
Abstract
Our studies have shown that coinjection of conventional single-stranded adeno-associated virus 2 (ssAAV2) vectors carrying the enhanced green fluorescent protein (EGFP) gene with self-complementary (sc) AAV2-T cell protein tyrosine phosphatase (TC-PTP) and scAAV2-protein phosphatase-5 (PP5) vectors resulted in an ∼16-fold increase in EGFP expression in primary murine hepatocytes in vivo [Jayandharan, G.R., Zhong, L., Li, B., Kachniarz, B., and Srivastava, A. (2008). Gene Ther. 15, 1287–1293]. In the present studies, this strategy was further optimized to achieve transgene expression at reduced vector/helper virus doses. These included the use of scAAV helper viruses containing (1) hepatocyte-specific promoters, (2) tyrosine-mutant AAV2 capsids, and (3) additional AAV serotype vectors known to efficiently transduce hepatocytes. The hepatocyte-specific transthyretin (TTR) promoter was ∼6- to 7-fold more efficient than the Rous sarcoma virus (RSV) promoter; tyrosine-mutant AAV2 capsids were ∼6- to 11-fold more efficient than the wild-type AAV2 capsids; and the AAV8 serotype helper virus was ∼16-fold more efficient than AAV2 serotype helper virus. With these modifications, the vector dose of the helper virus could be further reduced by ∼50-fold. Last, coadministration of scAAV8-PP5 helper virus increased coagulation factor IX expression from an ssAAV2 vector by ∼7- to 10-fold, thereby achieving therapeutic levels at lower vector doses. No adverse effect on hepatocytes was observed under any of these experimental conditions. The strategy presented here should be adaptable to any ssAAV transgene cassette and, specifically, liver-directed applications of ssAAV2 vectors containing larger genes that cannot be encapsidated in scAAV vectors.
Overview Summary
Although recombinant adeno-associated viral (AAV) vectors containing single-stranded genomes have been used in clinical trials, the single-stranded DNAs are transcriptionally inactive, and relatively large vector doses are needed to achieve expression of therapeutic genes. We have developed optimized recombinant self-complementary AAV vectors carrying the protein phosphatase-5 gene (scAAV-PP5) as helper viruses, coadministration of which leads to efficient transgene expression from conventional single-stranded AAV (ssAAV) vectors at reduced doses. These optimized helper viruses should prove useful in achieving high levels of expression at lower doses of therapeutic genes from ssAAV vectors containing large genes, such as factor VIII, which exceed the packaging capacity of scAAV vectors, for the potential gene therapy of hemophilia A.
Introduction
We have previously documented that phosphorylated forms of a 52-kDa cellular chaperone protein, FKBP52 (FK506-binding protein), interact specifically with the D-sequence within the inverted terminal repeat (ITR) of the AAV genome (Qing et al., 1997, 2001). Phosphorylation of FKBP52 at serine/threonine (Ser/Thr) and tyrosine (Tyr) residues inhibits viral second-strand DNA synthesis by ∼40 and ∼90%, respectively, leading to inefficient transgene expression (Qing et al., 1998, 2001, 2003; Zhong et al., 2004a–c). However, dephosphorylation of FKBP52 at tyrosine residues by the cellular T-cell protein tyrosine phosphatase (TC-PTP), and at serine/threonine residues by protein phosphatase-5 (PP5), prevents FKBP52 from binding to the D-sequence, leading to efficient viral second-strand DNA synthesis (Qing et al., 2003; Zhao et al., 2007). Augmented transgene expression from ssAAV2 vectors also occurs in transgenic mice overexpressing TC-PTP (Qing et al., 2003), and in mice deficient in FKBP52 (Zhong et al., 2004b). Subsequently, we have also developed scAAV-TC-PTP and scAAV-PP5 vectors (Zhong et al., 2004a; Zhao et al., 2007; Jayandharan et al., 2008). We reasoned that if scAAV-TC-PTP and scAAV-PP5 vectors are admixed with a conventional ssAAV vector before transduction, the rapid and simultaneous expression of TC-PTP and PP5 from scAAV vectors, which do not require viral second-strand DNA synthesis, would completely dephosphorylate FKBP52 at both tyrosine and serine/threonine residues, respectively. This would lead to more efficient second-strand DNA synthesis of the ssAAV vector, resulting in high-efficiency transgene expression. Indeed, this coadministration strategy led to an ∼16-fold increase in the transduction efficiency of ssAAV2 vectors in primary murine hepatocytes in vivo (Jayandharan et al., 2008).
In an effort to augment the transduction efficiency of ssAAV vectors in liver-directed gene therapy, the present study was designed to enhance the helper functions of scAAV-TC-PTP and/or scAAV-PP5 vectors by optimizing various parameters including the promoter, the AAV packaging serotype, and the helper virus dose. We also tested this helper system in an scAAV2 vector carrying a tyrosine-to-phenylalanine mutation in codon 730 of the VP3 region of the AAV2 capsid (AAV2-Y730F), shown to facilitate high-efficiency transduction of hepatocytes in vivo (Zhong et al., 2008a,b). Last, we also examined whether the optimized helper virus is capable of allowing expression of a therapeutic gene (human factor IX [F.IX]) in mouse liver at reduced vector doses.
Materials and Methods
Plasmids and vectors
Standard cloning techniques were used to construct all recombinant AAV-based plasmids. Recombinant expression plasmids containing the Rous sarcoma virus (RSV) promoter-driven murine TC-PTP cDNA, and the human PP5 cDNA, were generously provided by M. Tremblay (McGill University, Montreal, PQ, Canada) and D.J. Chen and B.P.C. Chen (University of Texas Southwestern Medical Center at Dallas, Dallas, TX). The scAAV2 plasmids containing the RSV promoter-driven murine TC-PTP (pdsAAV-RSV-TC-PTP) and the RSV promoter-driven human PP5 (pdsAAV- RSV-PP5) were constructed by standard cloning methods as described previously (Zhong et al., 2004a; Zhao et al., 2007). The scAAV plasmid containing the transthyretin (TTR) promoter (pdsAAV-TTR-PP5) driving human PP5 was constructed by a two-step subcloning protocol. The TTR promoter region from pAAV-TTR-Factor VIII was amplified by polymerase chain reaction (PCR), using the primer pair 5′-atac
Highly purified stocks of a recombinant ssAAV2 vector carrying the enhanced green fluorescent protein (EGFP) reporter gene driven by the cytomegalovirus (CMV) promoter (CMVp-EGFP) were obtained from Virapur (San Diego, CA). Highly purified stocks of recombinant ssAAV2 vectors carrying the human factor IX (hF.IX) gene under the control of the apolipoprotein enhancer/human α1-antitrypsin (ApoE/hAAT) promoter (ssAAV2-hF.IX) and scAAV2-PP5, scAAV2-Y730F-PP5, scAAV8-TC-PTP, and scAAV8-PP5 vectors driven by either the RSV or TTR promoter were generated by the triple-plasmid transfection protocol as described previously (Auricchio et al., 2001). The physical particle titers of recombinant vector stocks were determined by quantitative DNA slot-blot analyses (Kube and Srivastava, 1997).
