
Abstract
The rhodopsin gene (RHO) encodes a highly expressed G protein-coupled receptor that is central to visual transduction in rod photoreceptors. A suite of recombinant 2/5 adeno-associated viral (AAV) RHO replacement vectors has been generated in an attempt to recapitulate endogenous rhodopsin levels from exogenously delivered AAV vectors in the retina of mice with a targeted disruption in the rhodopsin gene (Rho –/– mice). Approximately 40% of wild-type mouse rhodopsin mRNA levels (RNA taken from whole retinas) was achieved in vivo in AAV-RHO-injected eyes, representing approximately 50-fold increases in expression compared with the initial vector. The main focus of this study was to test whether expression of AAV-RHO replacement in Rho –/– mice provided therapeutic benefit, which to date had not been achieved. Rho –/– mice neither elaborate rod outer segments nor have rod-derived electroretinograms (ERGs). Our results indicate for the first time in this model that subretinal AAV-RHO delivery leads not only to RHO immunolabeling but the generation of rod outer segments as evaluated by light and transmission electron microscopy. Improved histology was accompanied by rod photoreceptor activity as assessed by ERG for at least 12 weeks postinjection. The most efficient AAV-RHO constructs presented in this study provide sufficient levels of RHO to be of therapeutic benefit in Rho –/– mice and therefore represent important steps toward generating potent AAV-RHO replacement genes for gene therapy in RHO-linked human retinopathies.
Introduction
The rhodopsin gene is highly expressed in rod photoreceptors, with the encoded transmembrane protein constituting approximately 90% of the total protein content of mammalian rod outer segment (ROS) disk membranes. Rhodopsin traverses the disk membranes at seven domains, typifying the structure of GPCRs. On average each ROS in the murine retina contains approximately 900 disks, which are continually replaced at a rate of approximately 10% per day (Palczewski, 2006; Lodowski et al., 2009). The development of mice with disruptions of the endogenous mouse rhodopsin (Rho) gene (Rho –/– mice) (Humphries et al., 1997; Lem et al., 1999) has provided a valuable tool for further exploration of rhodopsin. Homozygous Rho –/– mice do not elaborate ROS, have no recordable rod photoreceptor electroretinogram (ERG), and lose photoreceptors over approximately a 3- to 4-month period postnatally; a time frame of loss modulated somewhat by genetic background (Hobson et al., 2000).
A repertoire of recombinant viral vectors including lenti-, adeno-, and adeno-associated viral (AAV) vectors mediates in vivo gene delivery to photoreceptor cells and can be used to explore gene therapies in animal models (Lebherz et al., 2008; Li et al., 2008; Surace and Auricchio, 2008; Stieger et al., 2009). In more recent studies AAV serotypes including AAV2/5 and AAV2/8 have been predominantly used for gene delivery to photoreceptors because of efficient transduction, relatively low toxicity, and the long-term gene expression that can be achieved (Lebherz et al., 2008; Li et al., 2008; Surace and Auricchio, 2008; Stieger et al., 2009). The profile of AAV for retinal gene therapy has been further enhanced by results from phase 1 human clinical trials supporting the safety of AAV2/2 in the human eye (Bainbridge et al., 2008; Cideciyan et al., 2008; Maguire et al., 2008).
Mutant rhodopsins have been implicated in a variety of inherited eye disorders (Farrar et al., 2002) including congenital stationary night blindness, and autosomal dominant and autosomal recessive retinitis pigmentosa (RP) (
Knowledge of the underlying genetic pathogenesis, together with the availability of animal models (Olsson et al., 1992; Li et al., 1996; Humphries et al., 1997) and vectors for delivery, has enabled exploration of gene-based therapies for inherited retinopathies. Many of these studies have focused on treatment of recessively inherited degenerations. For example, AAV2/2-mediated delivery of the RPE-65 gene to Briard dogs presenting with RPE-65-linked Leber congenital amaurosis (LCA) has been shown to restore vision (Acland et al., 2001). At present, four human clinical trials for RPE-65-associated LCA are either under way (Bainbridge et al., 2008; Cideciyan et al., 2008; Maguire et al., 2008) or scheduled to commence in 2010 (Le Meur et al., 2007). Therapeutic benefit in the recessive AIPL1 mouse model of LCA has also been demonstrated (Tan et al., 2009).
In contrast to recessive disease, progress in developing gene therapies for dominant retinal disorders such as RHO-linked adRP has been slower for a multiplicity of reasons. Because significant mutational heterogeneity is associated with RHO-linked adRP (
Materials and Methods
Recombinant adeno-associated virus preparation
Eight recombinant AAV2/5 (AAV) viruses were generated, using a helper virus-free system as described (O'Reilly et al., 2007). Briefly, expression cassettes were cloned into pAAV-MCS (Stratagene, La Jolla, CA), between the inverted terminal repeats of AAV2. The resulting constructs were transfected into human embryonic kidney (HEK)-293 cells (accession no. CRL-1573; American Type Culture Collection [ATCC], Manassas, VA) with pRep2/Cap521 and pHelper (Stratagene), at a ratio of 1:1:2. Fifty 150-mm plates of confluent cells were transfected, using polyethylenimine. Forty-eight hours posttransfection, crude viral lysates were cleared and purified by CsCl2 gradient centrifugation. AAV-containing fractions were dialyzed against phosphate-buffered saline (PBS). Genomic titers (DNase-resistant viral particles per milliliter; VP/ml) were determined by quantitative real-time reverse transcription-polymerase chain reaction (qRT-PCR). AAV-Rho-P-EGFP and AAV-CMV-P-EGFP contained a 1.7-kb mouse Rho promoter (Rho-P) or CMV-P-driven EGFP (Fig. 1). A CMV-P sequence, β-globin intron, and HGH polyadenylation signal from pAAV-MCS (accession no. AF396260.1; Stratagene) were used in AAV constructs (Fig. 1). Constructs also contained a selection of other sequences such as an EGFP coding sequence (accession no. U55761), CMV enhancer (CMVE; accession no. EF550208, nucleotides 308–734), and minimal synthetic poly(A) (Levitt et al., 1989) and WPRE sequences (accession no. J04514; Fig. 1). A replacement human rhodopsin cDNA (RHO) sequence (RHO-BB) was constructed by modifying the wild-type human RHO sequence (accession no. NM_000539.2) at nucleotides 254–274 as follows: 5′-AT

