
Abstract
Excessive angiogenesis mediated by vascular endothelial growth factor (VEGF) plays an important role in angioproliferative ocular diseases. We have previously developed a large animal model for these diseases by intravitreal adenoviral gene transfer of VEGF-A165. 15-Lipoxygenase-1 (15-LO-1), an oxidizing enzyme producing reactive lipid hydroperoxides, has been shown to induce aberrant angiogenesis in cancer models of transgenic mice overexpressing human 15-LO-1. Our purpose was to study the effects of 15-LO-1 on VEGF-A165-induced angiogenesis in New Zealand White rabbit eyes, using intravitreal adenovirus-mediated gene transfers. AdCMV and Adh15-LO-1 alone served as controls. As determined by immunohistochemistry, VEGF-A165 significantly increased the number and size of the capillaries in various compartments of the eyes. 15-LO-1 efficiently inhibited VEGF-A165-induced neovascularization and pathological changes by reducing VEGF-A165 mRNA and protein expression, determined by RT-PCR, ELISA, and immunohistochemistry. 15-LO-1, which produces endogenous ligands for peroxisome proliferator-activated receptor-γ (PPARγ), also prevented VEGF-A165-induced expression of PPARγ and VEGF receptor-2, as measured by quantitative RT-PCR. In conclusion, our findings show that 15-LO-1 prevents VEGF-A165-induced angiogenesis and consequent pathology in the eyes, suggesting that intravitreal 15-LO-1 gene transfer could be a potential new strategy for the treatment of neovascular complications in the eyes.
Introduction
Vascular endothelial growth factors (VEGFs) play a pivotal role in physiological angiogenesis, but they also induce excessive angiogenesis in many pathological conditions, such as ocular diseases. The most important angioproliferative ocular diseases are diabetic proliferative retinopathy and age-related macular degeneration (Witmer et al., 2003). Our group has previously developed a model for ocular angioproliferative diseases by showing that intravitreal adenoviral gene transfer of VEGF-A165 induces neovascularization and increases vascular leakage, resembling structural changes detected in diabetic retinopathy (Kinnunen et al., 2006).
In this work, we show that 15-LO-1 efficiently prevents angiogenic effects induced by VEGF-A165. 15-LO-1 prevented VEGF-A165-induced increase in the number and size of the capillaries in various compartments of the eyes, thus inhibiting VEGF-A165-induced neovascularization and pathological changes. VEGF-A165 expression was prevented by 15-LO-1 at the mRNA level, resulting in low production of VEGF-A protein. VEGF-A mediates its signaling effects mainly via VEGF receptor (VEGFR)-2, which is induced by peroxisome proliferator-activated receptor (PPAR)-γ. We show that the mRNA expression of PPARγ and VEGFR2 is induced by VEGF-A165, but that the inductions are efficiently inhibited by 15-LO-1. Thus, 15-LO-1 gene transfer is a potential new strategy for the treatment of pathological angiogenesis in the eyes.
Materials and Methods
Production of recombinant adenoviruses
The expression cassette of recombinant adenovirus Adh15-LO-1 contains nucleotides 1–2245 of human 15-LO-1 cDNA (Sigal et al., 1988) under the control of the human cytomegalovirus (CMV) immediate-early enhancer/promoter and intron with splicing signals, followed by bovine growth hormone transcriptional terminator and poly(A) signal sequences (Aggarwal et al., 2007). AdhVEGF-A165 (Rissanen et al., 2003b) contains the CMV immediate-early promoter/enhancer region, the coding sequence for human VEGF-A165, and the human growth hormone poly(A) signal. Recombinant adenoviruses containing nuclear-targeted Escherichia coli lacZ gene (AdlacZ) (Laitinen et al., 1998) or the CMV promoter without any insert (AdCMV) (Kinnunen et al., 2006) were used as controls in the in vitro and in vivo gene transfers, respectively. All recombinant adenoviruses were produced in 293 cells (Graham et al., 1977) by homologous recombination as described (Laitinen et al., 1998).