Animal handling
All animal experiments were performed according to the guidelines for animal care specified by the Animal Care Services at the University of Florida (Gainesville, FL). Ten-week-old C57BL/6J male mice were purchased from Jackson Laboratory (Bar Harbor, ME) and maintained at the University of Florida College of Medicine (Gainesville, FL). The Institutional Animal Care and Use Committee approved all protocols for the care and use of these mice.
Recombinant AAV vector transduction studies in vivo
Approximately 5 × 1010 vector genomes (VG) of ssAAV2-EGFP vector alone, or admixed with a high dose (5 × 1010 physical particles per animal, as published previously [Jayandharan et al., 2008]) or a low dose (1 × 109 physical particles per animal) of scAAV2-RSV-PP5, scAAV2-Y730F-RSV-PP5, scAAV2-Y730F-TTR-PP5, scAAV8-RSV-PP5, or scAAV8-TTR-PP5, or 5 × 1010 VG of scAAV8-RSV-TC-PTP and/or scAAV2-PP5 vectors together at a 1:1 ratio, were injected intravenously into C57BL/6J mice via the tail vein (n = 2 per group). Phosphate-buffered saline (PBS)-injected mice were used as an appropriate control. Liver sections from three hepatic lobes of the PBS-injected and vector-injected mice 2 weeks postinjection were evaluated to determine the transduction efficiency of the ssAAV2-EGFP vector. EGFP expression was measured by imaging, using an Axiovert 25 fluorescence microscope (Carl Zeiss, Thornwood, NY). Images from five visual fields of mock- and vector-administered hepatocytes were analyzed quantitatively with ImageJ software (National Institutes of Health, Bethesda, MD). Transgene expression (mean ± SD) was assessed as total area of green fluorescence (pixels squared) per visual field. Analysis of variance (ANOVA) was used to determine the significance of differences between test results and control results.
ssAAV2-hF.IX vectors were injected intravenously (via the tail vein) into C57BL/6J mice (n = 4 per group) at a dose of either 1 × 1010 or 5 × 1010 VG/animal alone or admixed with scAAV8-TTR-PP5 helper virus at a 1:1 ratio at either 1 × 109 or 1 × 1010 VG/animal. Plasma samples were obtained by retro-orbital bleeding and analyzed for hF.IX expression by enzyme-linked immunosorbent assay (ELISA) as described previously (Mingozzi et al., 2003).
Immunohistochemistry
Liver sections from two mice per group were obtained 10 weeks postinjection and analyzed for hF.IX expression. Briefly, liver sections in O.C.T. medium (Sakura Finetek USA, Torrance, CA) were fixed in acetone at room temperature for 10 min, washed, and blocked with 5% donkey serum in PBS for 15 min at room temperature. Goat anti-hF.IX antibody (Affinity Biologicals, Ancaster, ON, Canada) was applied in a 1:200 dilution in 2% donkey serum for 30 min after washing. Samples were washed and incubated with Alexa Fluor 568-conjugated donkey anti-goat IgG (Molecular Probes/Invitrogen, Eugene, OR) diluted 1:200 in 2% bovine serum albumin (BSA) in PBS for 30 min at room temperature. Fluorescence microscopy was performed with a Nikon E800 microscope (Nikon, Tokyo, Japan). Images were captured with a CoolSNAP-Pro camera and analyzed with Image Pro-Plus software (Media Cybernetics, Bethesda, MD). Percent positive cells were estimated by counting stained hepatocytes in 10 frames per mouse and dividing by the average cell number per frame.
Toxicology studies
Liver tissues from PBS-injected mice or those injected with either ssAAV2-EGFP or ssAAV2-hF.IX vectors alone or in combination with scAAV-PP5 helper viruses were collected 2 and 12 weeks postinjection and placed in 10% buffered formalin for fixation and processed for microscopic examination. Gross and microscopic examinations, after hematoxylin and eosin staining, were performed by a pathologist certified by the American College of Veterinary Pathologists affiliated with the University of Florida Pathology Core (Gainesville, FL).
Results
Coadministration with scAAV8-PP5 helper virus leads to maximal transgene expression from ssAAV2-EGFP vectors
We reported that coinjection of scAAV2-TC-PTP and scAAV2-PP5 vectors resulted in an ∼16-fold increase in the transduction efficiency of ssAAV2-EGFP vectors in murine hepatocytes in vivo (Jayandharan et al., 2008). Because AAV8 serotype vectors are known to transduce mouse hepatocytes efficiently (Gao et al., 2002), we wished to evaluate whether the use of scAAV8-TC-PTP and scAAV8-PP5 helper viruses would lead to a further increase in the transduction efficiency of ssAAV2-EGFP vectors. Normal C57BL/6J mice were injected via the tail vein with 5 × 1010 VG of each ssAAV2-EGFP vector alone, or with ssAAV2-EGFP vector combined with scAAV8-RSV-TC-PTP and/or scAAV8-RSV-PP5 vectors. Consistent with previously published studies (Ponnazhagan et al., 1997; Snyder et al., 1997; Nakai et al., 2000; Chen et al., 2001; Song et al., 2001; Zhong et al., 2004a; Zhao et al., 2006; Jayandharan et al., 2008), little green fluorescence was detected in hepatocytes 2 weeks postinjection of conventional ssAAV2-EGFP vectors alone (Fig. 1); however, coinjection with scAAV8-TC-PTP helper virus led to an ∼6-fold increase in the transduction efficiency of ssAAV2-EGFP vectors. Interestingly, scAAV8-PP5 vectors alone led to an ∼24-fold increase in EGFP expression, which was significantly higher than the ∼11-fold increase when scAAV8-TC-PTP and scAAV8-PP5 vectors were coinjected. This was possibly due to competition for receptor usage between the two helper viruses for entry into hepatocytes. It is noteworthy that only the laminin receptor has been described as a cellular receptor for AAV8 (Akache et al., 2006) as opposed to the AAV2 serotype, for which apart from the primary receptor, heparan sulfate proteoglycan (Summerford and Samulski, 1998), at least six different cellular coreceptors are known to exist (Qing et al., 1999; Summerford et al., 1999; Kashiwakura et al., 2005). The difference in helper function between TC-PTP and PP5 may also be due to the fact that in hepatocytes, FKBP52 is phosphorylated predominantly at serine/threonine residues as has been demonstrated previously with AAV2 serotype helper viruses (Jayandharan et al., 2008). Thus, it became clear from these experiments that coadministration of scAAV-PP5 vectors alone could achieve superior helper function, and therefore optimization of scAAV-PP5 helper viruses was envisaged.