Schematic representation of AAV vector constructs. EGFP was expressed using either the CMV immediate-early promoter (CMV-P) or the 1.7-kb fragment of the rhodopsin promoter (1.7 Rho-P). Replacement rhodopsin-BB (RHO-BB) was expressed with a 1.7- or 0.5-kb fragment of Rho-P, whereas shBB RNA was expressed from the H1 promoter (H1shBB). The RHO-BB replacement and the H1shBB expression cassettes were separated by a spacer fragment (spacer) in suppression and replacement constructs. The effects of various elements on RHO-BB expression were tested in these constructs. β-G int, human β-globin intron; HGH pA, human growth hormone polyadenylation signal; CMVE, CMV enhancer; 3′UTR, 3′ untranslated region of RHO mRNA, including polyadenylation signals; WPRE, woodchuck hepatitis posttranscriptional regulatory element; E, conserved region E of Rho-P (Supplementary Table 1); B, conserved region B of Rho (Supplementary Table 1); m pA, minimal polyadenylation signal. Restriction enzyme sites used for cloning are given. Numbers indicate molecular sizes in base pairs, and arrows indicate the direction of transcription. Color images available online at
Subretinal AAV injection, RNA isolation, and analysis
Subretinal injections were performed in strict compliance with the European Communities Regulations 2002 and 2005 (Cruelty to Animals Act) and the Association for Research in Vision and Ophthalmology statement for the use of animals in ophthalmic and vision research as described (O'Reilly et al., 2007). Briefly, adult mice were anesthetized by intraperitoneal injection of medetomidine and ketamine (10 and 750 mg/10 g body weight, respectively). Pupils were dilated with 1% cyclopentolate and 2.5% phenylephrine, and, using topical anesthesia (amethocaine), a small puncture was made in the sclera. A 34-gauge blunt-ended microneedle attached to a 10-μl syringe (Hamilton) was inserted through the puncture, AAV in PBS was administered to the subretinal space, and retinal detachment was induced. After subretinal injection, a reversing agent (atipamezole hydrochloride, 100 mg/10 g body weight) was delivered by intraperitoneal injection. Body temperature was maintained with a homeothermic heating device. Newborn (P0) mice were prepared for subretinal injection by the method described by Matsuda and Cepko (2004). Typically, 2 μl of AAV-RHO (5 × 1012 VP/ml [1 × 1010 VP] in PBS) were administered to adult wild-type 129 mice whereas 0.5 μl of AAV-RHO (5 × 1012 VP/ml [2.5 × 109 VP] or 1.3 × 1013 VP/ml [6.5 × 109 VP]) was administered to P0 Rho –/– mice. Two microliters of AAV-Rho-P-EGFP or AAV-CMV-P-EGFP (1 × 1012 VP/ml [2 × 109 VP]) were injected into adult wild-type 129 mice. RNA was isolated 10 days or 2 weeks postinjection and analyzed by RNase protection or qRT-PCR as described (O'Reilly et al., 2007).
Morphologic analyses by light and transmission electron microscopy
Rhodopsin immunocytochemistry and fluorescence microscopy were performed as described (Kiang et al., 2005) 2, 6, or 12 weeks postinjection. For RHO-specific immunocytochemistry, 3A6 primary rhodopsin antibody was used at a 1:10 dilution. For tissue preparation for transmission electron microscopy (TEM) a method previously described (Wolfrum, 1992) was slightly modified. Briefly, 6 weeks postinjection, AAV-transduced eyes were enucleated, fixed in 4% paraformaldehyde in PBS, and whole mounted. Using the EGFP tracer, EGFP-positive and -negative areas from the central part of the retinas were excised and fixed in 2.5% glutaraldehyde in 0.1 M cacodylate buffer (pH 7.3) for 2 hr at room temperature. Specimens were washed and fixed in buffered 2% OsO4, dehydrated, and embedded in Araldite. Semithin (0.5 μm) and ultrathin (60 nm) sections were cut with a Leica Ultracut S microtome (Leica Microsystems). For light microscopy semithin sections were analyzed with a Leica DM 6000 B microscope. Ultrastructural analyses were performed with a Tecnai 12 Bio-TWIN transmission electron microscope (FEI, Eindhoven, The Netherlands) and imaged with an SIS MegaView III surface channel charge-coupled device (SCCD) camera (Olympus Soft Imaging Solutions, Münster, Germany). ROS measurements were performed with analySIS software (Olympus Soft Imaging Solutions).
Electroretinography
Rod-isolated ERG recordings were prepared as described (Chadderton et al., 2009). Briefly, animals were dark-adapted overnight and all procedures were carried out under dim red light. For anesthesia, ketamine and xylazine (16 and 1.6 mg/10 g body weight, respectively) were injected intraperitoneally. Pupils were dilated with 1% cyclopentolate and 2.5% phenylephrine. Eyes were maintained in a proptosed position throughout the examination. Reference and ground electrodes were positioned subcutaneously, approximately 1 mm from the temporal canthus and anterior to the tail, respectively. The ERG responses were recorded simultaneously from both eyes by means of gold wire ring electrodes (Goldring; Roland Consult, Brandenburg, Germany), which were positioned to touch the central cornea of each eye. Corneal hydration and electrical contact were maintained throughout the examination by the application of a small drop of Vidisic (Dr. Mann Pharma, Berlin, Germany) to the cornea. Standardized flashes of light were presented to the mouse in a Ganzfeld bowl. Responses were analyzed with a RETI-scan/RETI-port electrophysiology unit (Roland Consult). The protocol used was based on that approved by the International Clinical Standards Committee for human ERG. Rod-isolated responses were recorded with a dim white flash (–25 dB maximal intensity, where maximal flash intensity was 3 candelas/m2/sec) presented in the dark-adapted state. In accordance with the standard convention, a-waves were measured from the baseline to the trough and b-waves from the baseline. After 10 min of light adaptation (30 candelas/m2) cone responses were recorded to the standard flash presented at 0.5- and 10-Hz flicker against the rod-suppressing background.
Statistical analysis
Means and standard deviation (SD) values of data sets were calculated. Statistical significance of differences between groups was determined by analysis of variance (ANOVA), using the least significant difference (LSD) post hoc test (Data Desk 6.1; Data Description, Ithaca, NY); differences with p < 0.05 were considered statistically significant.
Results
The primary focus of the current study was to optimize RHO expression from AAV vectors and to evaluate these vectors in vivo in Rho –/– mice. A comparative evaluation of a series of RHO constructs was undertaken in an attempt to optimize AAV-delivered RHO replacement expression in mouse retinas (Fig. 1).