15-LO-1 activity assay
The capacity of the recombinant Adh15-LO-1 virus to produce functionally active 15-LO-1 protein was verified from in vitro-transduced ECV304 cells 72 hr after transduction by gas chromatographic hydroxy fatty acid analysis as described (Viita et al., 2008).
Intravitreal gene transfer into rabbit eyes
New Zealand White rabbits (n = 32) were anesthetized as described (Kinnunen et al., 2006). The animals received intravitreal injections of a total of 1.0 × 1011 viral particles (AdCMV, Adh15-LO-1, AdhVEGF-A165, and a combination of Adh15-LO-1 with AdhVEGF-A165 [treatment group]) in a total volume of 100 μl. The injections were performed intravitreally (Kinnunen et al., 2006) by injecting AdCMV alone (1 × 1011 viral particles [VP]) and AdhVEGF-A165 (0.5 × 1011 VP) plus AdCMV (0.5 × 1011 VP), or Adh15-LO-1 (0.5 × 1011 VP) plus AdCMV (0.5 × 1011 VP) and the combination of Adh15-LO-1 (0.5 × 1011 VP) and AdhVEGF-A165 (0.5 × 1011 VP) into the left and right eyes of the same animal, respectively. The animals were killed 6 days after the gene transfers, when the concentration of the transduced human VEGF-A protein was maximal in the vitreous humor (Kinnunen et al., 2006). All animal experiments were approved by the Experimental Animal Committee of the University of Kuopio (Kuopio, Finland).
Fundus photographs of rabbit eyes were taken before gene transfer and 1 day before sacrifice, and vitreous samples were collected from each eye after sacrifice as described (Kinnunen et al., 2006). One set of animals (n = 5 per group) was perfusion fixed (Rissanen et al., 2003a). The eyes were enucleated and cut in half. One half was immersion fixed in 4% paraformaldehyde in 7.5% sucrose (pH 7.4) for 4 hr, rinsed in 15% sucrose (pH 7.4) overnight, and embedded in paraffin (Kinnunen et al., 2006). Another set of animals (n = 5 per group) was killed without perfusion fixation. The eyes were enucleated and cut in half from the side of the optic nerve. The half containing the optic nerve was processed for frozen sections and the other half was cut into longitudinal segments, which contained parts from the back of the eye all the way to the cornea, that is, parts of the outer layer (sclera and cornea), the middle layer (choroid, ciliary body, and the iris), and the innermost layer (the retina). The segments were snap-frozen in liquid nitrogen and stored at −70°C for RNA analysis. A third set of animals (n = 6 per group) was killed without perfusion fixation, the superior part of the eyes was identified for orientation, the eyes were enucleated, and the retinal whole mounts were detached from the sclera and postfixed for 24 hr in the same fixative solution.
Immunohistochemistry
Immunohistochemical analysis of capillaries was performed on serial paraffin-embedded sections, using a mouse monoclonal antibody against human CD31 (diluted 1:50; Dako, Glostrup, Denmark). Capillary mean area (μm2) and capillary density (capillaries per mm2) in the optic nerve head, retina, and anterior segments were measured as described (Kinnunen et al., 2006). Expression of transduced VEGF-A protein was detected by immunohistochemistry as described (Viita et al., 2008).
Whole mounts of retinas were incubated with fluorescein isothiocyanate (FITC)-conjugated lectin (Lycopersicon esculentum [tomato], diluted 1:200; Sigma-Aldrich, St. Louis, MO) overnight at room temperature. The retinas were then washed in 0.05 M Tris-buffered saline (pH 7.4) containing 0.5% Triton, three times for 15 min, and incubated with mouse monoclonal anti-glial fibrillary acidic protein (GFAP, diluted 1:1000; Sigma-Aldrich) for 24 hr at room temperature and with goat anti-mouse IgG (Alexa Fluor 594, diluted 1:500; Molecular Probes/Invitrogen, Eugene, OR) for 3 hr at room temperature. The stained retinas were flat mounted on a glass slide in glycerol and coverslipped.