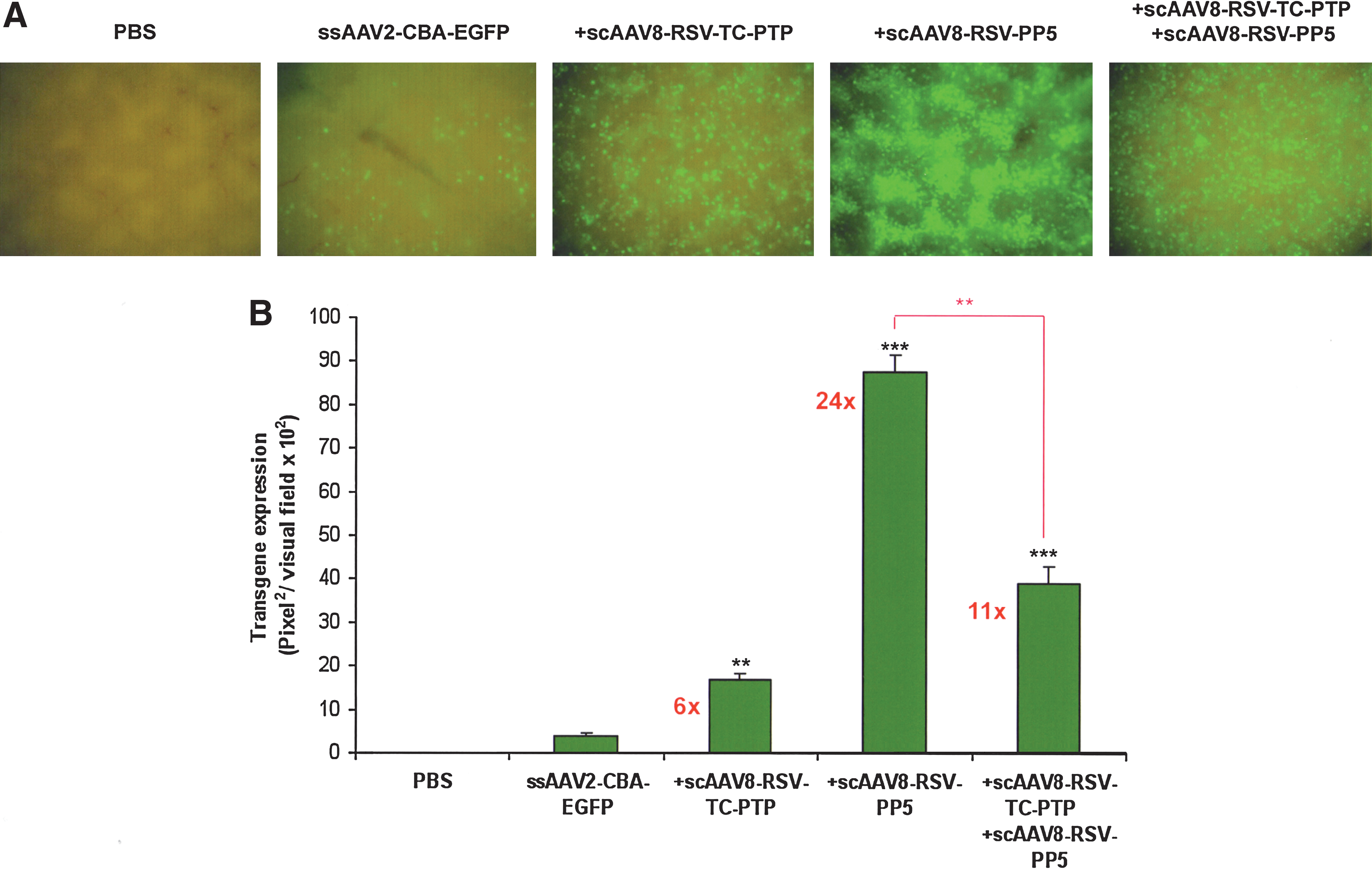
Single-stranded adeno-associated viral (ssAAV) vector-mediated transduction of hepatocytes in normal C57BL/6J mice injected with ssAAV2-EGFP vector alone, or coinjected with either scAAV8-TC-PTP and/or scAAV8-PP5 helper viruses. (
Optimized scAAV2-PP5 helper viruses containing a liver-specific promoter and a tyrosine-mutant capsid enhance transgene expression from ssAAV2-EGFP vectors
We reasoned that the transduction efficiency mediated by the scAAV2-PP5 helper vector could be further augmented by optimizing either the expression cassette or modifications in the AAV2 capsid, or both. We evaluated the TTR promoter, which is hepatocyte-specific, compared with the RSV promoter, expression from which is known to be ubiquitous (Liao et al., 2006). Second, we compared the efficiency of scAAV2-PP5 helper virus with or without capsid modifications in AAV2, because we have shown that a Y730F mutation in the AAV2 capsid significantly enhances transgene expression by bypassing proteasome-mediated degradation (Zhong et al., 2008b). This was evaluated in murine hepatocytes in vivo, at a dose of 5 × 1010 VG each for these optimized helper viruses. The results are shown in Fig. 2. The RSV promoter-driven PP5 helper virus increased the transduction efficiency of ssAAV2-EGFP vectors by ∼9-fold, which is similar to the increase reported previously (Jayandharan et al., 2008). Interestingly, coinjection of the RSV promoter-driven PP5 vector packaged into a Y730F-AAV2 capsid enhanced the transgene expression by ∼17-fold, which was further augmented to ∼22-fold by replacing the RSV promoter with the TTR promoter. These data correlate well with our previous studies that showed the transduction efficiency of Y730F-AAV2 vectors to be ∼29-fold higher than that of the wild-type AAV2 vectors in murine hepatocytes in vivo (Zhong et al., 2008b). These results demonstrate that scAAV2-PP5 helper viruses under the control of TTR promoter and the Y730F capsid mutation can additively augment PP5 expression, which in turn significantly enhances the transduction efficiency of ssAAV2-EGFP vectors.
