
Abstract
Recombinant adeno-associated virus (rAAV) vectors represent a promising approach to gene delivery for clinical use. Published data indicate that rAAV vector genomes persist in vivo as episomal chromatin in the skeletal muscle of nonhuman primates. In this study, we assessed the interconnection between the transcription factor cyclic AMP response element-binding protein (CREB) and recombinant AAV serotype 2 vector genomes after transduction in vitro and in vivo. rAAV-mediated myocyte transduction was potently blocked in the hearts of mice expressing CREB-S133A, which is a CREB-S133A dominant-negative mutant. Isoproterenol, a strong CREB activator, prominently increased rAAV transduction and the increase was abrogated by silencing the CREB gene with small interfering RNA. In addition, rAAV infection of muscle cells mildly but significantly induced CREB protein phosphorylation at serine-133, and was capable of stimulating CREB-dependent transcription from a reporter plasmid. Using chromatin immunoprecipitation and immunoblotting assays, both CREB and p300 were found to physically associate with two different rAAV genomes. Accordingly, CREB/p300 appears to have a role in rAAV transduction to establish active vector transcription in heart muscle cells.
Introduction
Here we examine the relationship between rAAV transduction and CREB and show that rAAV transduction was strongly suppressed in the heart of CREB-S133A transgenic mice, which overexpress a dominant-negative mutant CREB protein. Both CREB and p300 physically associate with the rAAV genome, as shown in a chromatin immunoprecipitation (ChIP) assay. Isoproterenol, which is a strong CREB activator, prominently increased rAAV transduction and the increase was abrogated by silencing the CREB gene with small interfering RNA (siRNA).
Materials and Methods
rAAV vector production
Vectors rAAV-CMV-hGH/lacZ and rAAV-MHC-hGH/lacZ, with titers of 2 × 1011 to 1 × 1012 vector genomes (VG)/ml, were prepared as described previously (Aikawa et al., 2002).
Protein analysis
Protein was separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and blotted onto nitrocellulose membrane. Anti-CREB antibody (cat. no. sc-58; Santa Cruz Biotechnology, Santa Cruz, CA), anti-phospho-CREB antibody (cat no. 9190; Cell Signaling Technology/New England BioLabs, Danvers, MA), and anti-p300 antibody (cat no. sc-585; Santa Cruz Biotechnology) antibodies were used for Western blot analysis using an ECL Advance chemiluminescence system (RPN2135; GE Healthcare Life Sciences, Piscataway, NJ). Coimmunoprecipitation of p300 and CREB was performed 24 hr after rAAV infection by incubating cell lysates with anti-p300 antibody followed by precipitation with protein A beads (P1052; Sigma-Aldrich, St. Louis, MO), and the precipitate was analyzed with anti-CREB antibody.
Transfection
The CRE-luc reporter plasmid was constructed by cloning three copies of a somatostatin gene CRE element in front of the thymidine kinase promoter in the pGL2-basic plasmid (courtesy of J. Lepore, University of Pennsylvania, Philadelphia, PA). Transient transfection assays were performed in 12-well plates seeded at 2–3 × 105 cells per well with C2C12 (mouse skeletal myoblasts) and H9C2 (rat cardiomyocytes), using PolyFect reagent (cat. no. 301107; Qiagen, Hilden, Germany) with 50 ng of reporter plasmid. rAAV or 10 μM forskolin (Sigma-Aldrich) was added 5 hr after transfection. After 24 hr, the C2C12 and H9C2 cells were harvested and luciferase activity (luciferase assay system; Promega, Madison, WI) was measured with a luminometer (Luminoskan Ascent; Thermo Fisher Scientific, Waltham, MA). To control for transfection efficiency luminescence values were normalized to the total protein concentration measured by the bicinchoninic acid (BCA) protein assay (cat no. 23223; Pierce Biotechnology/Thermo Fisher Scientific, Rockford, IL).