The CMV promoter (CMV-P) incorporated into AAV vectors has previously been shown to drive high levels of transgene expression in a wide variety of cell types (Lebherz et al., 2008; Li et al., 2008; Mueller and Flotte, 2008; Surace and Auricchio, 2008; Stieger et al., 2009). CMV-P-driven and 1.7-kb Rho-P-driven EGFP expression vectors containing the human β-globin intron and the human growth hormone polyadenylation signal were constructed (AAV-CMV-P-EGFP and AAV-Rho-P-EGFP; Fig. 1). Subsequent to subretinal injection of adult wild-type mice with AAV-CMV-P-EGFP or AAV-Rho-P-EGFP at 2 × 109 viral particles (VP)/eye, EGFP expression was analyzed by histology, fluorescence-activated cell sorting (FACS), and qRT-PCR 2 weeks postinjection. Histological analysis revealed strong EGFP expression in photoreceptor cell bodies and segments for both promoters (Fig. 2A and C); no expression was observed in other components of the retina. Retinas were then dissociated and analyzed by FACS (Fig. 2B and D) to compare intensities of EGFP expression. Higher intensities of EGFP fluorescence were found in cells from retinas transduced with AAV-Rho-P-EGFP versus AAV-CMV-P-EGFP (Fig. 2B and D). Subsequently, the sorted cells were used for qRT-PCR analysis (Fig. 2E), which revealed an approximately 5-fold (p < 0.001, n = 3) higher mRNA level in cells from retinas transduced with AAV-Rho-P-EGFP. The higher level of expression achieved with Rho-P, compared with CMV-P, stimulated additional engineering of the retina-specific Rho-P to augment expression levels, rather than further exploration of a general promoter such as CMV-P.
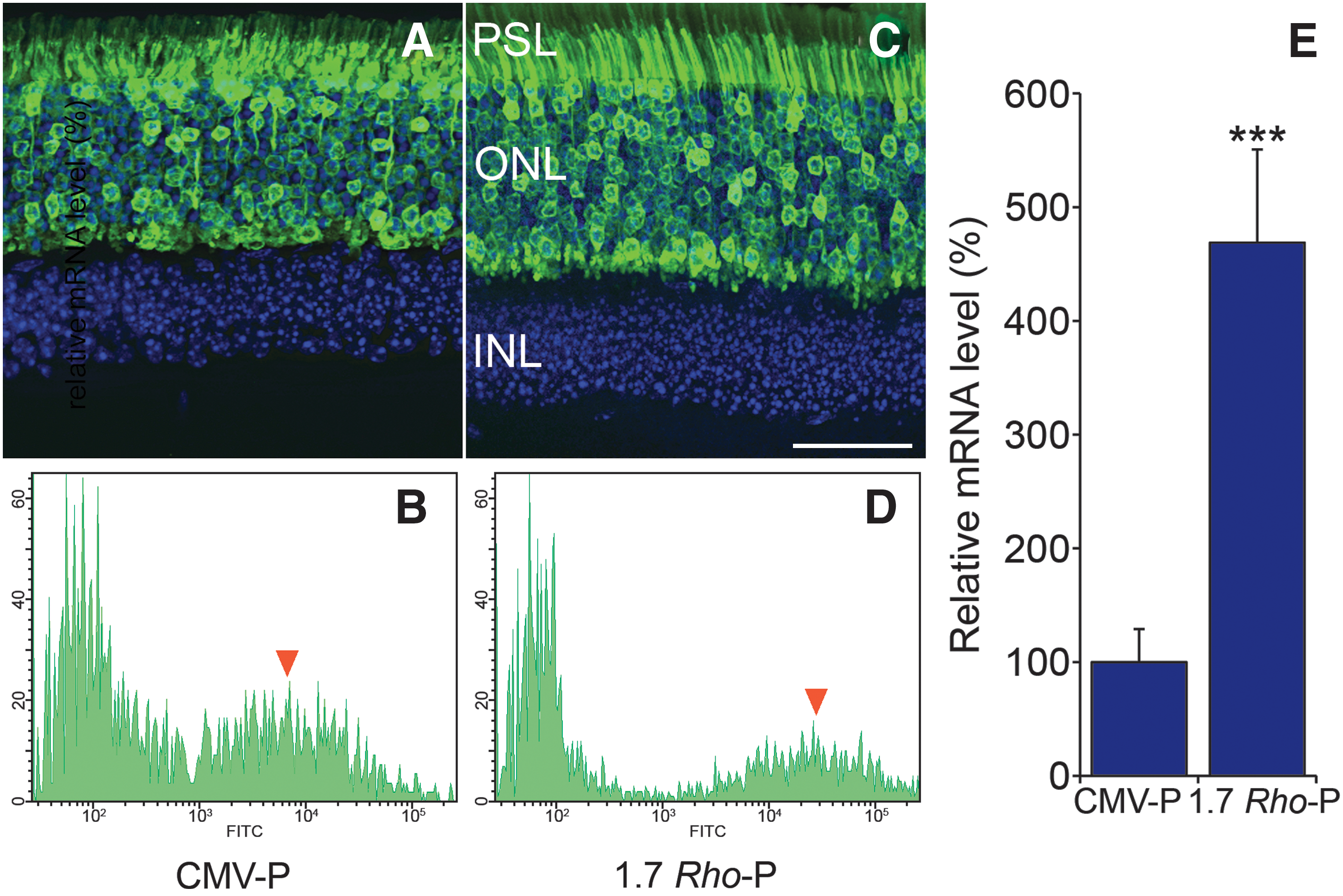
Retinal EGFP expression from CMV- and Rho promoter-driven AAV constructs. Adult wild-type mice were subretinally injected with AAV-CMV-P-EGFP or AAV-Rho-P-EGFP at 1 × 1010 VP/eye. Two weeks postinjection, eyes were fixed in 4% paraformaldehyde and cryosectioned (12 μm) and the nuclei were counterstained with 4′,6-diamidino-2-phenylindole (DAPI), or the eyes were prepared for FACS analysis. Representative microscopy images are shown of eyes injected with AAV-CMV-P-EGFP (
After the initial promoter experiments involving AAV-mediated EGFP expression in murine retina, six RHO replacement constructs termed AAV-BB8 (O'Reilly et al., 2007), AAV-BB9, AAV-BB10, AAV-BB13, AAV-BB24, and AAV-BB32 were engineered (Fig. 1). Construct designs were constrained by the insert size limitation for AAV vectors (approximately 4.5 kb). One set of constructs included large segments of the murine Rho-P whereas the other set of constructs included a shorter Rho-P complemented with CMV enhancer and/or WPRE sequences (Fig. 1). More specifically, AAV-BB8, AAV-BB9, AAV-BB24, and AAV-BB32 included a 1.7-kb Rho-P whereas AAV-BB10 and AAV-BB13 contained a 0.5-kb Rho-P augmented with the CMV enhancer element. AAV-BB13 contained a modified WPRE element (Loeb et al., 1999). In addition, a 0.7-kb fragment of the endogenous RHO 3′ untranslated region (3′-UTR, including polyadenylation signals) was present in all vectors except AAV-BB24 and AAV-BB32. Multiple sequence alignments enabled comparison of rhodopsin promoter sequences from various species, thereby highlighting conserved regions (Supplementary Table 1). AAV-BB24 and AAV-BB32 contained the 1.7-kb Rho-P together with conserved elements of a 1.9-kb fragment of Rho-P (Supplementary Table 1 and Fig. 1), a 0.4-kb fragment of the endogenous 3′-UTR, and a minimal polyadenylation signal (Levitt et al., 1989). AAV-BB8, AAV-BB10, AAV-BB13, and AAV-BB32 were generated with the H1 promoter-driven short hairpin RNA (shRNA) suppressor cassette (H1PshBB), whereas AAV-BB9 and AAV-BB24 were generated without this cassette. Subsequently, AAV preparations in the range of 1012–1013 VP/ml were produced from AAV-BB constructs.