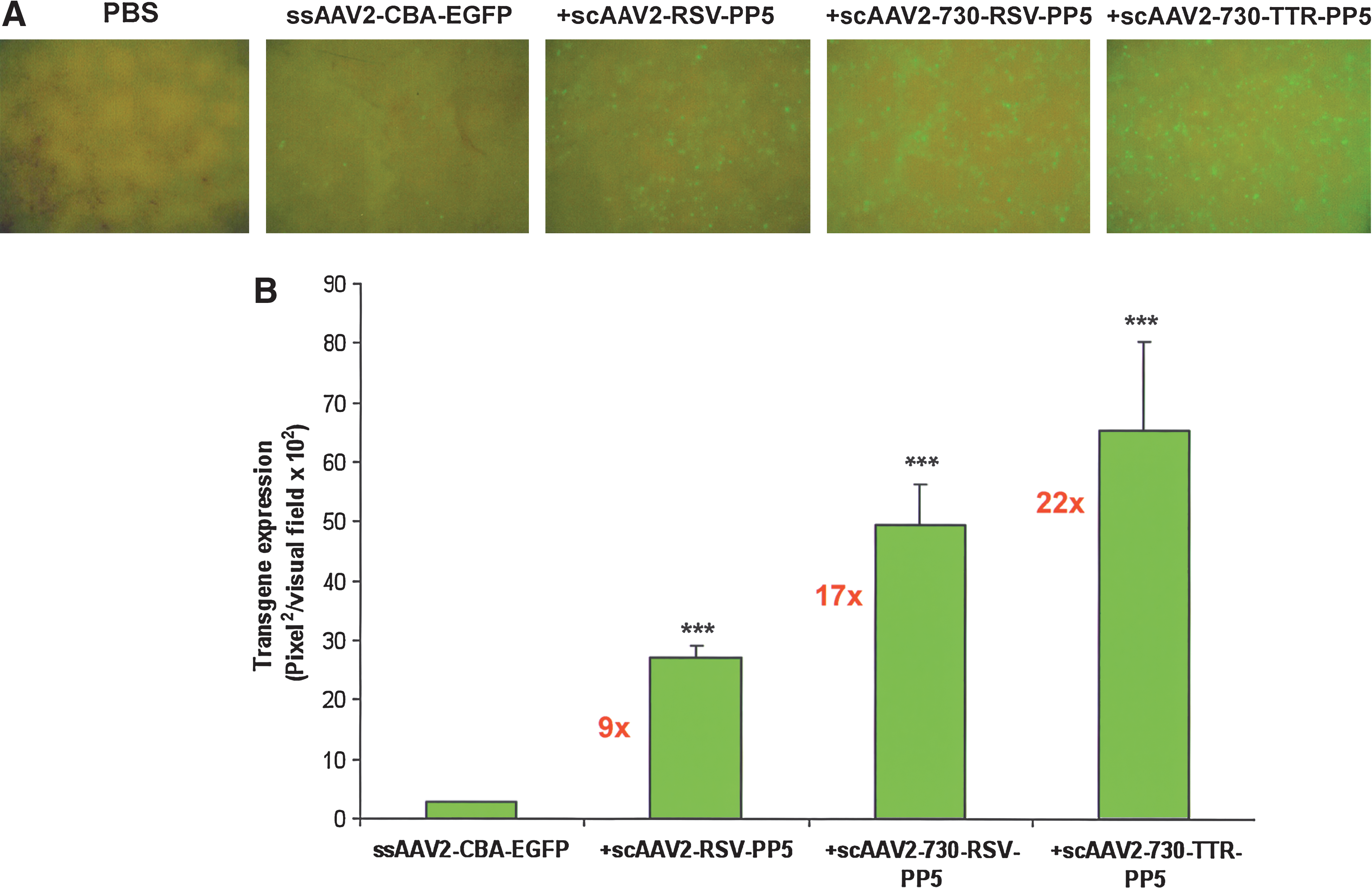
Analysis of self-complementary (sc) AAV2 helper virus-mediated EGFP expression in hepatocytes of normal C57BL/6J mice injected with ssAAV2-EGFP vector alone, or with ssAAV2-EGFP vector coinjected with scAAV2-PP5 helper virus. (
Use of the AAV8 serotype and liver-specific promoter further enhances transgene expression from ssAAV2-EGFP vectors
In anticipation of achieving robust liver-directed expression of PP5 and consequently its helper function, we next examined the potential efficacy of the scAAV-PP5 expression cassette under the control of the TTR promoter packaged into AAV8 serotype capsids. As can be seen in Fig. 3, coinjection with scAAV8-RSV-PP5 helper viruses at 5 × 1010 VG/mouse led to an ∼25-fold increase in the transduction efficiency of ssAAV2-EGFP vectors, which was similar to the ∼24-fold increase seen previously (see Fig. 1). Significantly, we observed an ∼36-fold increase in transgene expression from the ssAAV2-EGFP vectors when scAAV8-PP5 helper virus under the control of the TTR promoter was coinjected at a dose 5 × 1010 VG/mouse, further corroborating our hypothesis that the combination of a liver-specific promoter and the AAV serotype can substantially increase the transduction efficiency of an ssAAV2 vector in murine hepatocytes.