Cell culture and rAAV transduction assays
Twelve-well plates were seeded at 2–3 × 105 per well with H9C2 cells and C2C12 cells, and the cells were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% fetal bovine serum and penicillin (100 units/ml) in a humidified atmosphere at 37°C with 5% CO2. When cells were 70–80% confluent, the medium was replaced with serum-free fresh medium, and then rAAV-CMV-lacZ (2000 VG/cell) was added. After a 24-hr incubation, the cells were fixed and stained for β-galactosidase (β-Gal) activity (Aikawa et al., 2002). For the siRNA experiments, both mouse CREB-1 siRNA (cat. no. sc-35111; Santa Cruz Biotechnology) and control siRNA (cat. no. sc-36869; Santa Cruz Biotechnology) were purchased, and rAAV-CMV-lacZ was infected 24 hr after transfection.
Animal studies
MHC-CREB-S133A mice (CREB mice) are courtesy of J. Leiden (Fentzke et al., 1998). The Tufts University Institutional Animal Care and Use Committee approved all animal studies. Anesthetized 4- to 6-week-old wild-type or CREB mice received 50 μl containing 1 × 1010 VG of rAAV-hGH vector, injected into the anterior left ventricular wall under direct visualization through a left lateral thoracotomy, as previously described (Aikawa et al., 2002). Blood samples were taken from the retro-orbital vein of anesthetized mice, and the plasma human growth factor (hGH) concentrations were determined by enzyme-linked immunosorbent assay (ELISA) as described (Aikawa et al., 2002).
Chromatin immunoprecipitation assay
Six-well plates seeded with H9C2 and C2C12 cells at 1 × 106 cells per well were grown to 80–90% confluence, and infected with rAAV-CMV-lacZ or rAAV-MHC-hGH vector at 500 VG/cell. Four hours after infection, cells were washed twice with phosphate-buffered saline (PBS) and fixed with 1% formaldehyde at room temperature for 10 min. After being cross-linked, soluble chromatin was made as described (Braunstein et al., 1993; Chen et al., 1999). Anti-CREB or anti-p300 (cat. nos. sc-58 and sc-585, respectively; Santa Cruz Biotechnology) or control rabbit IgG (cat. no. sc-3871; Santa Cruz Biotechnology) was used for immunoprecipitation, using protein A beads (P1052; Sigma-Aldrich), and the precipitate was analyzed by polymerase chain reaction (PCR). The primers were used to amplify a 286-bp fragment of the rAAV-CMV-lacZ genome (Iwakura et al., 2005). For the rAAV-MHC-lacZ genome, the MHC-344 primer and the MHC+ 19 primer were used (Aikawa et al., 2002).
Statistical analysis
The mean and standard error (SE) were determined for replicate samples. For multiple treatment groups, a factorial analysis of variance was applied followed by the Fisher least significant difference test as computed by the StatView computer program. Values of p less than 0.05 were considered significant.
Results
Overexpression of CREB-S133A suppresses rAAV transduction in vivo
To analyze the role of CREB in rAAV transduction we used a previously published murine cardiomyopathy model induced by overexpression of the dominant-negative mutant CREB-S133A in the heart under the control of the tissue-specific α-myosin heavy chain (MHC) promoter (dnCREB mouse). Overexpression of nonphosphorylated CREB-S133A in the mouse heart inhibits CREB-mediated transcription activation (Fentzke et al., 1998). We verified the CREB phosphorylation state in the dnCREB mouse in skeletal muscle compared with cardiac muscle. Phosphorylated CREB protein was detected in skeletal muscle of dnCREB mice, but not in heart (Fig. 1a). By comparison, phosphorylated CREB was detected in both heart and skeletal muscle of wild-type mice (data not shown). To examine the role of CREB in transduction of muscle by rAAV we used four rAAV2 vectors with two different promoters paired with two different transgenes: rAAV-MHC-hGH or lacZ and rAAV-CMV-hGH or lacZ (Aikawa et al., 2002). We injected these vectors encoding human growth hormone (hGH) at a dose of 1 × 1010 VG per animal directly into the anterior left ventricular heart wall of wild-type and dnCREB mice. hGH was used as a secreted marker of rAAV expression for better quantitation of rAAV transduction. After injecting rAAV-MHC-hGH and rAAV-CMV-hGH, into the cardiac muscle of normal mice, the serum level of hGH remained stable for at least 12 weeks (Fig. 1b), similarly to previous results (Aikawa et al., 2002). At the same time we found that neither rAAV-CMV-hGH nor rAAV-MHC-hGH injection resulted in elevated serum hGH levels in hearts of dnCREB mice (Fig. 1b), indicating that overexpression of CREB-S133A in the heart tissue is capable of inhibiting rAAV gene expression.