In vivo RHO mRNA expression levels in RNA samples extracted from whole retinas achieved from the six AAV vectors described previously were compared in adult wild-type mice subsequent to subretinal administration of each virus at 1 × 1010 VP/eye (n = 10). Levels of RHO expression in whole retina achieved from AAV vectors were compared with levels of RHO expression observed in RHO-M+/– Rho –/– mice, which express a RHO replacement gene (at 70% of endogenous Rho levels in wild-type mice; O'Reilly et al., 2007, 2008) and display a normal retinal phenotype as evaluated by histology and ERG (O'Reilly et al., 2008). Ten days postinjection of AAV vectors, whole retinas were harvested and RNA was isolated. RHO mRNA expression in wild-type mice treated with AAV constructs, and in RHO-M+/– Rho –/– mice, was confirmed by RNase protection assay (data not shown) and expression levels were determined by qRT-PCR (Fig. 3). Significant RHO mRNA expression from AAV-BB8, AAV-BB9, and AAV-BB10 was detected. However, given the high levels of RHO found endogenously this level of expression represented up to 2% of that found in RHO-M+/– Rho –/– mice (p < 0.001; Fig. 3). No RHO mRNA amplification was observed in uninjected wild-type mice (data not shown). In contrast to AAV-BB8, AAV-BB9, and AAV-BB10, RHO mRNA expression levels from AAV-BB13, AAV-BB24, and AAV-BB32 were significantly higher, that is, 39–66% of that found in RHO-M+/– Rho –/– mice (p < 0.001), representing approximately 40% of that found in wild-type mice. RHO expression increased by 30-fold (p < 0.001), 50-fold (p < 0.001), and 51-fold (p < 0.001) in eyes injected with AAV-BB13, AAV-BB24, and AAV-BB32, respectively, when compared with pAAV-BB8 (Fig. 3). Although the objective of the study was to achieve potent expression of RHO from AAV vectors, comparison of expression between identical vectors except for the addition of the shRNA suppressor, that is, AAV-BB8 versus AAV-BB9 or AAV-BB24 versus AAV-BB32 (Fig. 1), suggested that the H1PshBB suppressor present in AAV-BB8 and AAV-BB32 did not significantly influence expression of the replacement RHO gene (Fig. 3).
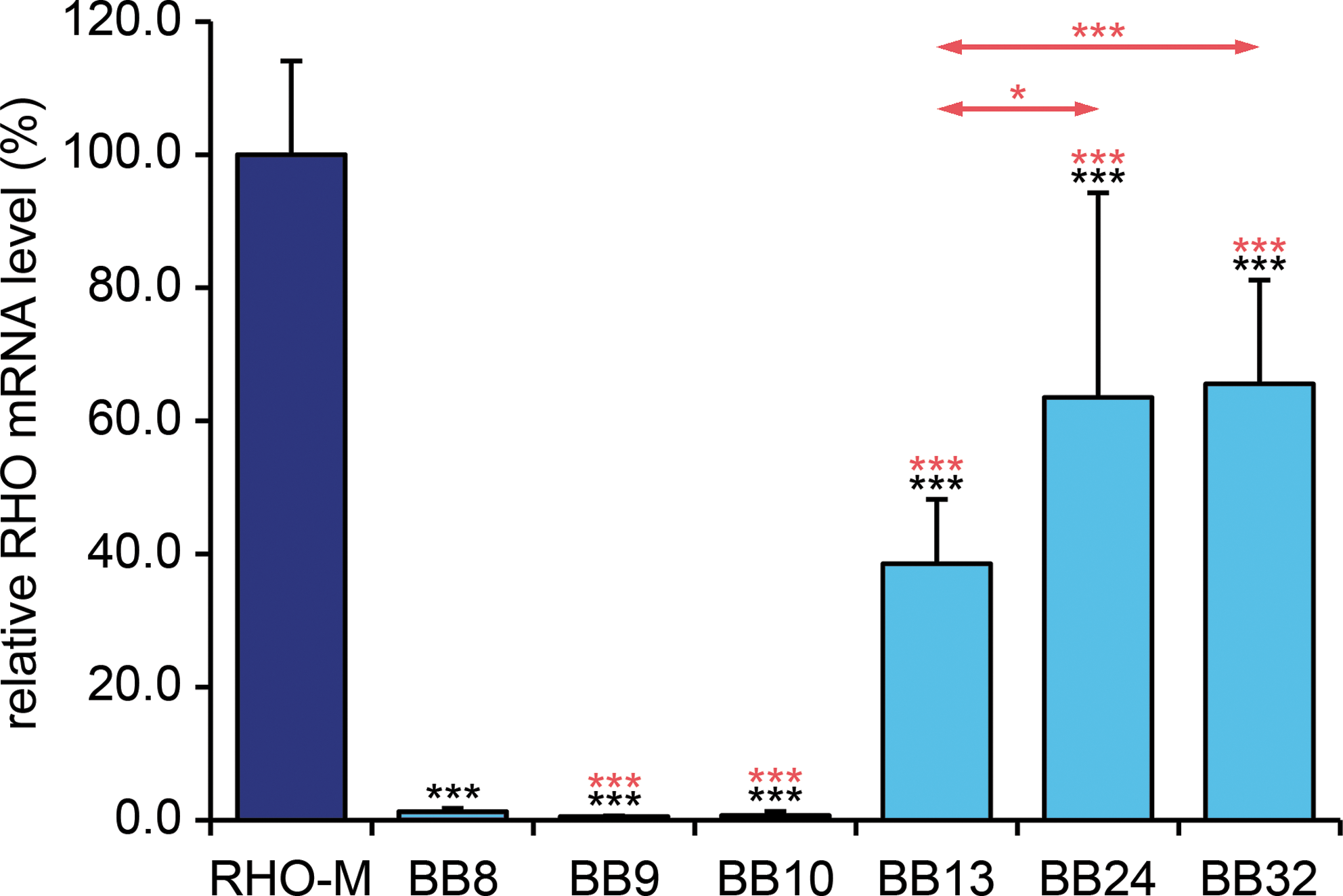
Retinal RHO mRNA expression from various AAV constructs. Adult wild-type mice were subretinally injected at 1 × 1010 VP/eye with AAV-RHO-BB constructs (BB8, BB9, BB10, BB13, BB24, and BB32; n = 10 except for BB32, where n = 5) and RHO mRNA expression was analyzed by qRT-PCR 10 days postinjection. Light blue columns indicate RHO mRNA expression from AAV-transduced retinas. RHO mRNA expression level in RHO-M transgenic mice was used as control and taken as 100% (dark blue column). Error bars represent SD values. Triple black asterisks indicate p < 0.001 (compared with RHO-M). Triple red asterisks indicate p < 0.001 (compared with BB8); single and triple red asterisks above the arrows indicate p < 0.05 (BB24 vs. BB13) and p < 0.001 (BB32 vs. BB13), respectively.
