
Abstract
Lentiviral vectors are promising tools for the treatment of chronic retinal diseases, including age-related macular degeneration (AMD), as they enable stable transgene expression. On the other hand, Sendai virus (SeV) vectors provide the unique advantage of rapid gene transfer. Here we show that novel simian immunodeficiency viral vectors pseudotyped with SeV envelope proteins (SeV-F/HN-SIV) achieved rapid, efficient, and long-lasting gene transfer in the mouse retina. Subretinal exposure to SeV-F/HN-SIV vectors for only a few minutes resulted in high-level gene transfer to the retinal pigment epithelium, whereas several hours were required for gene transfer by standard vesicular stomatitis virus G-pseudotyped SIV vectors. Transgene expression continued over a 1-year period. SeV-F/HN-SIV vector-mediated retinal overexpression of soluble Fms-like tyrosine kinase-1 (sFlt-1) or pigment epithelium-derived factor (PEDF) significantly suppressed laser-induced choroidal neovascularization (CNV). Histologically, 6-month-long sustained overexpression of PEDF did not adversely affect the retina; however, that with sFlt-1 resulted in photoreceptor degeneration associated with choroidal circulation defects. These data demonstrate that brief subretinal administration of SeV-F/HN-SIV vectors may facilitate safe and efficient retinal gene transfer, and suggest the therapeutic potential of PEDF with a higher safety profile for treating CNV in AMD patients.
Introduction
Advances in gene therapy technologies have been introduced as components of novel therapeutic approaches to the treatment of retinal diseases. Among several vector systems, recombinant adeno-associated viral (rAAV) and lentiviral vectors are likely to be useful for patients with chronic progressive retinal diseases, as these vectors can achieve gene transfer in nondividing cells and yield long-term transgene expression (Miyoshi et al., 1997; Acland et al., 2001; Lai et al., 2002; Ikeda et al., 2003). Clinical studies demonstrated the safety and efficacy of the subretinal delivery of rAAV carrying the RPE65 gene for patients with Leber's congenital amaurosis, thereby providing the proof of concept for retinal gene therapy strategies (Bainbridge et al., 2008; Hauswirth et al., 2008; Maguire et al., 2008; Cideciyan et al., 2009). The clinical study has shown that the viral vector solution left in the subretinal space is almost fully absorbed by 24 hr after injection (Bainbridge et al., 2008); however, it is more preferable to remove the vector solution and resolve the retinal detachment (RD) during gene transfer surgery, because the outer retinal cells are deprived of trophic and metabolic support from the retinal pigment epithelium (RPE) and choroid vessels during RD. Hauswirth and colleagues reported that retinal thinning was observed in one of three patients after subretinal injection of rAAV-RPE65 (Hauswirth et al., 2008). We developed novel simian lentiviral vectors pseudotyped with Sendai virus (SeV) hemagglutinin–neuraminidase (HN) and fusion (F) envelope proteins (SeV-F/HN-SIV) (Kobayashi et al., 2003). The F and HN proteins of SeV mediate viral attachment and penetration (Lamb and Kolakofsky, 1996), and SeV-derived vectors have shown efficient gene transfer with only a few minutes of vector–cell interaction (Yonemitsu et al., 2000; Masaki et al., 2001). Therefore, SeV-F/HN-SIV vectors are expected to facilitate rapid transfection, and they may enable the removal of the vector solution shortly after injection.
Neovascularization is thought to be regulated by the balance between angiogenesis inducers and inhibitors (Folkman, 1995). Soluble Fms-like tyrosine kinase-1 (sFlt-1)/VEGF receptor (VEGFR)-1, a splice variant of the VEGF receptor Flt-1, is an endogenous inhibitor of VEGF-A (Kendall et al., 1996), and has been used successfully to attenuate CNV in experimental models (Honda et al., 2000; Bainbridge et al., 2002; Gehlbach et al., 2003). However, studies have indicated that VEGF-A acts as a survival factor in normal vascular endothelial cells and neural cells (Darland et al., 2003; Lee et al., 2007; Maharaj et al., 2008), and some investigators have warned that a long-term blockade of VEGF signaling may lead to vascular and tissue dysfunction in the retina, as observed in the case of other systemic organs (Maynard et al., 2003; Levine et al., 2004; Hurwitz and Saini, 2006). Pigment epithelium-derived factor (PEDF) is a secreted glycoprotein isolated from the conditioned medium of human RPE (Tombran-Tink and Johnson, 1989; Steele et al., 1993), and it has been shown to exhibit both antiangiogenic and neuroprotective properties (Taniwaki et al., 1995; Dawson et al., 1999). Retinal gene transfer of PEDF by adenoviral vectors substantially inhibited experimental CNV by inducing apoptosis in activated endothelial cells (Mori et al., 2002); and this approach is currently under evaluation in a clinical trial investigating its efficacy in AMD patients (Campochiaro et al., 2006). However, the effects of long-term PEDF overexpression on the normal retinal vasculature and neurons remain unknown.
In the present study, we aimed to evaluate the availability of SeV-F/HN-SIV vectors for retinal gene transfer. We demonstrated the rapid and efficient transduction ability of these vectors, both in vitro and in vivo. Using this new vector system, we assessed the efficacy and safety of long-term overexpression of the antiangiogenic agents sFlt-1 and PEDF in a mouse model of laser-induced CNV.