Measurements of retinal blood vessels
Four peripheral areas of medullary rays from each whole-mounted retina were digitally imaged with a DP50 camera system (Olympus, Tokyo, Japan) operating on a BX40 microscope (Olympus). Retinal blood vessel diameters were measured with Adobe Photoshop CS3 software (Adobe Systems, San Jose, CA).
Human VEGF-A enzyme-linked immunosorbent assay
Production of transduced human VEGF-A protein was quantified by enzyme-linked immunosorbent assay (ELISA) (Quantikine human VEGF-A; R&D Systems, Minneapolis, MN) from snap-frozen vitreous samples according to the manufacturer's instructions. The results are presented as picograms of VEGF-A per milliliter.
Reverse transcription-polymerase chain reaction (RT-PCR) and quantitative RT-PCR
Total RNA was isolated and RT-PCR for transduced human VEGF-A, transduced human 15-LO-1, and endogenous rabbit α-actin was performed as described (Viita et al., 2008). The primers for human 15-LO-1 are specific for the transferred gene, because the 5′ primer binds to the CMV promoter area of the adenoviral vector 5′ to the splice signals and the 3′ primer binds to the cloning junction of 15-LO-1. These primers result in the amplification of a 956-bp fragment from the unspliced Adh15-LO-1 vector and of a 129-bp fragment from the spliced mRNA produced from the recombinant Adh15-LO-1 virus. Quantitative RT-PCR was used to quantify the mRNA expression of endogenous rabbit PPARγ and VEGFR2 as described (Viita et al., 2008). Twenty-five and 2.5 ng of cDNA were used as the template for PPARγ and VEGFR2 reactions, respectively.
Statistics
Results are presented as means ± SD. Statistical significance was evaluated with the GraphPad Prism 4.00 software package, using one-way analysis of variance (ANOVA) followed by the Newman–Keuls multiple comparison test or the Kruskal–Wallis test when necessary. A value of p < 0.05 was considered statistically significant.
Results
Adh15-LO-1 produces enzymatically active 15-LO-1