To analyze AAV-mediated RHO expression at the protein level, subretinal injections of AAV-BB8, AAV-BB13, AAV-BB24 (1 × 1010 VP/eye), and AAV-CMV-P-EGFP (2 × 109 VP/eye) vectors were undertaken in adult wild-type mice. The RHO-BB AAVs were coinjected with a 1/10th volume of AAV-CMV-P-EGFP tracer to facilitate localization of transduced areas of retinas. Two weeks postinjection, immunocytochemistry using rhodopsin antibody, which detects the human protein but not that of the mouse, was undertaken (Fig. 4). Significantly higher levels of RHO expression in retinas administered AAV-BB13 (Fig. 4E and F) or AAV-BB24 (Fig. 4G and H) were obtained when compared with retinas administered AAV-BB8 (Fig. 4C and D). RHO labeling was confined to the photoreceptor segment layer (PSL) whereas EGFP was expressed in both the outer nuclear layer (ONL) and the PSL. RHO-M Rho –/– transgenic mice and AAV-CMV-P-EGFP subretinal administration were used as controls for RHO labeling (Fig. 4I and J) and injections (Fig. 4A and B), respectively.
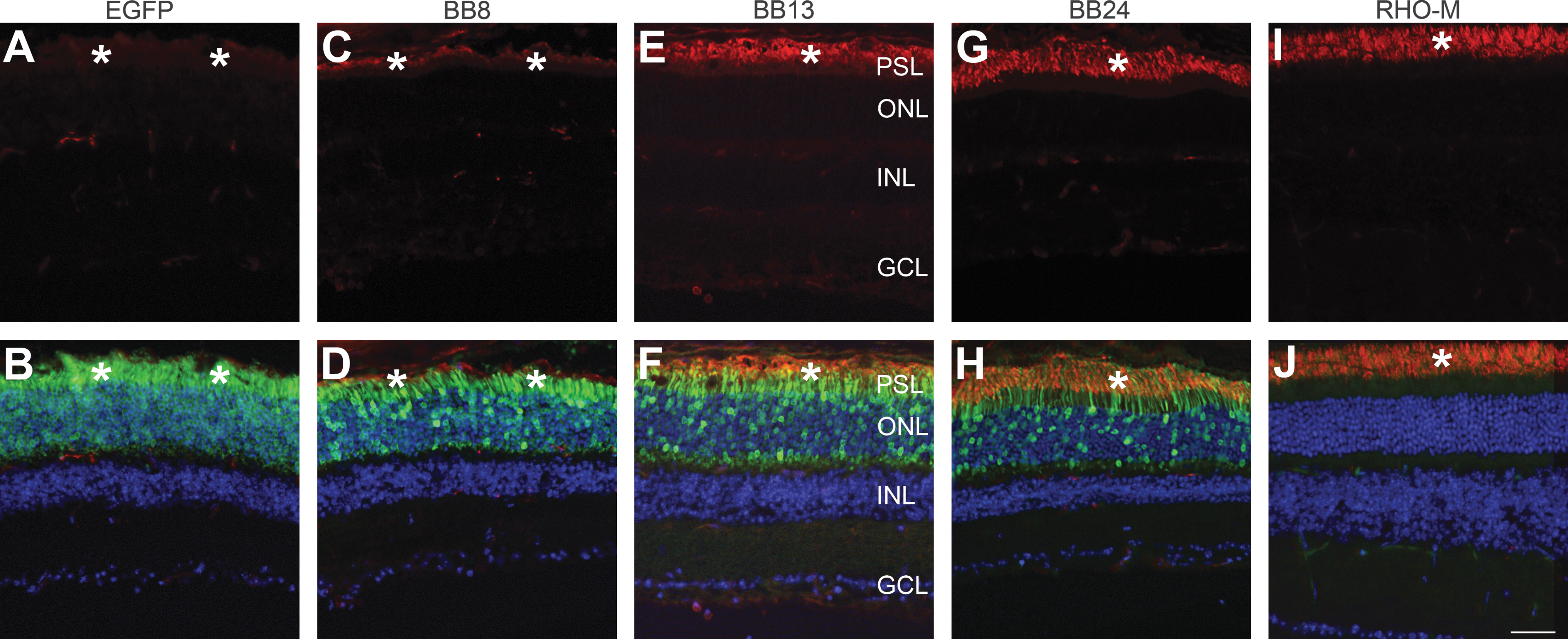
Immunohistochemical analysis of RHO expression in AAV-transduced retinas of wild-type mice. Adult wild-type mice were subretinally injected at 2 × 109 VP/eye with AAV-CMV-P-EGFP (
Two constructs, AAV-BB13 and AAV-BB24, expressing high levels of rhodopsin mRNA and protein in wild-type mice were further explored as a means to express RHO in Rho –/– mice. Initially, subretinal delivery at 3.9 × 109 VP/eye of AAV-BB13 and AAV-BB24 vectors (including a 1/10th volume of AAV-CMV-P-EGFP tracer) was undertaken in Rho –/– mice at P0 (n = 5) and expression was analyzed by immunocytochemistry 6 weeks postinjection. Significant RHO expression in the ONL and PSL, colocalized with the EGFP tracer, was detected in treated Rho –/– mouse retinas using both AAV vectors (Fig. 5C–F). The pattern of RHO labeling observed resembled that found in wild-type mouse retinas and the morphology of the PSL suggested the presence of ROS in treated Rho –/– mouse eyes (Fig. 5E and F). In contrast, RHO immunolabeling (or EGFP expression) was not observed in uninjected eyes (Fig. 5A and B). As is well documented, ROS do not develop in Rho –/– retinas (Humphries et al., 1997). Analysis of AAV-BB24 (3.9 × 109 VP/eye)-transduced Rho –/– mouse retinas at 12 weeks postinjection (n = 5; Fig. 6) indicated a pattern of RHO expression (Fig. 6C and D) similar to that observed at 6 weeks (Fig. 5). Photoreceptor cell numbers were decreased relative to the 6-week time point (Fig. 5) but still consisted of three or four rows of ONL (Fig. 6C and D), whereas in uninjected control eyes the ONL was composed of just a single-cell row of cone nuclei (Fig. 6B).