Material and Methods
SIV vectors
To produce third-generation recombinant SIV-based lentiviral vectors, HEK 293T cells were transfected with the packaging vector; gene transfer vectors encoding enhanced green fluorescent protein (EGFP), hPEDF, or hsFlt-1 driven by the cytomegalovirus promoter; the ReV expression vector; and the envelope vector (vesicular stomatitis virus G glycoprotein [pVSV-G] [Clontech Laboratories, Mountain View, CA] or pSeV-F/HN with a truncation of the cytoplasmic tail of F and the addition of the cytoplasmic tail of the SIV transmembrane envelope protein to the N terminus of HN) (Kobayashi et al., 2003). The SIV vector lacking a transgene cassette (SeV-F/HN-SIV-Empty) was used as the control vector. The U3 region in the 3′ and 5′ long terminal repeat of SIV was deleted to induce self-inactivation. The viral titer was determined by the transduction of HEK 293T cells and expressed as transducing units (TU) per milliliter, and the viruses were kept at −80°C until just before use.
Cell culture and in vitro gene transfer
ARPE-19 cells, a human RPE-derived cell line, were purchased from the American Type Culture Collection (Manassas, VA). ARPE-19 cells were seeded in 24-well plates at 1 × 105 cells per well in serum-free Dulbecco's modified Eagle's medium (DMEM)–F12. Twelve hours later, either SeV-F/HN-SIV-luciferase or VSV-G-SIV-luciferase was added to each well at a multiplicity of infection (MOI) of 10. After various times of incubation with each vector solution, the culture medium was removed, the cells were washed twice with phosphate-buffered saline (PBS), and fresh medium was added. At 48 hr after gene transfer, the cells were harvested and subjected to luciferase assay.
Animals and in vivo gene transfer
Adult C57BL/6 mice were maintained humanely, with proper institutional approval, and in accordance with the statement of the Association for Research in Vision and Ophthalmology. All animal experiments were carried out according to approved protocols and in accordance with the recommendations for the proper care and use of laboratory animals by the Committee for Animals, Recombinant DNA, and Infectious Pathogen Experiments at Kyushu University and according to the Law (No. 105) and Notification (No. 6) of the Japanese Government. The subretinal injection of each solution was performed as previously described (Murakami et al., 2008b). Briefly, mice were anesthetized by inhalation of ether. A 30-gauge needle was inserted into the subretinal space of the peripheral retina in the nasal hemisphere via an external transscleral transchoroidal approach. Subretinal injection of vector solution (SeV-F/HN-SIV or VSV-G-SIV vector; 2.5 × 107 TU/ml × 2 μl) resulted in a dome-shaped detachment of about half the retina. To assess the effects of vector–cell interaction time on retinal gene transfer, we used two different subretinal administration techniques: either simple injection (leave) or removal 5 min after gene transfer by draining the subretinal vector solution and injecting balanced salt solution (BSS) after drainage (remove). The following assessments for duration of transgene expression, the efficacy of antiangiogenic factors for laser-induced CNV, and their retinal toxicity were performed by the remove procedure. Eyes that sustained marked surgical trauma (e.g., retinal or subretinal hemorrhage, bacterial infection) were excluded from further analyses.
Luciferase assay
Procedures used for the luciferase assay have been described previously (Ikeda et al., 2002). ARPE-19 cells and enucleated mouse eyes were treated with 1 × lysis buffer (Promega, Madison, WI) with a protease inhibitor cocktail and centrifuged, and then 20-μl samples of the supernatants were mixed with 100 μl of luciferase assay buffer. Light intensity was measured with a luminometer (model LB 9507; Berthold Technologies, Bad Wildbad, Germany) with 10-sec integration.
Detection of GFP expression in vivo and indocyanine green angiography
GFP expression in the mouse retina was examined with a scanning laser ophthalmoscope (Heidelberg retinal angiograph; Heidelberg Engineering, Heidelberg, Germany), using a 488-nm excitation laser light and a 500-nm barrier filter. Indocyanine green (ICG) angiography was performed 7 min after intraperitoneal injection of 0.2 ml of 10% ICG, using a 795-nm excitation diode laser light and an 810-nm barrier filter, as previously described (Janssen et al., 2008).
Histological examination
Mouse eyes were enucleated, and both paraffin and cryosections were prepared. To prepare the paraffin sections, the eyes were fixed with 4% paraformaldehyde in PBS for 24 hr, and were then mounted in paraffin. For the cryosections, the eyes were frozen in liquid nitrogen, and 5-μm-thick sections were prepared along the horizontal meridian. The sections were subsequently stained with hematoxylin and eosin. The number of cells in the outer nuclear layer was counted per 250 μm at six points around the retinal section (A1–A3, from the ora serrata to the optic nerve of the temporal hemisphere; A4–A6, from the optic nerve to the ora serrata of the nasal hemisphere).
Immunohistochemistry
Five-micrometer-thick cryosections were cut, air dried, and fixed in cold acetone for 10 min. The sections were blocked with 3% nonfat dried milk and labeled with rabbit anti-GFP polyclonal antibody (diluted 1:300; Invitrogen Molecular Probes, Eugene, OR) at 4°C for 24 hr. After biotinylated goat anti-rabbit IgG (H+L) (diluted 1:200; Vector Laboratories, Burlingame, CA) was applied as a secondary antibody, the cells were incubated with fluorescein isothiocyanate (FITC)-conjugated streptavidin (diluted 1:100; BD Biosciences, San Jose, CA). After labeling, 4′,6-diamidino-2-phenylindole (DAPI) was used to counterstain the nuclei. Immunofluorescence images were acquired with an Olympus BX51 microscope with a fluorescence attachment (Olympus, Tokyo, Japan). For negative controls, the primary antibody was omitted.