Verification of the functionality of the recombinant Adh15-LO-1 virus was confirmed from in vitro-transduced ECV304 cells by gas chromatographic enzyme activity analysis after incubation of the cells with linoleic acid. Production of the specific 15-LO-1 reaction product 13-hydroxyoctadecadienoic acid (13-HODE) was highly increased in Adh15-LO-1-transduced cells 72 hr after transduction as compared with untransduced or AdlacZ-transduced cells (Fig. 1). The proportion of 13-HODE of all the HODEs detected was 77% in Adh15-LO-1-transduced cells.
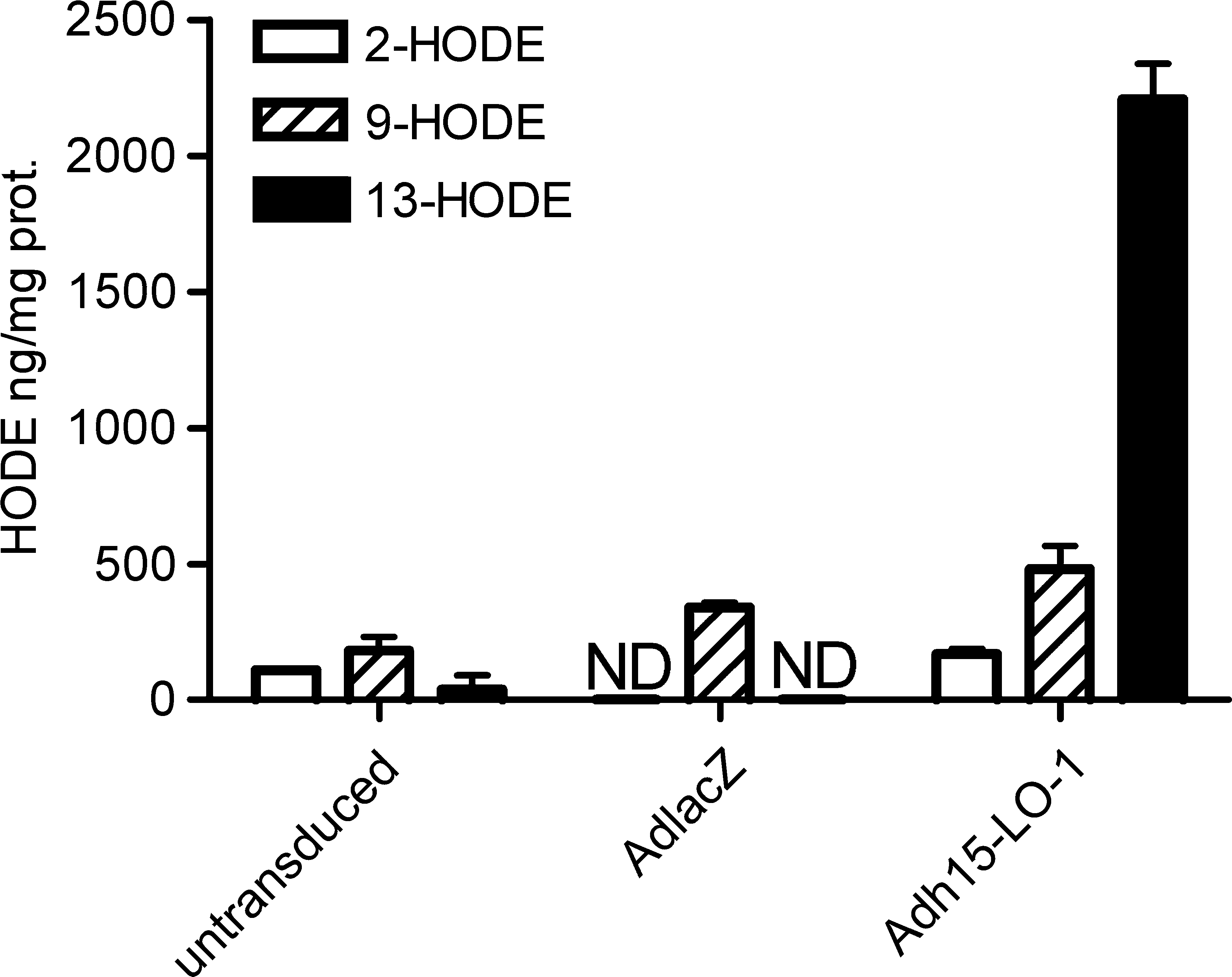
Gas chromatographic 15-lipoxygenase-1 (15-LO-1) enzyme activity analysis from in vitro-transduced ECV304 cells after incubation with linoleic acid. Untransduced, AdlacZ-transduced, and Adh15-LO-1-transduced cells were lysed and incubated with 50 μM linoleic acid for 15 min at 37°C. Results are presented as nanograms of hydroxyoctadecadienoic acid (HODE) per milligram of protein ± SD, n = 2 per group. ND, not detectable.
Adh15-LO-1 prevents AdhVEGF-A165-induced capillary enlargement and an increase in capillary number
Fundus photographs from the rabbit eyes 5 days after the gene transfer showed highly induced neovascularization in AdhVEGF-A165-transduced eyes compared with all other groups (Fig. 2). Capillary analysis of CD31-immunostained paraffin sections showed that AdhVEGF-A165 gene transfer resulted in a significant enlargement of the capillaries in the papilla (2.1-fold vs. control AdCMV; p < 0.001) (Fig. 3a–d and m) and in the retina (2.6-fold vs. control AdCMV; p < 0.01) (Fig. 3e–h and n). In the 15-LO-1 treatment group, where Adh15-LO-1 gene transfer was combined with AdhVEGF-A165, Adh15-LO-1 prevented AdhVEGF-A165-induced capillary enlargement (p < 0.001 in the papilla and p < 0.05 in the retina) (Fig. 3m and n). No effect on the size of the capillaries was detected in the anterior segments (Fig. 3i–l and o). AdhVEGF-A165 gene transfer significantly increased the average number of capillaries in the papilla (3.1-fold vs. control AdCMV; p < 0.001) (Fig. 3a-d and p), and in the anterior segments (3.3-fold vs. control AdCMV; p < 0.001) (Fig. 3i–l and r), whereas Adh15-LO-1 prevented the increase in capillary number (p < 0.001 in the papilla and p < 0.001 in the anterior segment) (Fig. 3p and r). No effect on the number of capillaries was detected in the retina (Fig. 3e–h and q). Adh15-LO-1 alone had no effect on mean capillary area or capillary number (Fig. 3m–r).