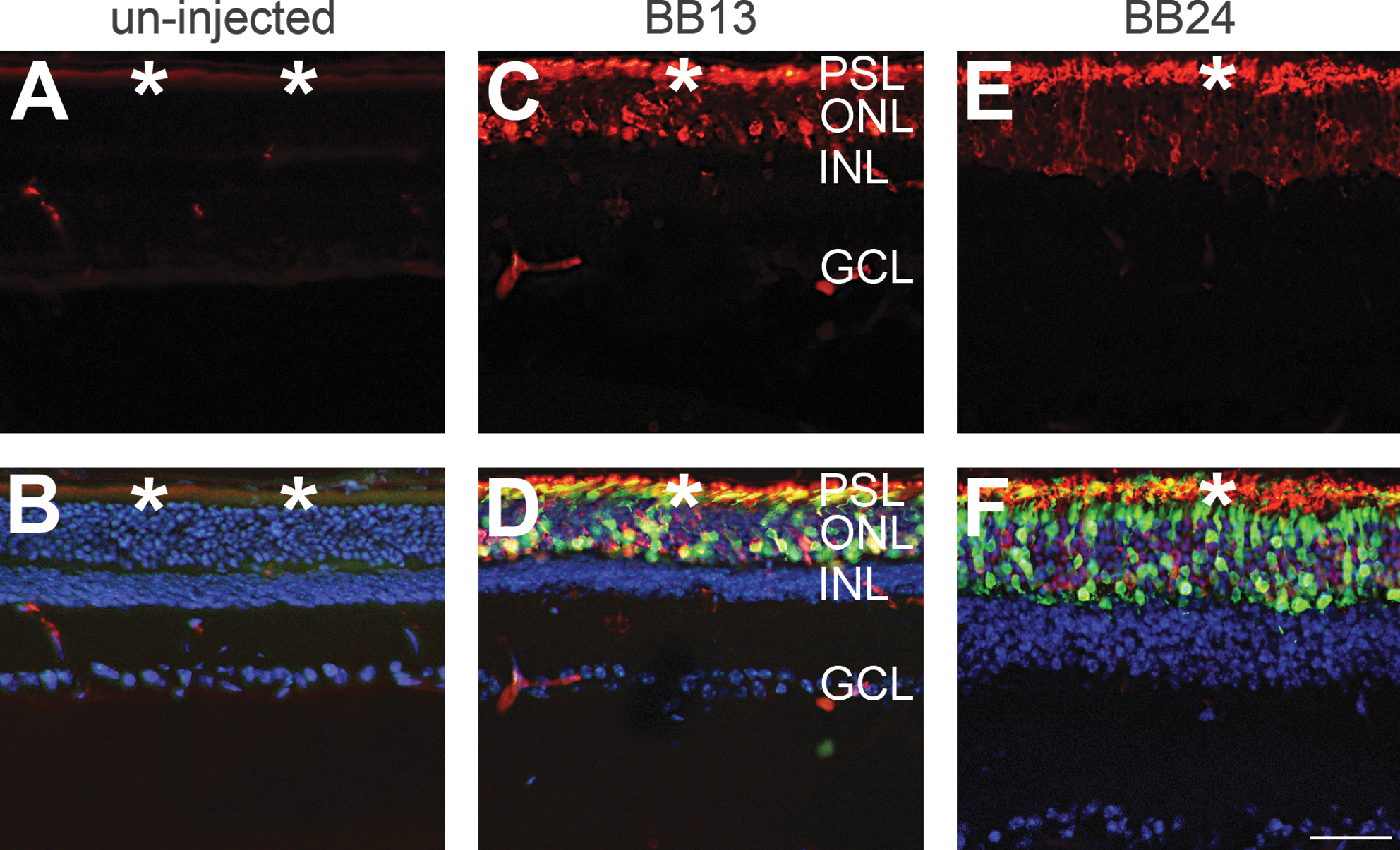
Immunohistochemical analysis of RHO expression in AAV-transduced retinas of Rho
–/– mice 6 weeks postinjection. The right eyes of newborn Rho–/–
mice were subretinally injected at 3.9 × 109 VP/eye with AAV-BB13 (
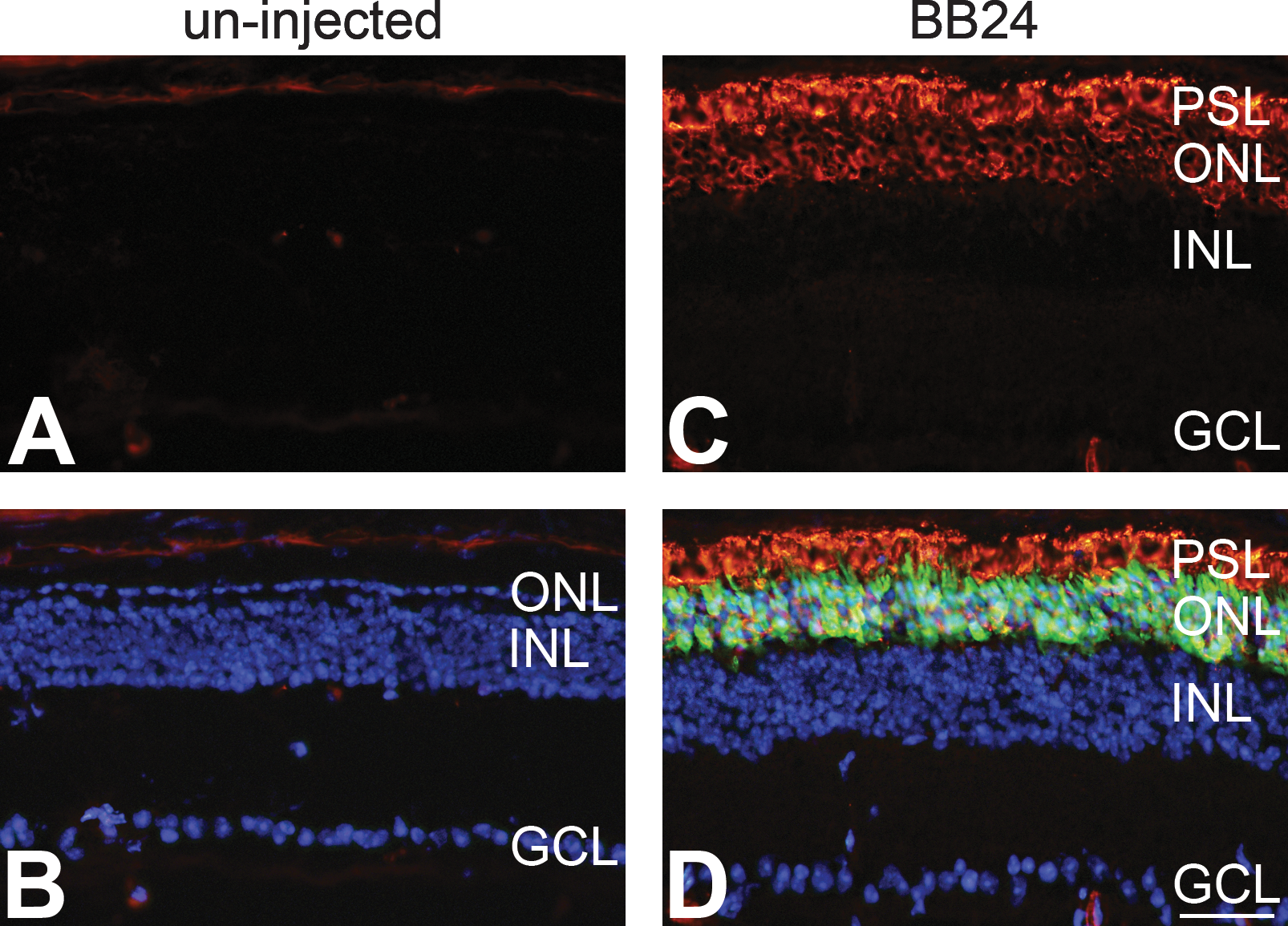
Immunohistochemical analysis of RHO expression in AAV-transduced retinas of Rho–/–
mice 12 weeks postinjection. The right eyes of newborn Rho–/–
mice (n = 5) were subretinally injected at 3.9 × 109 VP/eye with AAV-BB24 (BB24) supplemented with a 1/10 volume of AAV-CMV-P-EGFP tracer (
A more extensive analysis was undertaken with AAV-BB24. Two different doses of AAV-BB24, 3.9 × 109 VP/eye (n = 10) and 6.5 × 109 VP/eye (n = 3), both including a 1/10th volume of pAAV-CMV-P-EGFP tracer, were subretinally injected into the right eyes of Rho
–/– mice at P0 whereas the left eyes were uninjected; AAV-Rho-P-EGFP and PBS were used as controls for injection. Six weeks postinjection, ERG analyses were undertaken on the injected and control eyes from these mice. Notably, rod-isolated ERG recordings of AAV-BB24-injected Rho
–/– mouse eyes provided evidence of significant rod photoreceptor function compared with fellow uninjected Rho
–/– eyes, in which rod-generated responses were not detected (Fig. 7). In the uninjected versus injected eyes, means of rod-isolated b-wave amplitudes were 4.0 ± 5.2 μV compared with 37.6 ± 14.5 μV (p < 0.001; Fig. 7A and B) at the lower AAV dose whereas they were 