Enzyme-linked immunosorbent assay
The protein content of mouse eyes was determined with an enzyme-linked immunosorbent assay (ELISA) kit for human PEDF (not available for mouse PEDF; Chemicon International, Temecula, CA) and human sFlt-1 (not available for mouse sFlt-1; R&D Systems, Minneapolis, MN). For the preparation of ocular tissue, conjunctival and muscular tissues were removed from enucleated eyes. The eyes were washed with PBS, minced with scissors in 500 μl of 1 × lysis buffer with a protease inhibitor cocktail, and centrifuged at 15,000 rpm for 5 min at 4°C. The supernatants were subjected to ELISA according to the manufacturer's instructions.
Laser-induced CNV
Laser photocoagulation (630 nm, 150 mW, 0.1 sec, 75 μm) was performed on one eye (four spots per eye) of each animal, using a slit-lamp delivery system (NIDEK, Aichi, Japan), as previously described (Tsutsumi et al., 2003). Burns were performed at the 2, 5, 7, and 10 o'clock positions around the optic disk such that two laser-treated sites were within the vector-injected area, and the other two laser-treated sites were located outside of that area. Laser injury disrupted the RPE, Bruch's membrane, and choroid, and induced the subsequent proliferation and migration of choroidal endothelial cells, resulting in CNV. On day 14 after the laser induction of injury, the eyes were enucleated and fixed with 4% paraformaldehyde. The eyecups, obtained by removing the anterior segments and the entire neural retina, were incubated with FITC-conjugated isolectin B4 (Vector Laboratories) at 4°C for 24 hr. Then, CNV was visualized by Olympus BX51 fluorescence microscopy, and the area was measured with National Institutes of Health (Bethesda, MD) ImageJ software.
Transmission electron microscopy
The eyes were enucleated, and the posterior segments were fixed in 2.5% glutaraldehyde and 2% paraformaldehyde in 0.1 M cacodylate buffer with 0.08 M CaCl2 at 4°C. Sections of retina, RPE, and choroid complex in the vector-injected area were postfixed for 1.5 hr in 2% aqueous OsO4, dehydrated in ethanol and water, and embedded in Epon. Ultrathin sections were cut from blocks and stained with saturated, aqueous uranyl acetate and Sato's lead stain. The specimens were observed with a Hitachi H-7650 electron microscope (Hitachi High-Technologies, Tokyo, Japan).
Statistical analyses
All values are expressed as means ± SD. Statistical differences were assessed by analysis of variance (ANOVA) followed by Tukey–Kramer adjustments for multiple comparisons. Numbers per group are as indicated. A p value of less than 0.05 was considered statistically significant.
Results
SeV-F/HN-SIV vectors require only a brief vector–cell interaction to achieve efficient gene transfer