Adh15-LO-1 prevents AdhVEGF-A165-induced vascular effects. Top: Rabbit fundus photographs before the gene transfers (pre) show that the eyes were calm in all groups. Bottom: Five days after the gene transfers (post) the eyes remained fairly calm in the AdCMV, Adh15-LO-1 plus AdCMV, and Adh15-LO-1 plus AdhVEGF-A165 gene transfer groups, whereas increased neovascularization detected in the AdhVEGF-A165 plus AdCMV gene transfer group made evaluation of the retinal vessels difficult.

Adh15-LO-1 prevents AdhVEGF-A165-induced capillary enlargement and increase in capillary number. Immunohistochemical staining for CD31 was done in papilla, retina, and anterior segment. (
Retinal blood vessel diameter was measured from whole-mounted retinas immunostained with FITC-conjugated lectin. Intravitreal injections of AdhVEGF-A165 alone induced a significant increase in retinal blood vessel diameter (1.9-fold vs. control AdCMV; p < 0.001), whereas in the 15-LO-1 treatment group, Adh15-LO-1 prevented the capillary enlargement (p < 0.001) (Fig. 4a–d and i). No major differences between the study groups were observed in glial fibrillary acidic protein staining (Fig. 4e–h). However, intravitreal injections caused microglial activation in all study groups (data not shown).

Adh15-LO-1 blocks the AdhVEGF-A165-induced increase in retinal blood vessel diameter. Immunohistochemical staining of blood vessels (
Adh15-LO-1 reduces the expression of transduced VEGF-A165 mRNA, resulting in a significant reduction in the production of VEGF-A protein