4.7 ± 3.6 μV compared with 135.3 ± 58.5 μV (p < 0.05; Fig. 7C and D) at the higher AAV dose. The mean value of control rod-isolated b-wave amplitudes was 703.6 ± 116.1 μV (n = 8) in age-matched RHO-M mice (Fig. 7E and F). In contrast, cone-isolated ERGs (b-wave amplitudes; see Supplementary Fig. 1 at
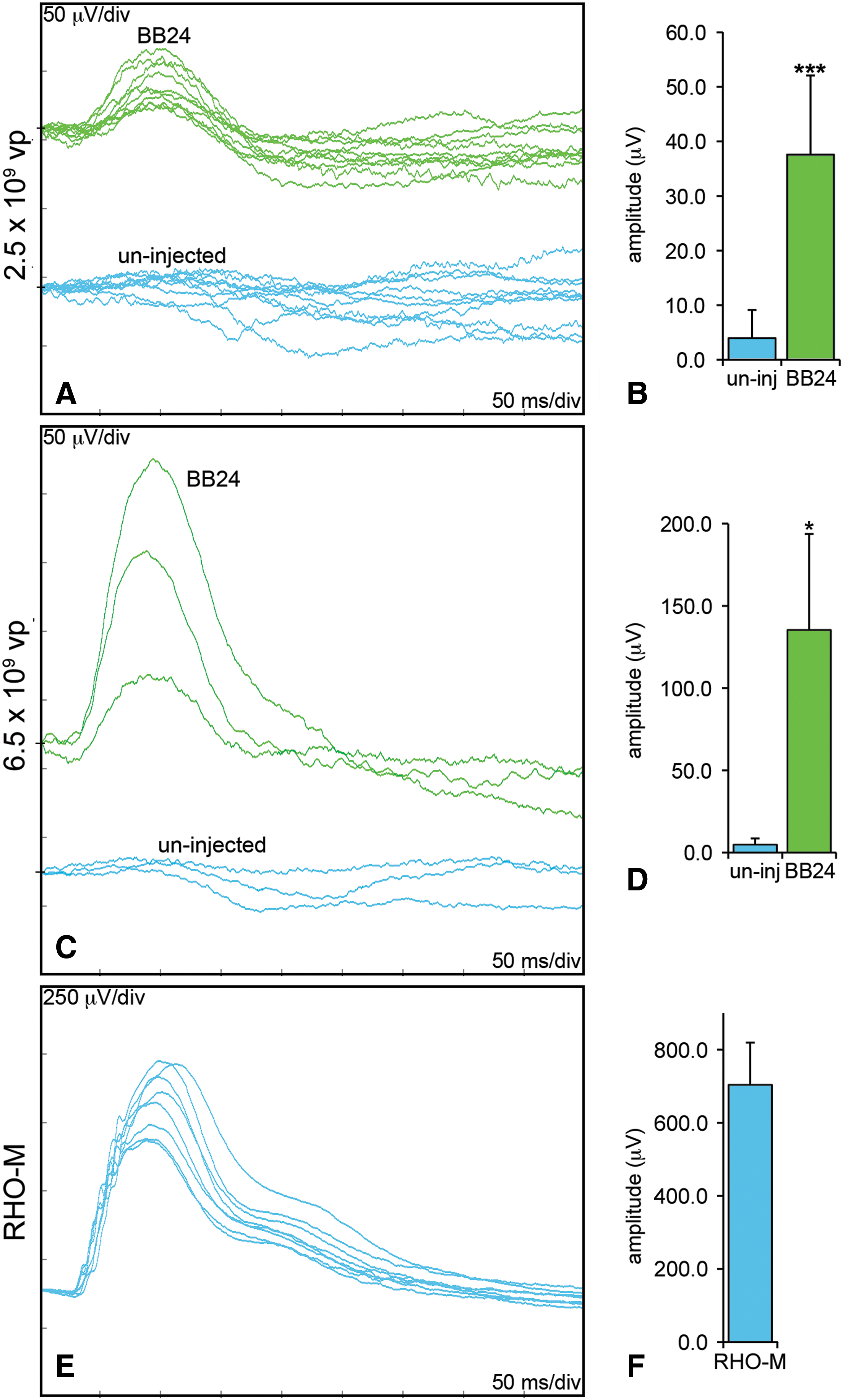
Rod-derived ERGs of Rho–/–
mice 6 weeks postinjection of AAV-BB24. The right eyes of newborn Rho–/–
mice were subretinally injected with two doses, either 3.9 × 109 VP/eye (n = 10) or 6.5 × 109 VP/eye (n = 3), of AAV-BB24, including a 1/10th volume of AAV-EGFP tracer, whereas the left eyes were uninjected. Six weeks postinjection, mice were dark-adapted overnight and rod-isolated ERG responses were recorded from both eyes; age-matched RHO-M mice were used as controls (n = 8). (
In a further set of experiments, rod-isolated ERGs recorded 6 and 12 weeks after injection of AAV-BB24 (3.9 × 1012 VP/eye, n = 12) on P0 into right eyes were compared; left eyes were uninjected. Considerable rod-ERG response was detected 12 weeks postinjection (20.6 ± 19.3 μV; Fig. 8) but b-wave amplitudes were significantly lower than at 6 weeks (61.9 ± 35.6 μV; p < 0.01) (Fig. 8). Although the b-wave amplitudes varied, a general trend toward a decrease in b-wave amplitudes of individual animals was observed between 6 and 12 weeks (Fig. 8). Notably, no rod function was detected in control uninjected eyes at either 6 weeks (Fig. 7) or 12 weeks (data not shown).