We previously demonstrated that recombinant Sendai viral (rSeV) vectors exhibited efficient gene transfer into the RPE of the rat retina with a brief exposure time (Ikeda et al., 2002). To determine whether SeV F- and HN-pseudotyped SIV vectors would be capable of rapid and efficient transfection, we first investigated the vector–cell interaction time-dependent transgene expression level. In a human RPE-derived cell line, ARPE-19, conventional VSV-G-pseudotyped SIV (VSV-G-SIV) vectors encoding luciferase showed an interaction time-dependent increase in luciferase expression, and more than 24 hr of exposure was required to achieve maximal gene transfer (Fig. 1A). In contrast, with SeV-F/HN-SIV-luciferase, only 1 min of exposure yielded efficient luciferase expression at levels similar to those observed with 48 hr of exposure (Fig. 1B). Moreover, the luciferase expression levels achieved with SeV-F/HN-SIV vectors were more than 10-fold higher than the peak value obtained with VSV-G-SIV vectors (Fig. 1A and B).
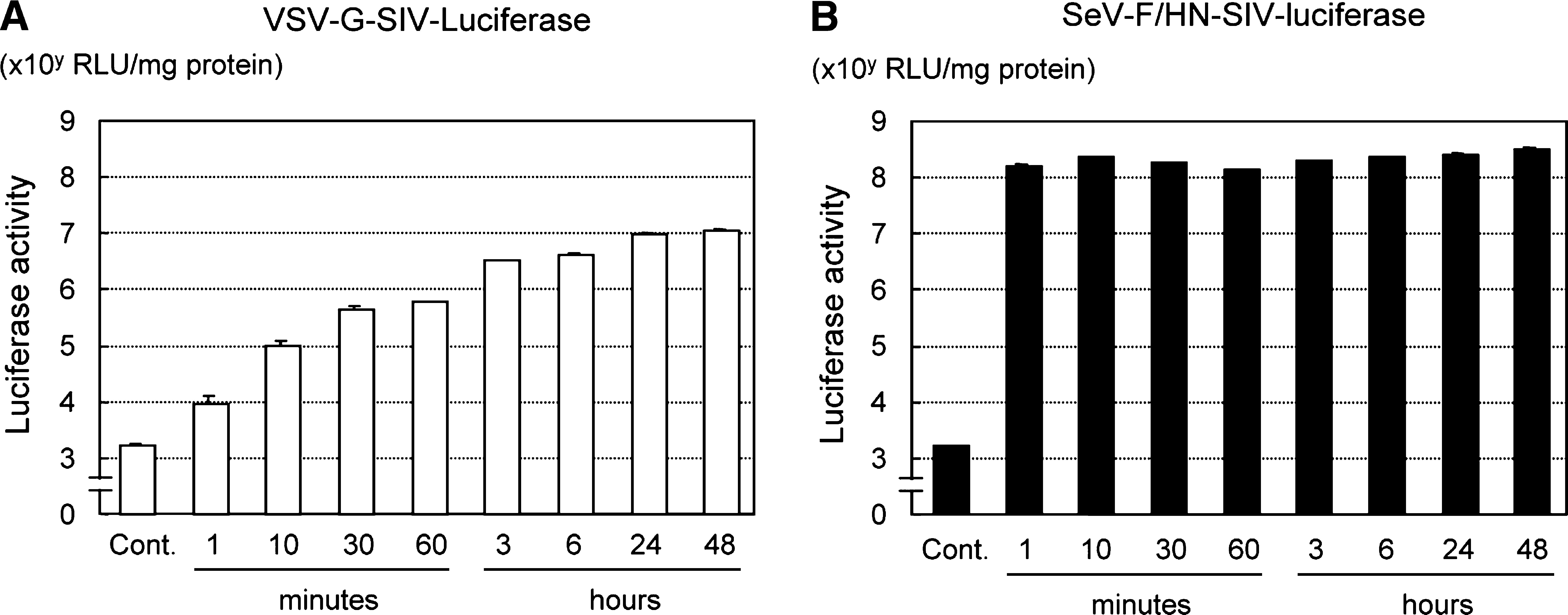
Effect of vector–cell interaction time on gene transfer to ARPE-19 cells. (
Next, we assessed the in vivo transduction efficiency of SeV-F/HN-SIV vectors in the mouse retina. To assess the effects of vector–cell interaction time on retinal gene transfer, we subretinally injected the SIV vectors by one of two different techniques, that is, either the vector solution was left in the subretinal space (leave), or the solution was removed 5 min after vector injection (remove). In eyes treated with VSV-G-SIV-luciferase, transgene expression in the remove group was markedly reduced, by approximately one-tenth, in comparison with that of the leave group (Fig. 2A). In contrast, eyes treated with SeV-F/HN-SIV-luciferase showed efficient transgene expression even in the remove group, and the luciferase expression levels in the SeV/F/HN-SIV-vector remove group were 1.3-fold higher than those of the VSV-G-SIV-vector leave group (Fig. 2A). These data indicate that brief vector–cell contact is sufficient for SeV-F/HN-SIV vectors to achieve efficient retinal gene transfer.
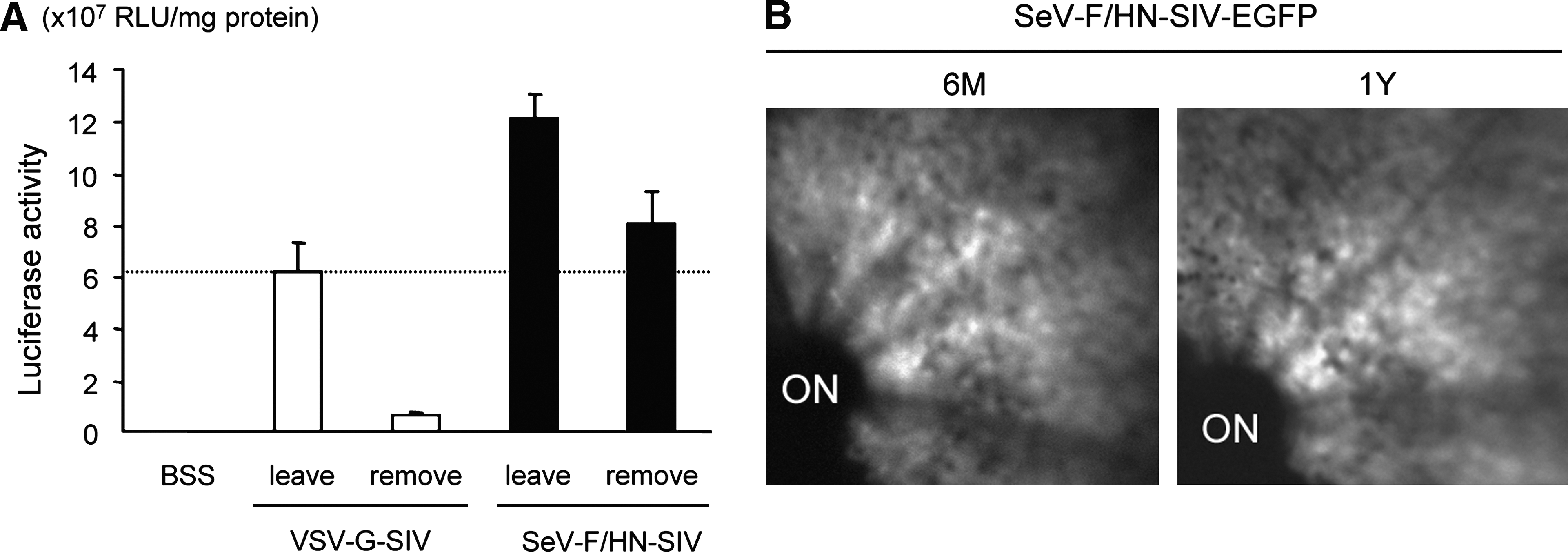
SeV-F/HN-SIV vector-mediated retinal gene transfer with brief exposure time. (

Stable long-term transgene expression in RPE by SeV-F/HN-SIV vectors
Our previous study demonstrated that subretinal injection of VSV-G-SIV vector at 2.5 × 107 TU/ml resulted in sustained transgene expression over a 1-year period in the rat retina (Ikeda et al., 2003). To determine the longevity of SeV-F/HN-SIV vector-mediated retinal gene transfer, we monitored the time course of transgene expression using GFP as a reporter. Mouse retinas treated with SeV-F/HN-SIV-EGFP by the remove procedure showed intense GFP fluorescence in an area corresponding to the vector-injected area, and the extent of GFP fluorescence was maintained for at least 1 year (Fig. 2B). Histological examination revealed that GFP expression was located in the RPE layer (Fig. 2C), as previously observed in the case of rSeV-mediated retinal gene transfer (Ikeda et al., 2002; Murakami et al., 2008b). Taken together, these findings indicate that SeV-F/HN-SIV vectors exhibit the advantageous features of both SeV vectors and SIV vectors, that is, rapid transduction and long-term transgene expression ability, in retinal tissue.