RT-PCR showed that expression of transduced human VEGF-A165 mRNA was reduced in 15-LO-1 treatment group samples compared with AdhVEGF-A165-transduced eyes (Fig. 5a). Expression of transduced, spliced 15-LO-1 mRNA was clearly seen as a 129-bp product both in Adh15-LO-1 group samples and in 15-LO-1 treatment group samples (Fig. 5b). 15-LO-1 RT-PCR also showed that there was no contaminating DNA, because the amplified PCR fragment from the 15-LO-1 adenoviral vector DNA would have been differently sized (956 bp). Rabbit α-actin RT-PCR showed equal expression in all samples (Fig. 5c).
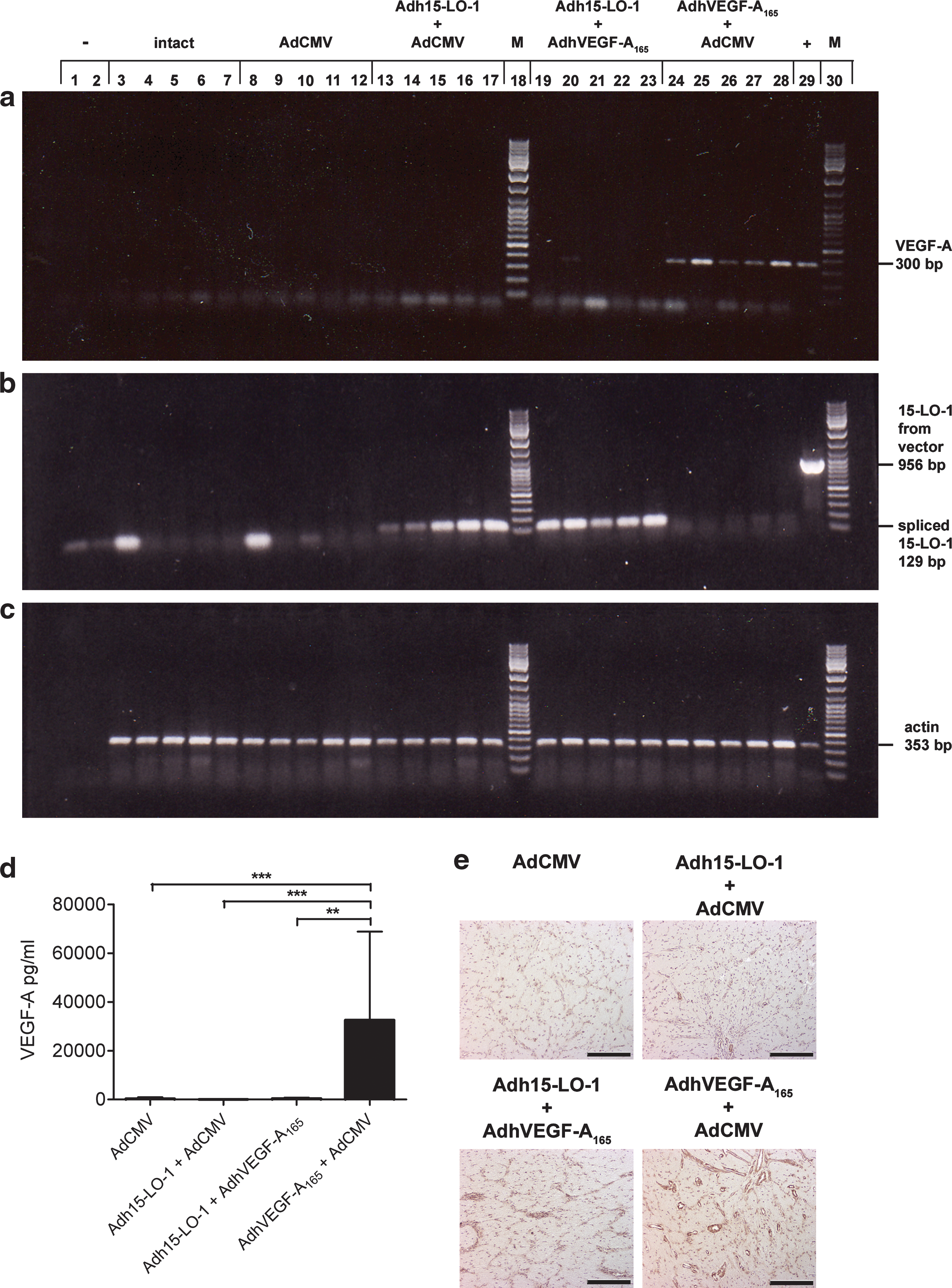
Adh15-LO-1 reduces mRNA expression and prevents protein production of transduced human VEGF-A165. Shown is the RT-PCR analysis of (
The reduced expression of transduced human VEGF-A165 transcript was also reflected in the human VEGF-A165 ELISA. AdhVEGF-A165 gene transfer resulted in a large induction (p < 0.001) in the production of human VEGF-A165 protein in the vitreous humor compared with control AdCMV (Fig. 5d), whereas in the 15-LO-1 treatment group the levels did not differ from the control AdCMV samples or the Adh15-LO-1-transduced samples. The reduction in VEGF-A protein expression was also detected by immunohistochemistry (Fig. 5e).
AdhVEGF-A165-induced mRNA expression of endogenous rabbit PPARγ and VEGFR2 is prevented by Adh15-LO-1
Quantitative RT-PCR analysis was used to measure expression levels of endogenous rabbit PPARγ and VEGFR2 mRNAs in the eye samples. AdhVEGF-A165 gene transfer resulted in a 2.9-fold increase in PPARγ (Fig. 6a) and a 2.5-fold increase in VEGFR2 (Fig. 6b) mRNA expression compared with the control AdCMV group. In the 15-LO-1 treatment group these inductions were efficiently prevented.