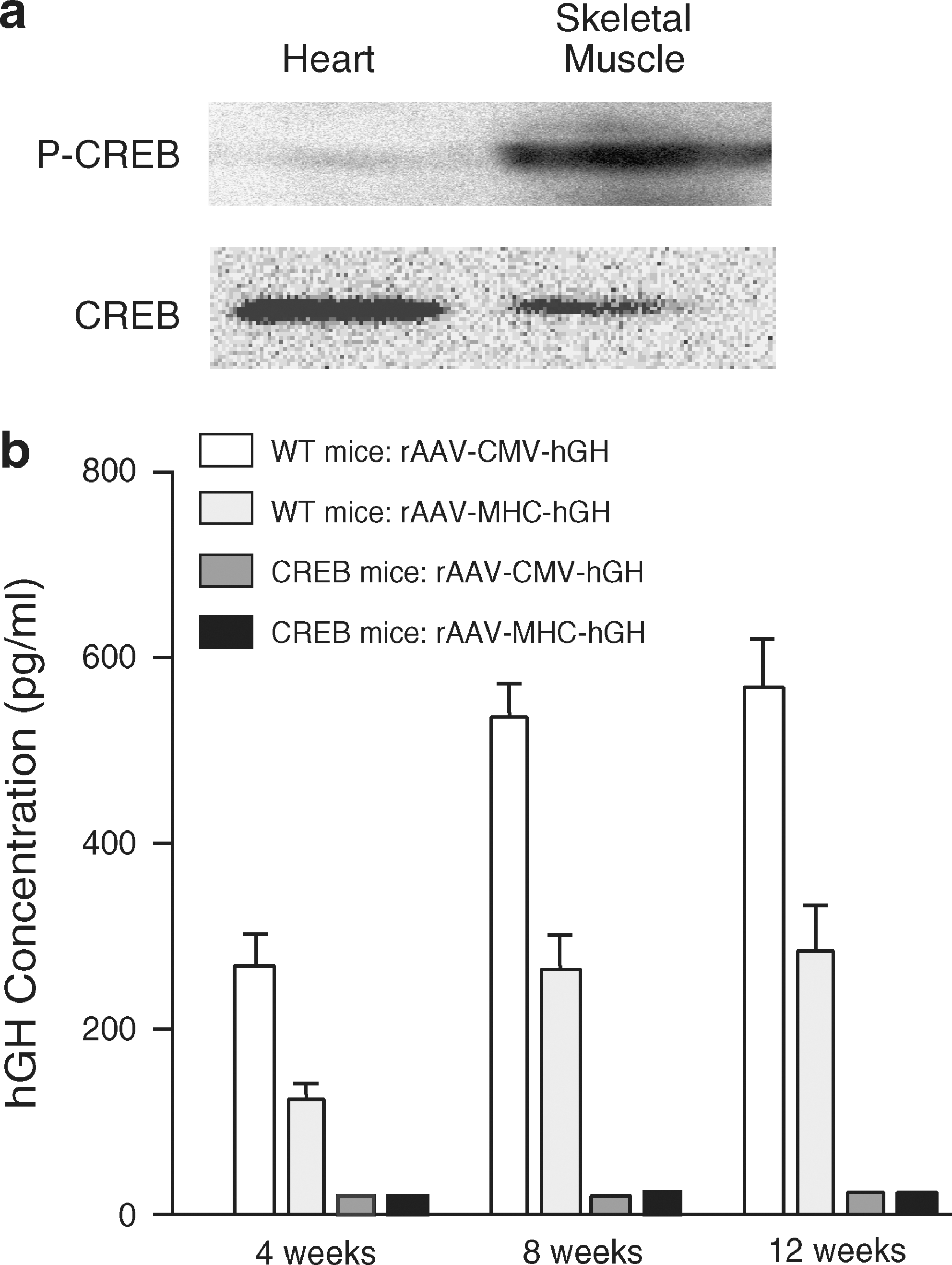
Transgenic overexpression of dominant negative CREB in the heart suppresses rAAV transduction in vivo. (
Isoproterenol increases rAAV transduction through CREB
Because a β-adrenergic receptor stimulator, isoproterenol (ISO), can activate CREB protein (Yamamoto et al., 1988), we examined whether upregulation of CREB by ISO can increase rAAV transduction. First, we observed that rAAV transduction was significantly inhibited after reducing CREB gene expression with siRNA in C2C12 cells (Fig. 2a). Next, pretreatment with ISO enhanced rAAV2 transduction in C2C12 cells more than 5-fold (Fig. 2b). When CREB expression is specifically downregulated by siRNA-CREB before rAAV2 infection and ISO treatment, lacZ transgene expression was downregulated to the baseline level (Fig. 2b).
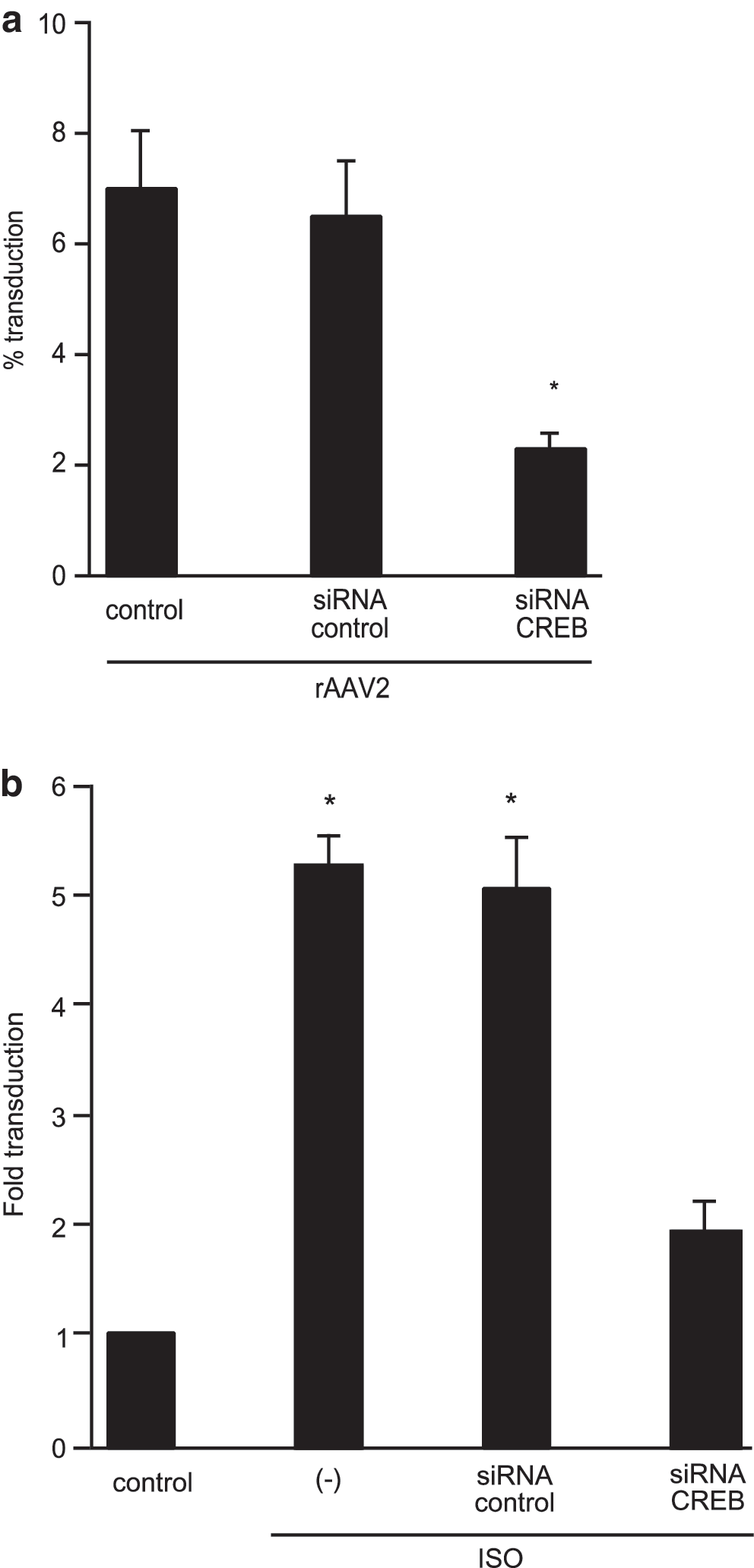
Isoproterenol (ISO) increases rAAV transduction through CREB. C2C12 cells were pretreated with control siRNA or CREB siRNA followed by infection with rAAV-CMV-lacZ at 1000 VG/cell, with or without preincubation with 10−6 M ISO. Twenty-four hours after infection, the cells were stained for β-Gal activity. Data represent means ± SE, *p < 0.01 (n = 4).
Presence of CRE sites within rAAV vector genomes