SeV-F/HN-SIV vector-mediated retinal delivery of antiangiogenic factors suppresses laser-induced CNV
Next, we sought to investigate the effects of SeV-F/HN-SIV vector-mediated PEDF or sFlt-1 gene transfer on laser-induced CNV. Mouse eyes were treated with SeV-F/HN-SIV-Empty, -hPEDF or -hsFlt-1 at 2.5 × 107 TU/ml by the remove procedure. ELISA revealed that sufficient levels of hPEDF (790.3 ± 96.1 ng/g protein) and hsFlt-1 (47.6 ± 10.0 ng/g protein) were obtained 2 weeks after vector injection, and were sustained for at least 3 months postinjection (Fig. 3A and B). We performed laser photocoagulation 2 weeks after vector injection, and assessed the area of CNV 2 weeks after the laser treatment. CNV size was significantly reduced with SeV-F/HN-SIV vector-mediated retinal delivery of hPEDF or hsFlt-1, compared with CNV size in the eyes untreated or treated with SeV-F/HN-SIV-Empty (p < 0.01; Fig. 3C and D). Treatment with SeV-F/HN-SIV-hPEDF or hsFlt-1 also showed potent suppression of CNV when laser photocoagulation was performed 3 months after vector injection (p < 0.01; Fig. 3F and G), suggesting potential long-term inhibitory effects on CNV. Next, we compared CNV size within and outside of the vector-injected area. Treatment with SeV-F/HN-SIV-hPEDF or -hsFlt-1 was associated with significant CNV suppression in both areas (p < 0.05; Fig. 3E and H).
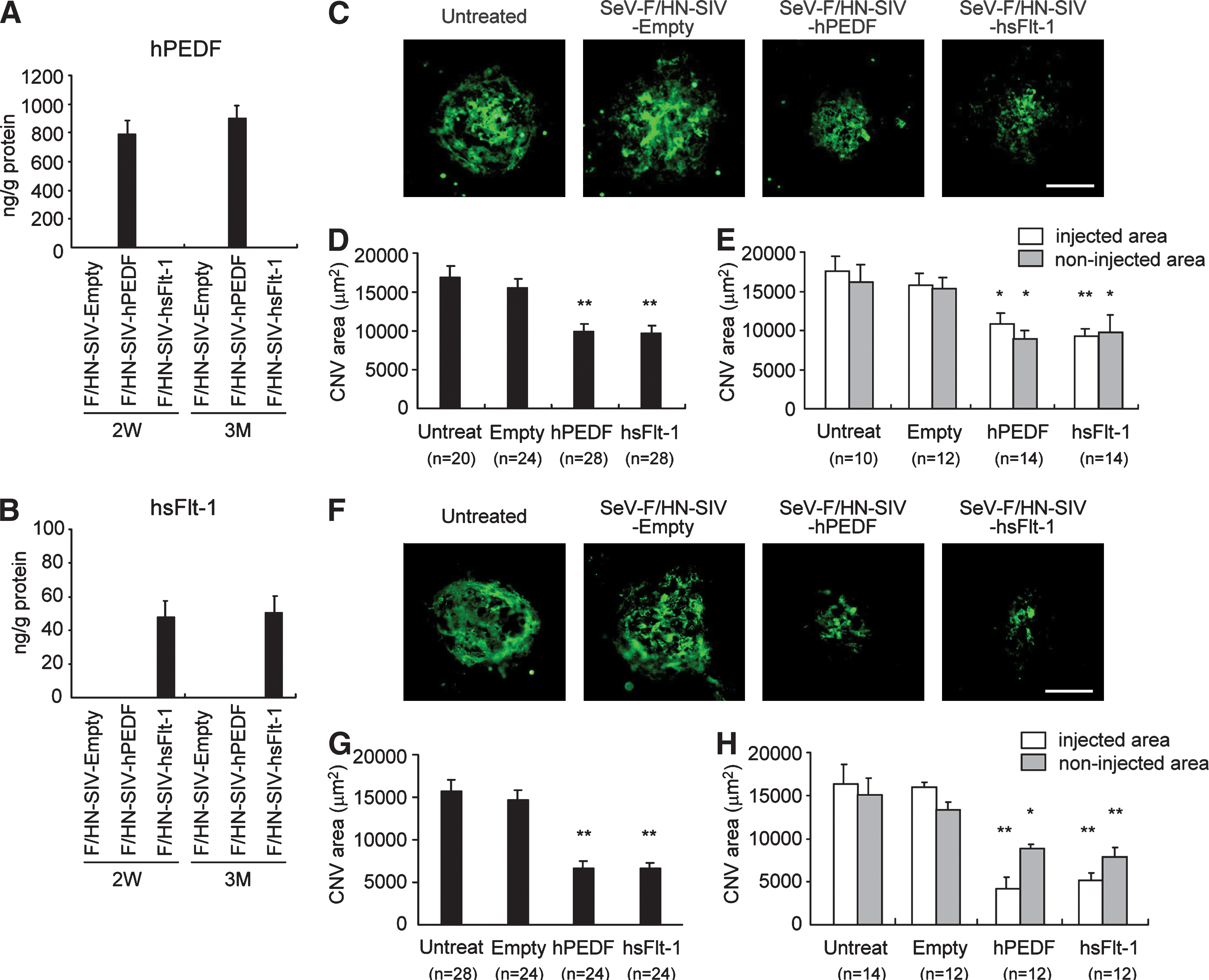
Suppression of laser-induced choroidal neovascularization (CNV) by subretinal injection of SeV-F/HN-SIV-hPEDF or SeV-F/HN-SIV-hsFlt-1. (
Assessment of retinal toxicity with long-term overexpression of antiangiogenic factors
To determine any potential adverse effects of long-term overexpression of antiangiogenic factors on the retina, we next performed a histological examination of SeV-F/HN-SIV-treated retinas 6 months after vector injection by the remove procedure. Eyes that had undergone 6 months of sustained overexpression of hPEDF showed no significant changes in retinal structure; however, 6 months of sustained overexpression of hsFlt-1 resulted in a significant loss of photoreceptors in the vector-injected area (p < 0.05; Fig. 4A and B). These findings suggest that long-term blockade of VEGF signaling may be deleterious for maintaining retinal homeostasis in adult mice.