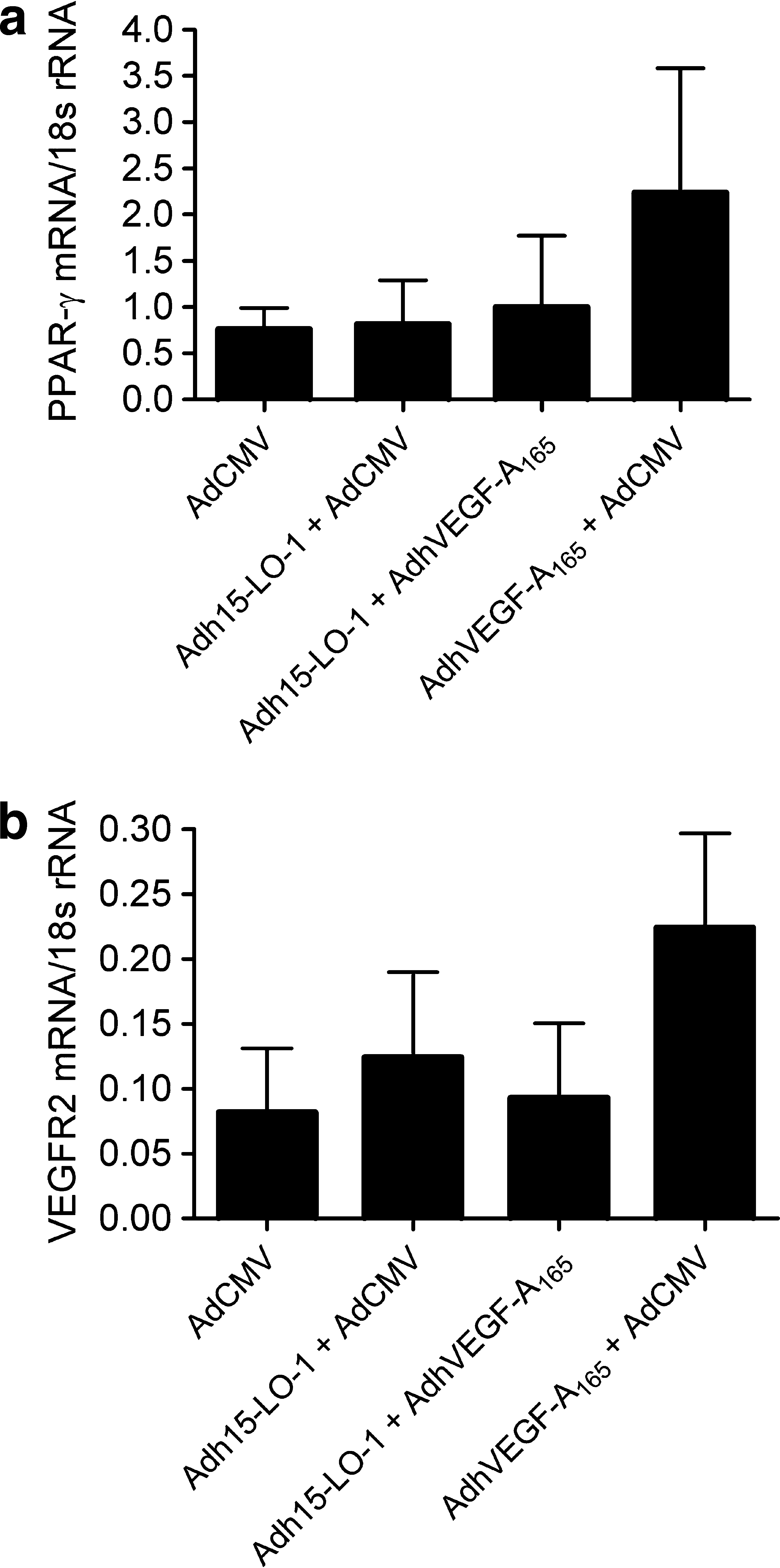
Adh15-LO-1 prevents AdhVEGF-A165-induced mRNA expression of peroxisome proliferator-activated receptor-γ (PPARγ) and vascular endothelial growth factor receptor-2 (VEGFR2). Shown is a quantitative RT-PCR analysis of PPARγ and VEGFR2 expression from the transduced eye samples. (
Discussion
We have previously shown that intravitreal adenoviral gene transfer of VEGF-A165 induces neovascularization and increases vascular leakage in the rabbit eye (Kinnunen et al., 2006), thus producing a large animal model for VEGF-A165-dependent angioproliferative diseases of the eye. Transductions with 1010, 5 × 1010, and 1011 VP of AdhVEGF-A165 result in a dose-dependent increase in the production of human VEGF-A protein in the vitreous humor. Histological analyses reveals that significant differences in vascular structures already occur with 1010 VP of AdhVEGF-A165 and that an inflammatory reaction is seen with the highest dose of 1011 VP. The dose of 5 × 1010 VP of AdhVEGF-A165 was selected for this study to achieve maximal expression of transduced human VEGF-A protein in the vitreous humor, while avoiding any adverse inflammatory effects. AdCMV control virus was used to bring the total number of viral particles to 1011 VP in each group. The animals were killed 6 days after gene transfer, because our previous study showed that the amount of transduced human VEGF-A protein was the highest at this time point and the histological analyses showed equally strong responses 6 days and 2 weeks after gene transfer (Kinnunen et al., 2006).