Published studies have shown the importance of CRE sites within the cytomegalovirus (CMV) promoter for transcriptional activation of a transgene situated after this internal promoter in the context of an adenoviral vector (Wheeler and Cooper, 2001). Therefore we searched the rAAV genomes used in the present study for the presence of CREB-binding sites (CRE). Unlike the CMV promoter, there are no typical CRE sites (TGACGTCA) or CRE half-sites (TGACG) to which CREB can bind (Sands and Palmer, 2008) in the MHC promoter (Molkentin et al., 1996; Gupta et al., 1998). The inverted terminal repeat (ITR) of rAAV2 and the poly(A) sequences do not include a consensus CRE site (Table 1). We then looked for the presence of a noncanonical CRE site (TGTCA) to which CREB is capable of binding to induce at levels up to 30% activity of a full CRE site (DiNardo et al., 2001). As shown in Table 1, two noncanonical CRE sites exist in the MHC promoter and the poly(A) sequence. Both lacZ and hGH transgenes used in this study harbor CRE half-sites and noncanonical CRE sites (Table 1) that are capable of CREB binding. Thus multiple sites are available in these vectors for CREB binding.
Abbreviations: CMV, cytomegalovirus; CRE, cyclic AMP response element; hGH, human growth hormone; ITR, inverted terminal repeat; MHC, major histocompatibility complex; poly (A), polyadenylation site.
The presence of the sites was determined for plus and minus DNA strands.
These half-sites are not included in the CRE site count.
rAAV mildly induces CREB phosphorylation and both CREB and p300 physically interact with the rAAV vector genome
To determine whether rAAV influences serine-133 CREB phosphorylation, CREB phosphorylation was analyzed by Western blot analysis after infection of H9C2 and C2C12 with rAAV2-CMV-lacZ at 2000 VG/cell. CREB phosphorylation was mildly but significantly increased at 15 and 30 min after rAAV infection. This effect required intact viral particles because boiled rAAV did not increase CREB phosphorylation (Fig. 3a). A study was carried out to determine whether rAAV-stimulated CREB phosphorylation could stimulate the CREB-dependent transcription of a CRE-regulated luciferase reporter plasmid (CRE-luc). After transfection with CRE-luc, H9C2 and C2C12 cells were infected with increasing amounts of rAAV vector or treated with the PKA activator forskolin. Infection with rAAV at 1000 and 2000 VG/cell significantly increased luciferase activity, and as expected, forskolin strongly activated the CRE-luc reporter plasmid (Fig. 3b).