Assessment of potential retinal toxicity resulting from gene transfer of PEDF or sFlt-1 in normal adult mice. (
In the adult mouse retina, VEGF-A is secreted basally from the RPE, and fetal liver kinase-1 (Flk-1)/VEGFR-2 is expressed on the endothelium of the choriocapillaris facing the RPE layer, which is suggestive of a paracrine interaction between the two tissues (Saint-Geniez et al., 2006). To analyze the changes in choroidal vasculature observed with long-term overexpression of sFlt-1, we performed ICG angiography by in vivo scanning laser ophthalmoscopy. Retinas treated with SeV-F/HN-SIV-hPEDF or -Empty showed no significant alteration of ICG fluorescence, compared with that of untreated retinas. In contrast, fluorescein-filling defects of choroidal vessels were observed in those retinas treated with SeV-F/HN-SIV-hsFlt-1 in the area corresponding to the vector-injected site (Fig. 5). Ultrastructural analysis by transmission electron microscopy revealed that there were choriocapillaris vessels filled with densely packed, malformed erythrocytes with adjoining thrombocytes 1 month after SeV-F/HN-SIV-hsFlt-1 treatment (Fig. 6A and B). These findings were not observed in retinas untreated or treated with SeV-F/HN-SIV-Empty or -hPEDF (Fig. 6C and data not shown). No alteration of choriocapillaris endothelial cell fenestrations was observed between the groups (Fig. 6D). Ultrastructural signs of apoptosis of endothelial cells or RPE cells were not evident (data not shown). These findings indicate that long-term retinal delivery of PEDF is a safe and effective approach to suppressing CNV, but that long-term sFlt-1 expression may disturb the homeostatic balance of the retina, resulting in a disruption of the choroidal circulation and photoreceptor degeneration.
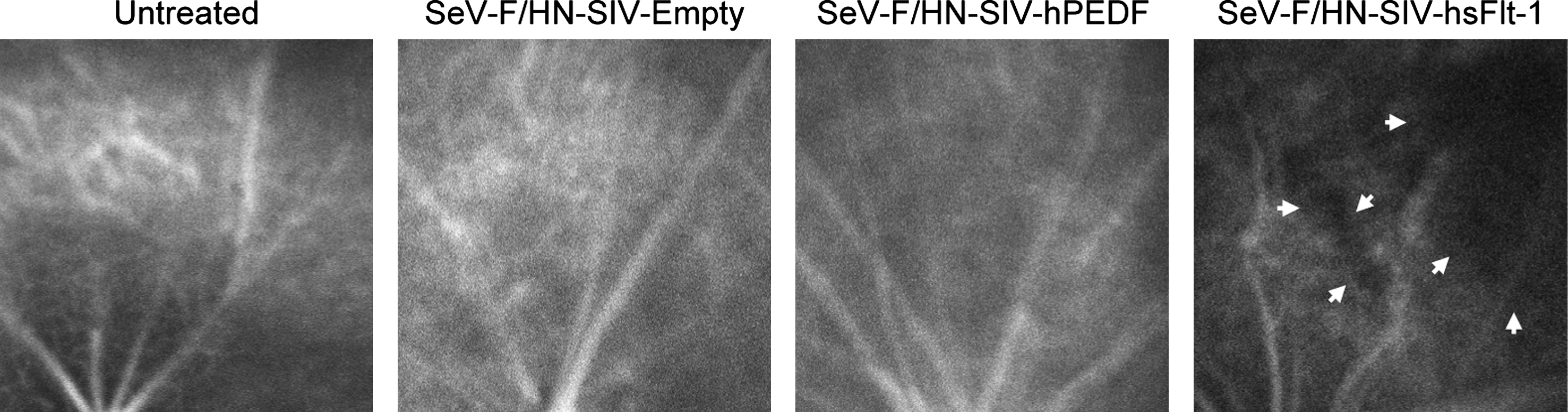
In vivo imaging of choroidal vessels by indocyanine green (ICG) angiography. Shown are ICG angiograms of the vector-injected area of mouse retinas 6 months after treatment with SeV-F/HN-SIV-Empty, SeV-F/HN-SIV-hPEDF, or SeV-F/HN-SIV-hsFlt-1 (n = 4 each). Eyes of untreated mice were used as a control. Arrows indicate defects in choroidal circulation in a retina treated with SeV-F/HN-SIV-hsFlt-1.