The expression of transduced VEGF-A165 was already reduced by 15-LO-1 at the mRNA level. 15-LO-1 might affect the production of transduced human VEGF-A165 either by preventing transcription or by destabilizing the transcript, or both. Both production and stability of VEGF-A165 mRNA are highly regulated. Our AdhVEGF-A165 construct lacks all the regulatory elements of the endogenous human VEGF-A165, as it contains only the protein coding region of VEGF-A165 under the control of the CMV promoter. Because the human 15-LO-1 cDNA in our adenoviral construct is also under the control of the CMV promoter, and there was no reduction in the expression of transduced human 15-LO-1 in the treatment group, it is highly unlikely that the reduction in mRNA expression of VEGF-A165 would be caused by blocking the CMV promoter. Thus, the most likely explanation is that 15-LO-1 expression and/or its lipid hydroperoxide reaction products led to destabilization of the VEGF-A165 transcript by an as yet unidentified mechanism that would affect the coding sequence of VEGF-A165.
The VEGF-A165-induced expression of both PPARγ and VEGFR2 was prevented in the 15-LO-1 treatment group. The 15-LO-1 product 13-HODE is an endogenous activator and ligand of PPARγ (Nagy et al., 1998). Interestingly, it has been shown in a rabbit eye model that PPARγ binding to the VEGFR2 promoter induces VEGFR2 expression, but ligand binding to PPARγ actually results in an inhibition of VEGFR2 expression (Sassa et al., 2004). PPARγ ligands have also been shown to inhibit choroidal (Murata et al., 2000) and corneal neovascularization (Xin et al., 1999; Panigrahy et al., 2002; Sarayba et al., 2005). We hypothesize, that 15-LO-1 affects VEGF-A165-induced neovascularization both in transduced cells and via its secreted products. Thus, 15-LO-1 expression in cells next to the vascular endothelial cells (ECs) directly inhibits VEGF-A165 expression and prevents angiogenic signal transduction events mediated by secreted VEGF-A165, whereas in 15-LO-1-transduced ECs the 15-LO-1 products prevent PPARγ-induced VEGFR2 expression.
In conclusion, we have shown that intravitreal Adh15-LO-1 gene transfer prevents all angiogenic effects induced by AdhVEGF-A165 in the eyes. The mechanism for the inhibition involves prevention of the expression of VEGF-A165 at the mRNA level, resulting also in the reduced expression of PPARγ and VEGFR2. Therefore, 15-LO-1 gene transfer is a potential new treatment option for VEGF-induced pathological angiogenesis in the eyes.
Footnotes
Acknowledgments
This study was supported by grants from the University of Kuopio, the Finnish Foundation for Cardiovascular Research, the Academy of Finland, EU grant Baculogenes (LSHB-CT-2006-037541), the Orion-Farmos Research Foundation, and the Ark Therapeutics Group. The authors thank Ms. Anne Martikainen, Ms. Sari Järveläinen, Ms. Tiina Koponen, Ms. Seija Sahrio, and Ms. Marja Poikolainen for technical assistance.
Author Disclosure Statement
No competing financial interests exist.