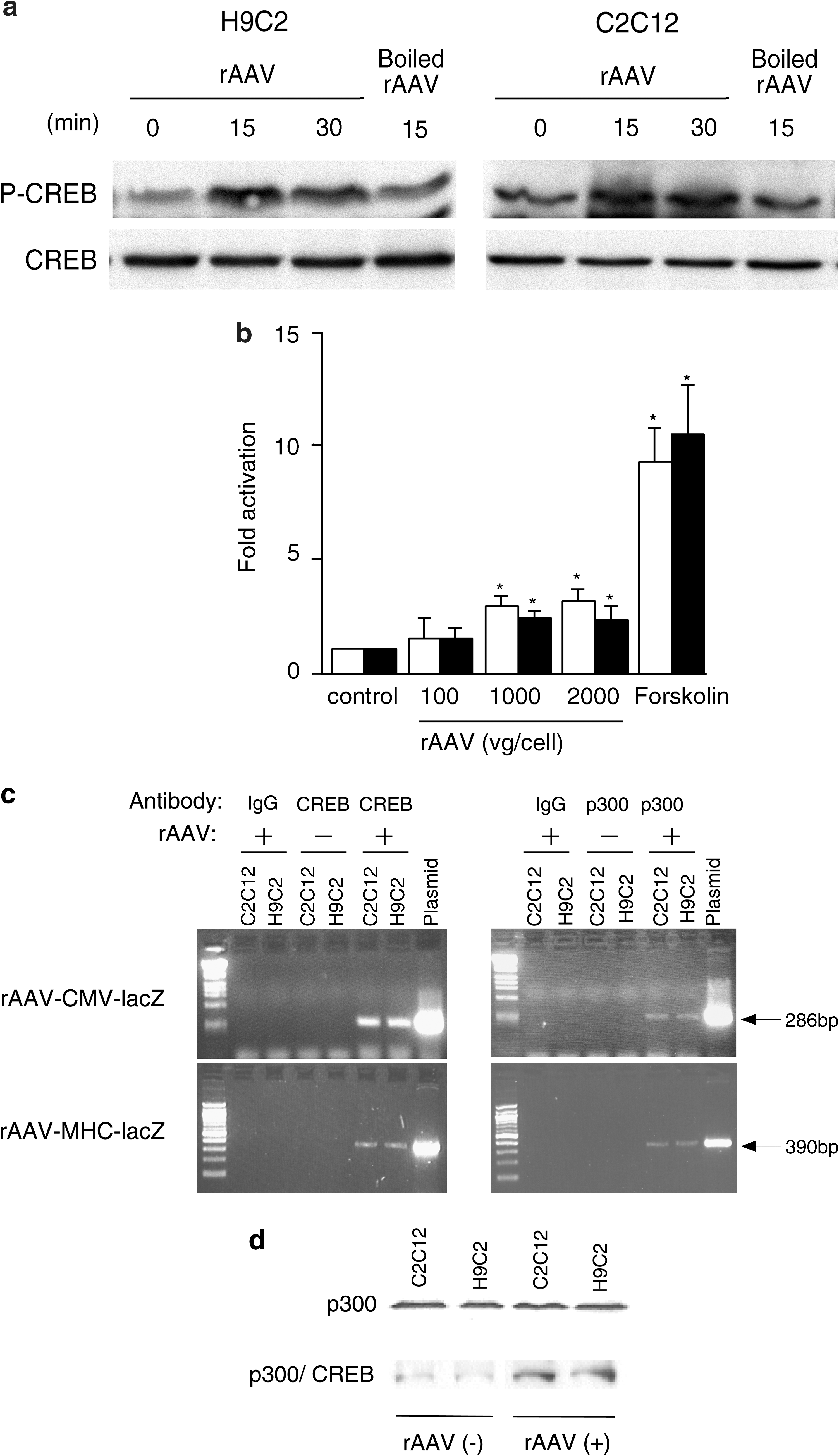
Activation of CREB by rAAV and interaction of CREB and p300 with rAAV. (
To investigate further the possible role of CREB in mediating transgene regulation, we examined whether the CREB protein physically interacts with the rAAV-CMV or rAAV-MHC genome, using chromatin immunoprecipitation (ChIP). After infection of cultured H9C2 and C2C12 cells and formaldehyde-mediated cross-linking, the rAAV genome was recovered by immunoprecipitation with an anti-CREB antibody but not with a control immunoglobulin (Fig. 3c, left). CREB phosphorylation can enhance recruitment of CREB-binding protein and p300 cofactors to enhance transcription (Goodman and Smolik, 2000). To investigate the association of p300 with the rAAV genome, we performed the ChIP assay with an anti-p300 antibody. These studies confirmed p300 association with rAAV-CMV-lacZ or rAAV-MHC-lacZ (Fig. 3c, right), which could result via the interaction of the DNA-binding intermediary transcription factor CREB. Serine-133-phosphorylated CREB activates transcription through increased binding to coactivator proteins CBP and p300 (Goodman and Smolik, 2000). These results are consistent with CREB binding to CRE sites located in the rAAV genome and recruitment of p300.