Ultrastructural analysis of the choriocapillaris by transmission electron microscopy. (
Discussion
In the present study, we characterized novel SIV-based lentiviral vectors pseudotyped with SeV-F and SeV-HN for retinal gene transfer. The key observations made in this study are as follows: (1) a brief vector–cell interaction period was sufficient for the SeV-F/HN-SIV vectors to achieve efficient gene transfer into RPE cells; (2) transgene expression mediated by SeV-F/HN-SIV vectors was stable and sustained over a 1-year period; (3) SeV-F/HN-SIV vector-mediated retinal gene transfer of hPEDF or hsFlt-1 substantially suppressed experimental CNV in mice; and (4) long-term overexpression of hPEDF did not exert any significant deleterious effects on the retinal tissue, whereas the long-term overexpression of hsFlt-1 resulted in photoreceptor degeneration in association with choroidal circulation defects. The rapid transduction ability of SeV-F/HN-SIV vectors is in clear contrast to reported findings obtained with conventional VSV-G-pseudotyped lentiviral vectors, rAAV, or adenoviral vectors (Teramoto et al., 1998; Masaki et al., 2001). To the best of our knowledge, this is the first report to demonstrate a limitation of the use of sFlt-1 for retinal gene therapy.
In this study, we demonstrated that the conventional VSV-G-pseudotyped SIV vectors required more than 24 hr of interaction time to achieve maximal gene transfer. rAAV and adenoviral vectors, which have been used in clinical trials of gene therapy for retinal diseases, also showed an interaction time-dependent increase in transgene expression and required more than 12 hr to reach the maximal expression level (Maeda et al., 1998; Teramoto et al., 1998). In contrast, the novel SIV vectors pseudotyped with SeV-F and SeV-HN achieved high-level gene transfer within several minutes, both in vitro and in vivo, as seen in previous reports using rSeV vectors (Ikeda et al., 2002). This unique feature of SeV-F/HN-SIV vectors enables the removal of subretinal vector solution and the resolution of RD during gene transfer surgery. Although subretinal injection of rAAV-RPE65 at the doses used in clinical studies has been shown to be safe in dogs and nonhuman primates (Jacobson et al., 2006a,b), retinal thinning was observed after subretinal injection of rAAV-RPE65 in one of three patients (Hauswirth et al., 2008). Of note, optical coherence tomography showed that the thickness of the outer nuclear layer was especially reduced in this patient. Retinal neurons, which are chronically stressed by degenerative retinopathy, may be more vulnerable to environmental changes. It is premature to assess the complications associated with subretinal viral vector injection, due to the small number of patients treated; however, we believe that the remove technique may reduce the nutrient starvation in outer retinal cells caused by RD and may provide safer retinal gene transfer. In addition, SeV-F/HN-SIV vectors exhibited stable and long-term transgene expression in the RPE over a 1-year period. In the clinical setting, most cases of AMD progress over several years, and long-lasting therapeutic effects are required for treatment. One limitation of the currently available anti-VEGF drugs is their short half-life in the eye and the need for repeated injections (Bakri et al., 2007). Therefore, SeV-F/HN-SIV vector-mediated continuous delivery of therapeutic proteins would be an attractive approach for the long-term inhibition of CNV in AMD patients.
Clinical studies have demonstrated the beneficial therapeutic effects of intravitreal injection of anti-VEGF-A drugs such as pegaptanib (an aptamer specific for VEGF-A165) and ranibizumab (an Fab fragment of a humanized monoclonal pan-VEGF-A antibody) (Gragoudas et al., 2004; Brown et al., 2006; Rosenfeld et al., 2006). However, some investigators have raised concerns about the safety of long-term intraocular VEGF-A neutralization (Sang and D'Amore, 2008). In this study, we delivered the sFlt-1 gene into the RPE of the mouse retina via subretinal injection of SeV-F/HN-SIV-hsFlt-1, and we demonstrated that 6-month sustained overexpression of sFlt-1 resulted in photoreceptor degeneration. Although we previously demonstrated that subretinal injection of high-titer VSV-G-SIV vector (2.5 × 108 TU/ml) induced sustained inflammation around the RPE and caused retinal degeneration (Ikeda et al., 2003), such inflammatory reaction and retinal damage were not observed in retinas treated with SeV-F/HN-SIV-Empty or -hPEDF at 2.5 × 107 TU/ml, or in retinas treated with VSV-G-SIV vector at this dose (Ikeda et al., 2003), suggesting that overexpression of sFlt-1 is a major mediator of photoreceptor degeneration after SeV-F/HN-sFlt-1 treatment. In line with our data, Saint-Geniez and colleagues demonstrated that systemic VEGF neutralization by adenoviral vector-mediated overexpression of sFlt-1 induced the apoptosis of retinal neuronal cells, including photoreceptors (Saint-Geniez et al., 2008). However, there are several contradictory reports on the retinal effect of sFlt-1. Lai and colleagues and Pechan and colleagues demonstrated the long-term safety of rAAV vector-mediated retinal gene transfer of sFlt-1 (Lai et al., 2005; Pechan et al., 2009). One reason for these differences might involve differences in the levels of sFlt expression, because hsFlt-1 protein levels achieved with rAAV vectors were substantially lower than those observed in our study and in the study by Saint-Geniez and colleagues. In another study, Ueno and colleagues generated transgenic mice with doxycycline-inducible expression of sFlt-1 in their photoreceptors, and no significant changes in retinal structure or function were observed after a 7-month period of doxycycline treatment (Ueno et al., 2008). The reasons for these discrepant results remain unclear, but differences between cells expressing sFlt-1 may account for such discrepancies. Because VEGF-A is secreted basally from the RPE (Blaauwgeers et al., 1999), sFlt-1 expressed by the RPE may more effectively sequester VEGF-A and disrupt the homeostatic balance of the RPE–choriocapillary complex. These retinal effects of sFlt-1 are not necessarily identical to those of anti-VEGF-A drugs, because sFlt-1 binds not only to VEGF-A, but also to other VEGF family members such as VEGF-B, which exerts a potent neuroprotective effect on retinal neurons (Takahashi and Shibuya, 2005; Li et al., 2008). However, long-term follow-up of patients administered treatment with anti-VEGF-A drugs is required to monitor the eyes for retinal toxicity, as intravitreal injection of bevacizumab is known to cause mitochondrial swelling and disruption of the cristae in the photoreceptors of rabbit eyes (Inan et al., 2007).
