check for carcinogenesis, in our ongoing long-term safety study using nonhuman primates.
In summary, the results of our current toxicity study using nonhuman primates suggested that the local (ophthalmic) and systemic toxicity of the SIVagm-based lentiviral vector-mediated subretinal gene delivery was well tolerated. To successfully translate this gene delivery system to clinical gene therapy for retinitis pigmentosa, we are currently conducting a preclinical safety study, which will estimate the local and systemic toxicity and the carcinogenesis over the long term, using nonhuman primates receiving subretinal injection of our third-generation SIV-hPEDF.
Footnotes
Acknowledgments
The authors thank Y. Katakai, E. Nagasaka, M. Yoshikawa, and H. Fujii for assistance with the experiments. KN International provided language assistance. This work was supported in part by a Grant-in-Aid (to Y.I., Y.Y., K.S., and T.I.) from the Japanese Ministry of Education, Culture, Sports, Science, and Technology (15209057, 16390118, 17689047, and 19209012) and by a grant for the Promotion of Basic Science Research in Medical Frontiers from the National Institute of Biomedical Innovation (Y.Y., K.S., and T.I., project 21).
Author Disclosure Statement
Yoshikazu Yonemitsu is a member of the Scientific Advisory Board of DNAVEC.