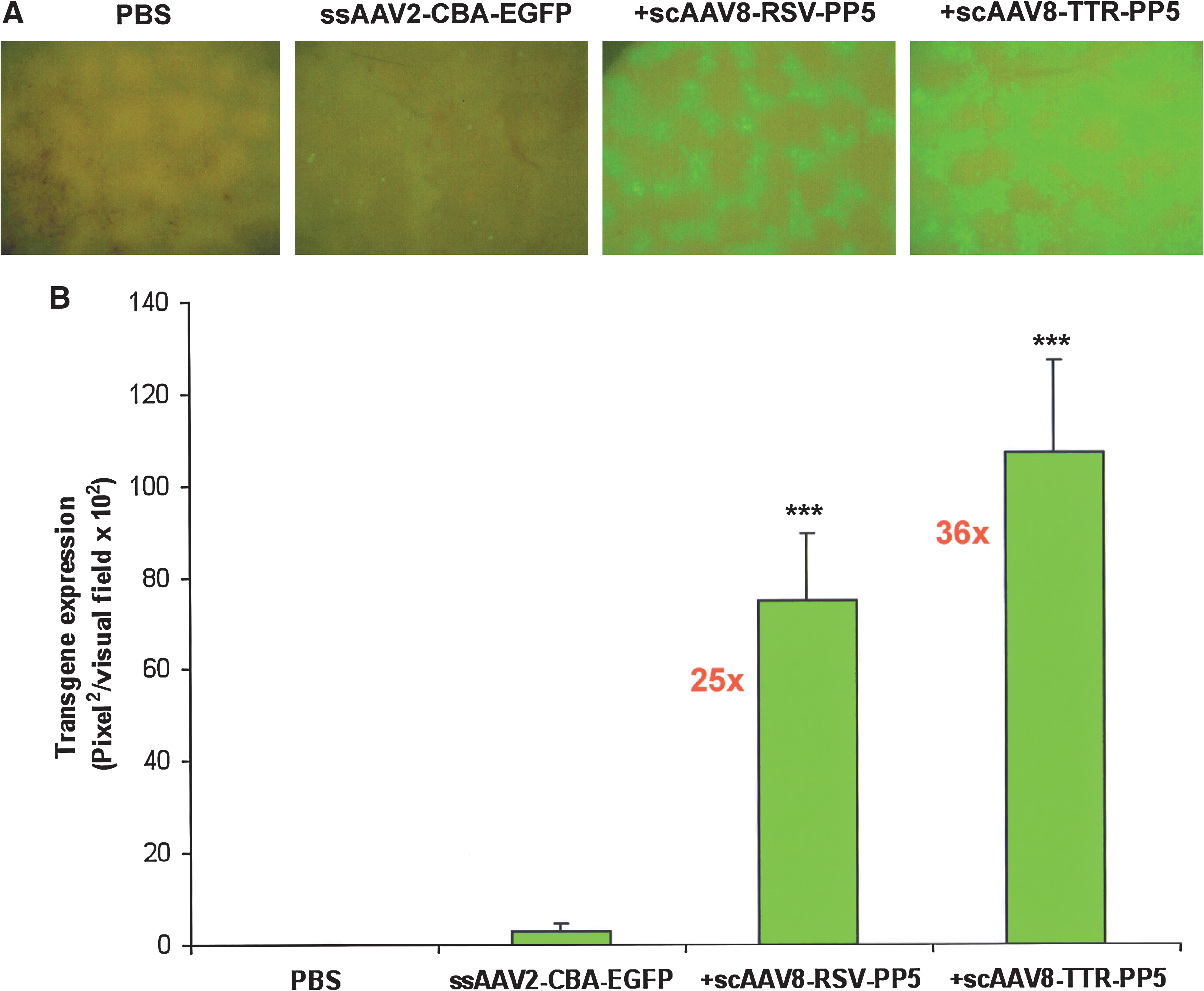
Analysis of scAAV8 helper virus-mediated EGFP expression in hepatocytes of normal C57BL/6J mice injected with ssAAV2-EGFP vector alone, or coinjected with ssAAV2-EGFP vector and scAAV8-PP5 helper virus. (
Reduced dose of helper viruses achieves comparable transgene expression from ssAAV2 vectors
In view of the significant enhancement in the transduction efficiency of ssAAV2-EGFP vectors mediated by scAAV-PP5 helper viruses, it was of interest to examine whether the efficacious helper function could be achieved at reduced helper virus doses. Two different doses of scAAV-PP5 helper viruses, 5 × 1010 (high dose, the original dose; Jayandharan et al., 2008) and 1 × 109 (low dose), were coadministered with 5 × 1010 VG of ssAAV2-EGFP vector. At a 50-fold reduced helper virus dose, the extent of EGFP expression was similar in mice injected with scAAV2-RSV-PP5 vector (∼8- vs. ∼9-fold for the high dose), and only a marginal decrease (statistically insignificant) in transgene expression was seen with scAAV2-Y730F-PP5 capsid mutant helper virus containing the RSV promoter (∼12- vs. ∼17-fold) or the TTR promoter (∼19- vs. ∼22-fold). Similarly, low doses of scAAV8-PP5 helper virus also achieved comparable EGFP expression as in animals that received the high dose (Fig. 4A–C). Taken together, our data demonstrate an incremental effect at both high and low doses for the use of the hepatocyte-specific TTR promoter (∼6- to 7-fold vs. the RSV promoter), capsid-modified AAV2-Y730F vectors (∼6- to 11-fold vs. wild type), and the AAV8 serotype (∼16-fold vs. AAV2) in enhancing transgene expression from an ssAAV2-EGFP vector. It is also clear from these experiments that a maximal ∼19-fold increase in transgene expression mediated by the scAAV2-Y730F-PP5 helper virus, and an ∼30-fold increase with the scAAV8-TTR-PP5 helper virus, can be achieved at a lower dose, thus minimizing any potential risk of toxicity associated with helper virus administration.
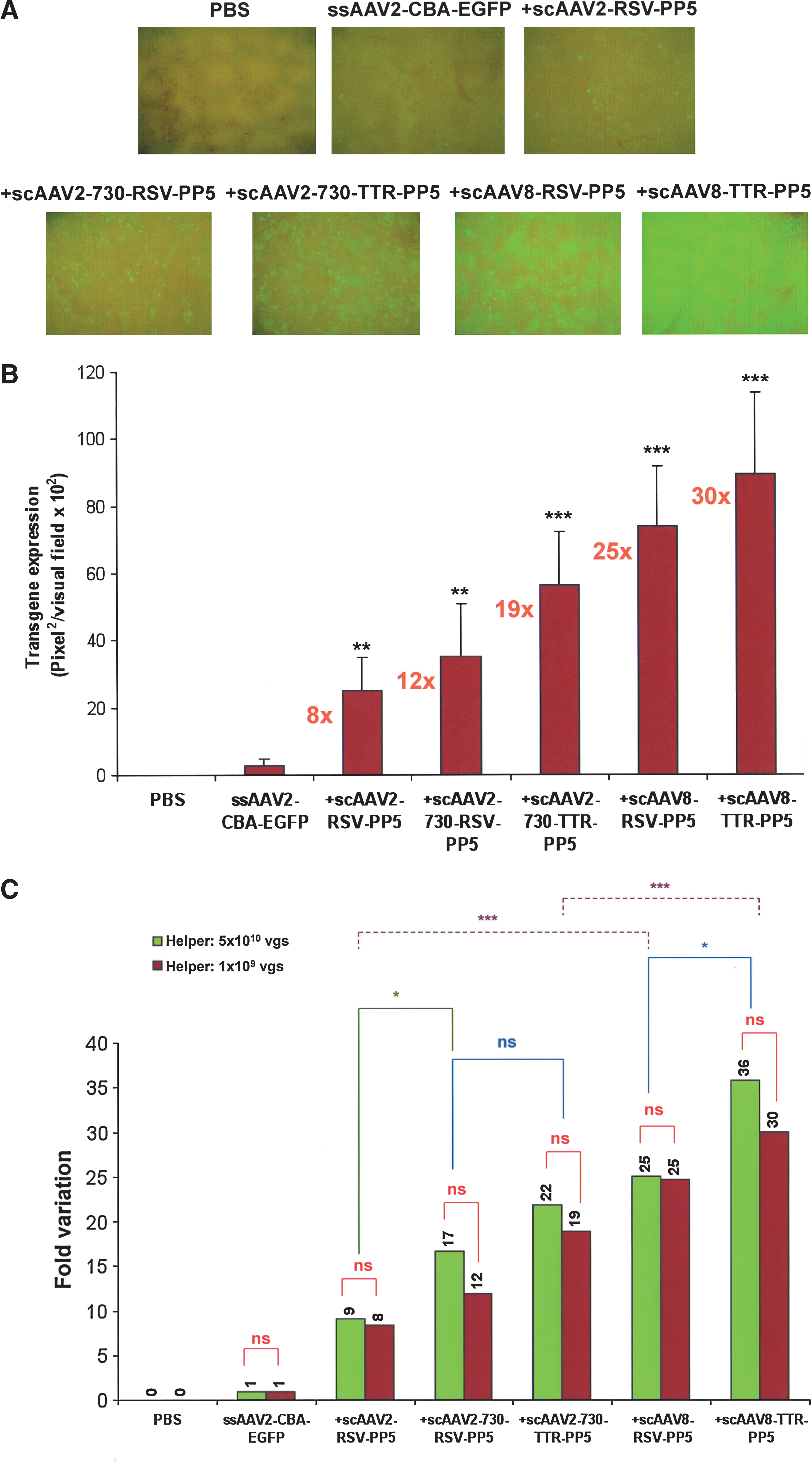
Analysis of scAAV2-PP5 or scAAV8-PP5 helper virus-mediated EGFP expression in hepatocytes of normal C57BL/6J mice injected with a reduced dose of helper viruses. (
Optimized helper viruses allow expression of therapeutic levels of human factor IX at a reduced vector dose in mice