To assess whether rAAV-induced CREB phosphorylation can increase the association of CREB with p300 we investigated the interaction of CREB with p300 after rAAV infection. Although the amount of p300 immunoprecipitated from H9C2 and C2C12 cells was not influenced by rAAV infection (Fig. 3d, top), a greater amount of CREB protein was found in the p300 immunoprecipitate 24 hr after rAAV infection (Fig. 3d, bottom).
Discussion
Molecular mechanisms important for wild-type AAV infection, latency, and viral gene expression are relevant to recombinant vector-mediated gene therapy. To explore the role of CREB in rAAV transduction we used CREB-S133A-expressing mice. Transduction using rAAV regulated by either a tissue-specific (MHC) or promiscuous promoter/enhancer (CMV) was blocked in the dnCREB mouse (Fig. 1b). Importantly, in dnCREB transgenic mice, the same MHC promoter is driving CREB-S133A expression, and only rAAV expression was suppressed after rAAV injection. Isoproterenol (ISO) is a potent β-adrenergic receptor activator, which can activate CREB through the PKA pathway (Yamamoto et al., 1988). Phosphorylated CREB can then recruit CBP and p300, which have HAT activity, and increase promoter accessibility by acting on chromatin structure (Gusterson et al., 2002). Pretreatment with 10−6 M ISO significantly augmented rAAV transduction (Fig. 2a) and we observed that the ISO effect on rAAV transduction occurred in a dose-dependent manner between 10−7 and 10−5 M without an increase in vector genomes per cell (Nattonson et al., unpublished data). ISO-augmented rAAV transduction was strongly inhibited by siRNA-CREB treatment (Fig. 2b), suggesting that CREB plays a role in CMV-driven transgene expression. Those experiments including loss of CREB function (Fig. 1) and gain of CREB function (Fig. 2) revealed that CREB is important for rAAV transduction. To address the possibility that, in addition to directly acting on the vector genome, CREB may be enhancing transduction by influencing viral uptake and trafficking, a pseudotype vector in which the rAAV2 genome was packaged in an rAAV1 capsid (rAAV1/2) was tested. Similar to the rAAV2 capsid vector, transduction with the rAAV1 capsid vector was significantly blocked by dnCREB overexpression in vitro (data not shown). By inhibiting CREB protein function (dnCREB), stimulating CREB function (with ISO), or knocking down CREB gene expression (with siRNA), we have shown that CREB plays an important role in rAAV transduction. Importantly, although the CMV promoter has abundant CRE sites and the MHC promoter has limited CRE sites (as shown in Table 1), both vectors were regulated by CREB. Using the ChIP assay we demonstrate an association of CREB with the rAAV-CMV vectors and rAAV-MHC vectors (Fig. 3c). CRE, CRE half-sites, and noncanonical CRE sites are present throughout the recombinant AAV vector genomes used here, and even though most of the sites are located outside of the actual promoter sequences, concatemerization and/or circularization of the rAAV genomes may enhance the effect of CREB/p300 by bringing distal CRE sites into closer proximity to the promoter region. In addition, 1 CRE half-site and 2 noncanonical CRE sites exist in the Rous sarcoma virus (RSV) promoter and 12 additional sites are present over the length of the rAAV-RSV-LEA29Y-WPRE vector used by Penaud-Budloo and colleagues (2008) (M. Penaud-Budloo, C. Le Guiner, and P. Moullier, unpublished data). Furthermore, 18 CRE half-sites or noncanonical CRE sites exist over the length of the wild-type AAV2 genome, 26 sites in AAV1, 29 sites in AAV4, 24 sites in AAV5, and 22 sites in AAV8, and some of these sites are located proximal