The mechanisms of photoreceptor loss after VEGF neutralization remain unknown. One possible explanation would be that sFlt-1 may block the neuroprotective signaling of VEGF-A in photoreceptors. Nishijima and colleagues reported that VEGF-A directly protected retinal neurons in the ganglion cell layer and inner nuclear layer after ischemic reperfusion injury via the activation of Flk-1 (Nishijima et al., 2007). However, it remains unclear whether or not VEGF-A also provides direct neuroprotection to photoreceptors, because the levels of expression of VEGF receptors in photoreceptors were significantly lower than those in inner retinal neurons (Stitt et al., 1998; Nishijima et al., 2007; Li et al., 2008). As an alternative possibility, the impairment of choroidal circulation, which provides vascular support for photoreceptors and RPE cells, might induce photoreceptor degeneration. Flk-1 is strongly expressed in the endothelium of the choriocapillaris facing the RPE layer in the adult mouse retina (Saint-Geniez et al., 2006). In this study, we showed that persistent expression of sFlt-1 in the RPE resulted in a disruption of choroidal circulation, which suggests that VEGF signaling plays an essential role in maintaining the choroidal circulation. This explanation may also be supported by the finding that conditional inactivation of VEGF-A expression in the RPE layer resulted in an absence of choroidal vessels, disorganization of photoreceptors, and loss of visual function (Marneros et al., 2005). In ultrastructural analysis, we found choriocapillaris vessels filled with packed erythrocytes and thrombocytes after sFlt-1 gene transfer. These findings were similar to those made after intravitreal bevacizumab treatment in nonhuman primates (Peters et al., 2007), and suggest that these obstructions of the choriocapillaris might be a cause leading to choroidal circulation defects. No alteration of choriocapillaris endothelial cell fenestrations was observed in this study. Although Peters and colleagues reported early reduction of fenestrations after bevacizumab treatment, the reduction was recovered transiently (Peters et al., 2007). These changes in fenestrations were not observed after sustained systemic or local overexpression of sFlt-1 (Saint-Geniez et al., 2008; Ueno et al., 2008), suggesting that the loss of fenestrations might be a transient effect and would not be detectable at later time points.
Retinal gene transfer of PEDF by SeV-F/HN-SIV vectors efficiently inhibited experimental CNV at a level equivalent to that seen with SeV-F/HN-SIV-hsFlt-1, and did not exert any toxic effects on the normal retinal tissue. It has been demonstrated that PEDF induces the apoptosis of stimulated endothelial cells, but not that of quiescent endothelial cells, by targeting Fas/CD95 and the nuclear factor of activated T cells upregulated or activated by VEGF-A (Volpert et al., 2002; Zaichuk et al., 2004). Using the same mechanism, PEDF also induces the apoptosis of endothelial cells stimulated by other angiogenic factors such as basic fibroblast growth factor. Taken together, our findings and those of previous reports indicate that PEDF may specifically target activated endothelial cells during neovascularization, without affecting mature existing vessels. In addition to its antiangiogenic effects, PEDF has a neuroprotective effect on retinal neuronal cells (Miyazaki et al., 2003; Takita et al., 2003). We reported that PEDF directly inhibited photoreceptor apoptosis by regulating the mitochondrial release of apoptosis-inducing factor via Bcl-2 upregulation (Murakami et al., 2008a). These results suggest that gene therapy strategies using PEDF, which possesses both antiangiogenic and neuroprotective abilities, may be safe and effective for the treatment of retinal neovascular and degenerative diseases.
In conclusion, we have demonstrated that novel SIV vectors pseudotyped with SeV-F and SeV-HN showed rapid and efficient gene transfer to the RPE; thus, this system would enable the removal of subretinal vector solution shortly after vector injection. The long-term retinal delivery of PEDF by SeV-F/HN-SIV vectors was safe and effective at suppressing experimental CNV, whereas long-term delivery of sFlt-1 led to a disruption of the choroidal circulation and photoreceptor degeneration. These findings indicate that retinal gene therapy using PEDF may be a useful therapeutic strategy for the long-term management of CNV in AMD patients with a higher safety profile.
Footnotes
Acknowledgments
This work was supported in part by the Japanese Ministry of Education, Culture, Sports, Science, and Technology (grants-in-aid 20791259, 18390115, and 19791277 to Y.I., Y.Y., and M.M.). The authors are grateful to Drs. Eiji Akiba, Katsuyuki Mitomo, and Toshiaki Tabata for excellent technical assistance with vector construction and large-scale production. The authors also thank Mr. Hiroshi Fujii and Miss Chie Arimatsu for assistance with the experiments.
Author Disclosure Statement
Yoshikazu Yonemitsu is a member of the Scientific Advisory Board of the DNAVEC Corporation.