Finally, it was important to examine whether scAAV-PP5 helper viruses were capable of allowing efficient expression of a therapeutic gene at a reduced vector dose in vivo. To this end, 1 × 1010 VG of an ssAAV2-human factor IX (hF.IX) vector alone or coadministered with 1 × 1010 VG of the most efficient helper virus (scAAV8-TTR-PP5) was evaluated in C57BL/6J mice. Over a period of 8 weeks, the PP5 helper virus led to the expression of ∼7-fold higher levels of circulating hF.IX compared with ssAAV2-F.IX vector alone (Fig. 5A). At this dose of ssAAV2-F.IX vector (1 × 1010 VG/mouse), expression was augmented from subtherapeutic to therapeutic levels (216 ± 22 vs. 30 ± 11 ng/ml, 4–5% vs. <1% of normal human F.IX levels). A similar (∼10-fold) enhancement was observed when helper virus at 1 × 109 VG/animal was used (Fig. 5A), resulting in systemic hF.IX expression of 300 ± 90 ng/ml (∼6% of normal) by week 8. A 5-fold higher dose of ssAAV2-F.IX vector alone was required to achieve similar therapeutic levels. When combined with the high-dose ssAAV2-F.IX vector, the helper virus only marginally increased F.IX levels (Fig. 5A).

Comparative analyses of ssAAV2-ApoE/hAAT-hF.IX vector-mediated transduction efficiency in hepatocytes with and without coadministration of scAAV8-PP5 helper viruses in C57BL/6J mice in vivo. (
To investigate whether this enhancement was caused by an increase in hF.IX-producing hepatocytes, liver tissue was collected at 10 weeks from mice receiving a high or low dose of ssAAV2-hAAT-F.IX with or without 1 × 1010 VG of scAAV8-TTR-PP5, and immunofluorescence staining for hF.IX was performed. The percentage of hF.IX-positive hepatocytes in each group is shown in Fig. 5B along with representative images from each group. The results of fluorescence staining correlate with systemic hF.IX expression in that the helper virus had the greatest effect in mice receiving the lower dose of ssAAV2-F.IX, bringing the percent positive hepatocytes from < 1% without helper virus to 3–5% with the helper virus (Fig. 5B). PP5-mediated increases in hF.IX-expressing hepatocytes were less substantial for high-dose ssAAV2-hAAT-F.IX-treated mice (Fig. 5B).
PP5 overexpression is nontoxic in mice
The injection of scAAV-PP5 helper viruses, encapsidated in either AAV2 or AAV8 serotype capsids, did not lead to any detectable histological abnormalities in the liver of C57BL/6J mice at both the low dose (1 × 109 VG/animal) and the high dose (5 × 1010 VG/animal) 2 weeks postinjection, or in mice treated long term (12 weeks) with 1 × 1010 VG of scAAV-PP5 helper viruses. Both PBS- and the helper virus-injected groups of mice were grossly normal. The liver tissues from all PBS- or helper virus-injected animals had no evidence of any toxicity or any pathological lesions, as examined by an experienced pathologist certified by the American College of Veterinary Pathologists. A set of representative data, shown in Fig. 6, corroborate that PP5 overexpression was indeed nontoxic in murine hepatocytes.