to the P5 and P40 promoters (including a full CRE site at the AAV5 P40 promoter) (data not shown), indicating a role for CREB in the wild-type AAV life cycle. However, other transcription factors that are capable of binding CRE elements may have a role too, including CREM (cAMP response element modulator) and ATF-1 (activating transcription factor-1), and CREB can bind to CRE as a homodimer or heterodimer with other transcription factors belonging to the CREB/ATF or Jun/Fos families (Hai and Curran, 1991). CREB phosphorylation can enhance recruitment of CBP and p300 cofactors to establish transcriptionally active chromatin (Roth et al., 2001). It is also possible that CREB influences rAAV transduction via an as yet unknown indirect mechanism. As shown in Fig. 3a, the enhanced phosphorylation of CREB starts as early as 15 min post-AAV infection, which is a time point after infection when AAV particles are detected in the nucleus of about 50% of cells grown in vitro (Seisenberger et al., 2001). The rapid appearance of AAV particles in the nucleus may trigger CREB phosphorylation by activating signal transduction pathways in addition to DNA binding.
The ChIP assay carried out with a p300 antibody revealed an association with both the rAAV-CMV and rAAV-MHC genomes (Fig. 3c). Because p300 is able to interact with the rAAV genome via CREB (Fig. 3d), CREB may be functioning as a transcription regulator either by itself or in association with as yet unidentified factors. It is possible that p300 acetylates rAAV-associated histones to establish a transcriptionally active chromatin structure, which may relate to findings demonstrating that the rAAV genome is converted to circular episomal chromatin with a typical nucleosomal pattern in vivo in the skeletal muscle of the nonhuman primate (Penaud-Budloo et al., 2008). Accordingly, CREB involvement may be a general feature of rAAV vector expression in muscle cells.
HATs may be required to establish rAAV open chromatin, which is the basis of long-term gene expression in muscle. In agreement with our observations, Okada and colleagues demonstrated an association of acetylated histone H3 with the rAAV vector genome in cancer cells in vitro (Okada et al., 2006). In vitro, the vector may exist in a circular episomal state (Duan et al., 1999) or integrated into the host chromosome where it can be repressed, but reactivated with histone deacetylase inhibitors (Chen et al., 1997; Chen and Townes, 2000). Previous reports show that p300 and CBP have similar tissue patterns of HAT activity, with the highest level in the brain; a relatively high level in the lung, spleen, and heart; an intermediate level in testes and skeletal muscle; and a lower level in liver and kidney (Li et al., 2002). The data presented in this report support a role for CREB and HATs in the regulation of rAAV vector chromatin in muscle cells.
Footnotes
Acknowledgments
The authors thank Dan Eisenberg and Serban Georgescu for technical assistance, and John Lepore for construction of CRE-luciferase plasmids. Magalie Penaud-Budloo and Philippe Moullier provided unpublished data. This work was supported by the Uehara Memorial Foundation and Starr Foundation (R.A.), by grant R01 HL54592 (G.S.H.) from the National Institutes of Health, and by Association Française contre les Myopathies (AFM) award 12263 to R.O.S.
Author Disclosure Statement
R.O.S. is an inventor on patents related to recombinant AAV technology and owns equity in a gene therapy company that is commercializing AAV for gene therapy applications. To the extent that the work in this manuscript increases the value of these commercial holdings R.O.S. has a conflict of interest.