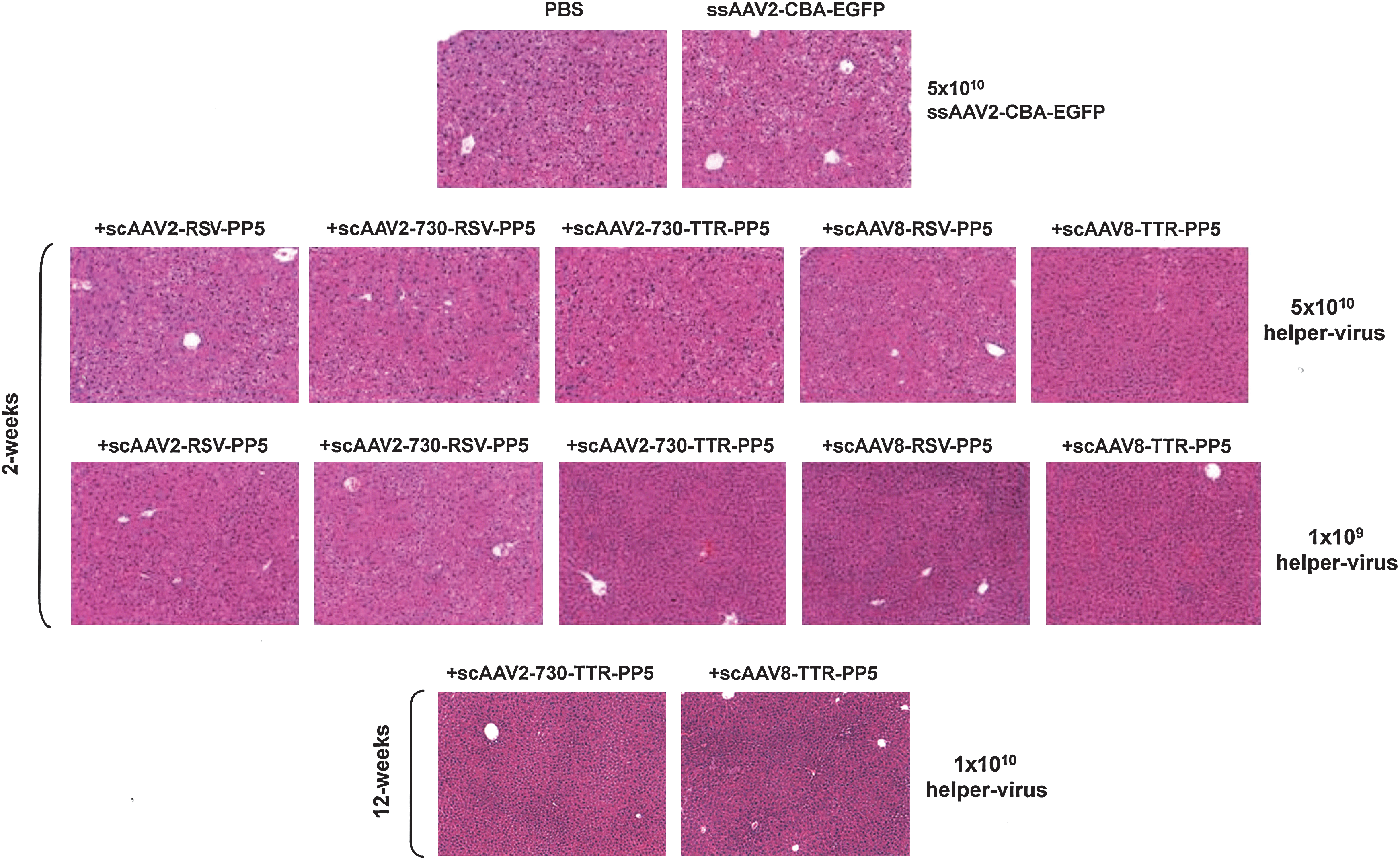
Morphological analysis of murine hepatocytes 2 and 12 weeks postinjection of either PBS or ssAAV2-EGFP at 5 × 1010 VG/animal, and of ssAAV2-EGFP at 5 × 1010 VG/animal coinjected with scAAV-PP5 helper virus (each 5 × 1010, 1 × 1010, or 1 × 109 VG/animal). Liver sections were stained with hematoxylin–eosin (H&E) and visualized under a light microscope. Original magnification, × 400.
Discussion
The efficacious use of recombinant AAV vectors for gene therapy is contingent on a thorough understanding of the fundamental steps in virus–host cell interactions, which include cell surface receptor and coreceptor-mediated viral binding and entry (Summerford and Samulski, 1998; Qing et al., 1999; Summerford et al., 1999), intracellular trafficking (Duan et al., 1999, 2000; Bartlett et al., 2000; Hansen et al., 2000, 2001a), nuclear transport, uncoating (Duan et al., 1999; Bartlett et al., 2000; Sanlioglu et al., 2000; Hansen et al., 2001b; Seisenberger et al., 2001; Thomas et al., 2004), and viral second-strand DNA synthesis (Ferrari et al., 1996; Fisher et al., 1996; Wang et al., 1997; Mah et al., 1998; Qing et al., 1998, 2001). As previously noted, inhibition of viral second-strand synthesis mediated by cellular FKBP52 is an important rate-limiting step that influences directly the efficiency of transgene expression (Ferrari et al., 1996; Qing et al., 1998; Gao et al., 2006). Strategies aimed at reversing the phosphorylation status of FKBP52 by using pharmacologic inhibitors or genetic approaches against epidermal growth factor receptor protein tyrosine kinase (EGFR-PTK), although effective (Mah et al., 1998; Qing et al., 2001, 2003; Zhong et al., 2004b), are not practically feasible for human gene therapy applications because of the associated toxicity and the improbability of generating TC-PTP- and/or PP5-transgenic humans. As an alternative, we have shown in our studies a simple approach to deliver the TC-PTP and the PP5 genes through scAAV vectors, which serve as helper viruses, and significantly improve the efficacy of single-stranded AAV vectors (Zhong et al., 2004a; Jayandharan et al., 2008). To potentially use this coadministration strategy in liver-directed gene therapy, we have carried out further optimizations to achieve therapeutic levels of expression of hF.IX.
In our previous studies the RSV promoter was used, expression from which is ubiquitous (Jayandharan et al., 2008). In the present studies, the use of hepatocyte-specific TTR promoter-driven PP5 expression further enhanced EGFP expression by ∼6- to 7-fold. The liver specificity of the TTR promoter has been reported by several laboratories using a variety of genes (Scallan et al., 2003; Liao et al., 2006; Wu et al., 2008). Apart from increased transduction efficiency, the use of a cell-specific promoter to limit PP5 expression to the liver may be necessary for safe gene therapy applications to reach the correct target and to minimize potential detrimental side effects, if any, caused by PP5 misexpression and overexpression.
Furthermore, given that AAV2 remains the predominant serotype vector currently in use in human gene therapy (Flotte et al., 1996, 2004; Kay et al., 2000; Aitken et al., 2001; Wagner et al., 2002; Manno et al., 2003; Snyder and Francis, 2005), coupled with the fact it is also the best characterized in terms of vector toxicology, we sought to demonstrate the efficacy of the PP5 expression cassette packaged into AAV2 tyrosine-mutant vectors (Zhong et al., 2008b). Compared with wild-type AAV2 capsid vectors, we observed an ∼6- to 11-fold increase in transgene expression with scAAV2-Y730F-PP5 capsid-mutant helper viruses. Tyrosine mutations in scAAV-PP5 helper virus capsids allow escape from proteasome-mediated degradation, and thus efficient vector delivery to the nucleus. Rapid expression of PP5 maintains FKBP52 in a dephosphorylated state in hepatocytes, potentiating transduction from ssAAV vectors. The availability of multiple tyrosine-mutant AAV2 vectors also offers hope for further enhancement of PP5 expression from the next generation of scAAV-PP5 helper viruses at further reduced doses (Zhong et al., 2008b).
Owing to the high efficacy of AAV8-pseudotyped vectors in hepatocytes in vivo, we speculated that scAAV-PP5 genomes packaged into AAV serotype 8 vectors would be more effective for liver-directed gene delivery and expression from an ssAAV2 vector because of the rapid rise and subsequently high levels of expression that can be achieved after systemic administration (Gao et al., 2002; Thomas et al., 2004; Conlon et al., 2005). Second, packaging of the transgene cassette and the helper virus into two different serotype capsids should also reduce any potential competition for cellular receptors on hepatocytes. Indeed, in our study, the AAV8 serotype alone had the most incremental effect of PP5 function on transgene expression (∼16-fold) compared with the AAV2 serotype capsid. Such a function is probably mediated by rapid uncoating of the PP5-carrying AAV8 vector, postinternalization, as has been reported previously (Thomas et al., 2004). This, combined with the use of a hepatocyte-specific TTR promoter, led to ∼36-fold increase in EGFP expression in our present studies. It is also noteworthy that the AAV8 capsid is less immunogenic than the AAV2 capsid (Vandenberghe et al., 2006), and the long-term efficacy and safety of AAV8 vectors have been documented in liver-directed gene therapy in dogs (Wang et al., 2005).
One of the advantages of this helper virus system is that this approach is applicable to any ssAAV transgene cassette, and can be adapted to any of the current clinical protocols without modifications to the therapeutic vector. This could be achieved by a choice of either scAAV2 tyrosine-mutant or scAAV8 capsids, with or without cell-specific promoters for the desired cell tropism. The use of an scAAV8-TTR-PP5 helper virus was the most efficient in enhancing transgene expression from an ssAAV-F.IX vector in the mouse liver. At least ∼6- to 7-fold higher circulating hF.IX levels in a therapeutic range (>200 ng/ml) were detected in helper virus-administered mice compared with those injected with the ssAAV2-F.IX vectors alone. This should enable one to attain the same therapeutic level of expression of a therapeutic gene at a log lower dose, thus minimizing immunological responses against the transgene and the viral capsid. Furthermore, enhancement was possible at a dose of 1 × 109 VG/mouse of scAAV8-TTR-PP5 helper virus, implicating that such a low dose of the helper virus was sufficient to achieve therapeutic levels of expression. This is important in light of evidence suggesting that the level of neutralizing antibody is proportional to the vector dose administered for AAV1 (Petry et al., 2008), and the loss of hepatic hF.IX expression in the hemophilia B clinical trial was due to the CD8+ T cell response to input capsids in patients who received high vector doses (Mingozzi et al., 2007).
The additional optimization steps with the PP5 transgene cassette described here have yielded a further 3-fold increase in transgene expression compared with our previously reported coadministration strategy with scAAV2-TC-PTP and scAAV2-PP5 helper viruses (∼36- vs. ∼11-fold) (Jayandharan et al., 2008). This increase is still substantial and also helps us understand the biological basis of this strategy. Only low levels of PP5 are required for complete dephosphorylation of FKBP52 in hepatocytes, and this in turn pushes transduction through the bottleneck of viral second-strand synthesis, thereby yielding a substantial effect on expression from the therapeutic vector, while additional expression has limited effects.
This paper, for the first time, demonstrates the usefulness of the approach for therapeutic gene expression. A low dose of optimized helper virus was sufficient to achieve therapeutic levels of F.IX expression at otherwise subtherapeutic doses of hF.IX vector, thereby reducing concerns about the safety of PP5 overexpression. The observed difference in transgene enhancement mediated by scAAV8-TTR-PP5 helper virus for EGFP and hF.IX (∼30-fold vs. ∼7- to 10-fold) may be attributable to differences in the transgene product (a cytoplasmic vs. a systemically delivered protein). Clearly, the helper virus was similarly efficacious over a wide dose range, and thus its exact titration will not be required to achieve therapy from any therapeutic ssAAV vector, thereby increasing the probability of achieving the desired effect with a single, uniform dose of the helper virus. For hF.IX expression, the approach was highly effective for a low dose of the therapeutic vector. Less efficient enhancement at the high hF.IX vector dose suggests an additional limiting factor for expression or secretion of this transgene product, but the level of enhancement may in part also have been influenced by the hF.IX expression cassette (which uses a highly optimized liver-specific promoter).
It is noteworthy that PP5 is known to be antiapoptotic, and the development of a viable PP5-deficient mouse model suggests that PP5 is a modulatory, rather than an essential, factor in phosphorylation pathways (Hinds and Sanchez, 2008). This bodes well for the potential use of this strategy in a liver-directed gene therapy application to minimize any vector dose-associated liver toxicity. This is further supported by the fact that toxicological studies performed with FKBP52-dephosphorylating enzymes in normal C57BL/6J mice, with an scAAV-PP5 helper virus, showed no evidence of toxicity up to 12 weeks in the present study, administration of scAAV2-TC-PTP vector did not show any toxicity up to 13 weeks (Zhong et al., 2004a), deliberate overexpression of TC-PTP was not deleterious in TC-PTP-transgenic mice for up to more than 1.5 years of age (Qing et al., 2003; Zhong et al., 2004b), and overexpression of PP5 did not affect cellular growth (Zhao et al., 2007) or hepatocyte characteristics in vivo in C57BL/6 mice (Jayandharan et al., 2008). The development of PP5-transgenic mice, currently underway, is likely to contribute to a better understanding of the underlying mechanism of PP5-mediated enhanced transduction by ssAAV vectors, and will help as well in ascertaining the safety profile of scAAV-PP5 helper viruses.
Footnotes
Acknowledgments
The authors thank Drs. Michel Tremblay, David Chen, and Benjamin Chen for their kind gifts of TC-PTP and PP5 expression plasmids, respectively, and Drs. Xiao Xiao, R. Jude Samulski, and James M. Wilson for generously providing the pdsCBAp-EGFP, pACG2-RC, and pAAV8-RC plasmids, respectively. This research was supported in part by grant 8187368876 from the Roche Foundation for Anemia Research, a research grant from the Fanconi Anemia Research Fund, and by institutional research grant IRG-01-188-04 from the American Cancer Society (to L.Z.), a grant from the St. Baldrick Foundation (to A.E.R.), Public Health Service grants P01 HL-078810 and R01 AI/HL-51390 from the National Institutes of Health (to R.W.H.), and Public Health Service grants R01 EB-002073, R01 HL-065770, HL-076901, and P01 DK-058327 (Project 1) from the National Institutes of Health (to A.S.). G.R.J. was supported in part by an Overseas Associate Fellowship-2006 from the Department of Biotechnology, Ministry of Science and Technology, Government of India.
Author Disclosure Statement
R.W.H. has been receiving royalty payments from Genzyme for license of AAV-F.IX technology.