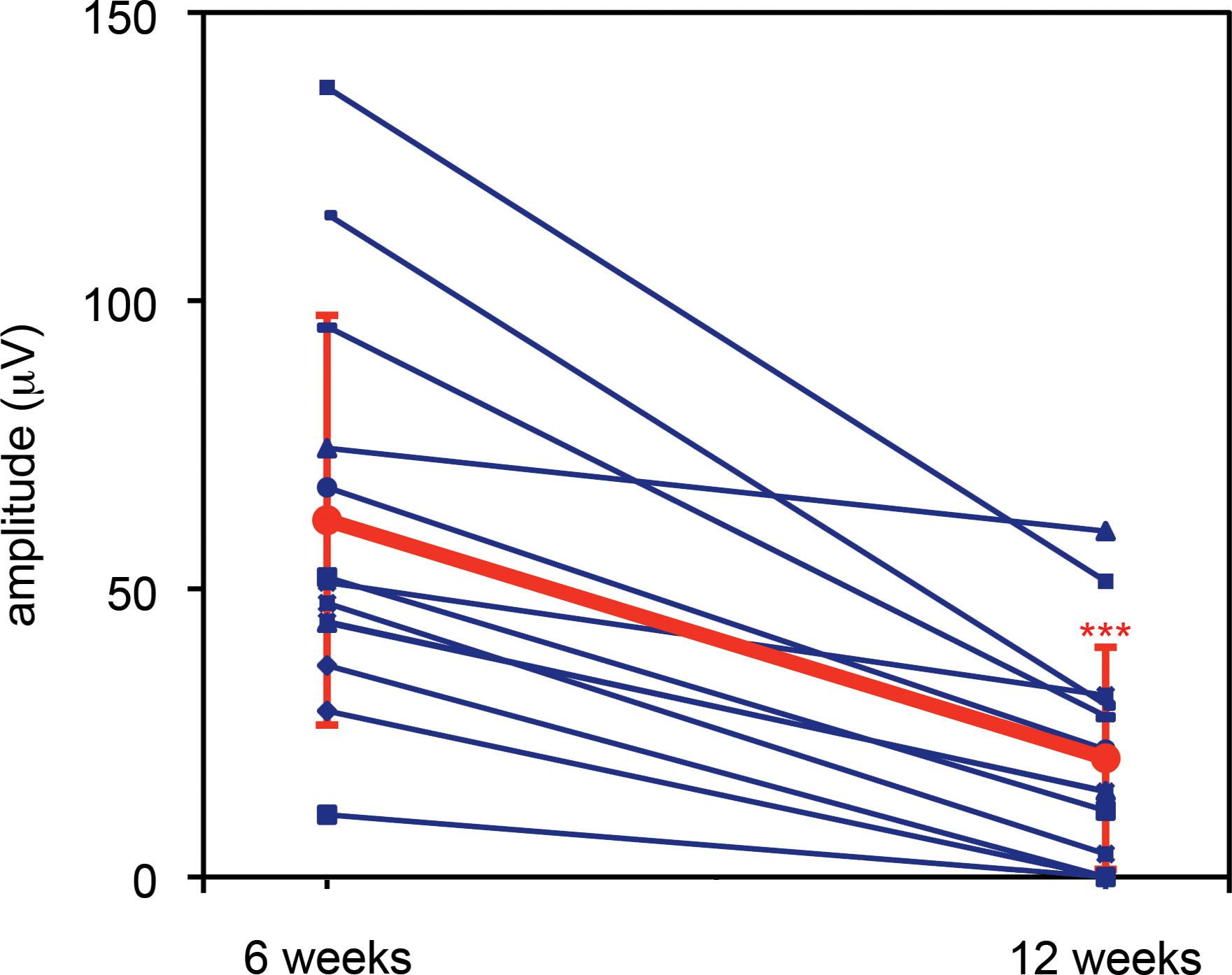
Comparison of rod-derived ERGs of Rho–/–
mice 6 and 12 weeks after subretinal administration of AAV-BB24. The right eyes of newborn Rho–/–
mice were subretinally injected at 3.9 × 109 VP/eye (n = 12) with AAV-BB24 (including a 1/10th volume of AAV-EGFP tracer) whereas the left eyes remained uninjected. Rod-isolated ERGs were recorded at both 6 and 12 weeks postinjection in each mouse; mice were dark-adapted overnight before analysis. b-Wave amplitudes at 6 and 12 weeks are given for individual animals (blue). The means of amplitudes are given in red. Error bars represent SD values. ***p < 0.001. Color images available online at
The histology of AAV-BB24-transduced Rho
–/– retinas was further analyzed at the ultrastructural level 6 weeks after injection of 6.5 × 109 VP of AAV-BB24 (n = 3) including a 1/10th volume of AAV-CMV-P-EGFP tracer. Paraformaldehyde-fixed eyes were whole mounted and, using the EGFP tracer, transduced and untransduced areas were identified; the transduced area covered by a single injection was 38.8 ± 7.9% of the retina (n = 3; Fig. 9B). Representative areas of uninjected retinas (Fig. 9A) and of injected untransduced and injected transduced retinas (Fig. 9B) were excised and processed for resin embedding. Semithin sections from these eyes (Fig. 9C, E, and G) indicated a marked protection of retinal structure in the AAV-BB24-transduced areas (Fig. 9G), including more extensive nuclear and photoreceptor segment layers compared with the untransduced or uninjected areas (Fig. 9C and E). Using TEM, ROS were detected in the AAV-BB24-transduced areas, with ROS reaching up to the retinal pigment epithelium (RPE) (Fig. 9H). Only rod photoreceptor inner segments (RIS) were detected in the untransduced or uninjected retinal areas (Fig. 9D and F); also note the wide extracellular space between the photoreceptor segments and the RPE cells, which was confirmed at higher magnification (Fig. 10A). TEM at higher magnification also revealed that RIS in the untransduced areas possessed ciliary processes (connecting cilium), which in wild-type photoreceptors connect the inner and the outer segments. However, in these photoreceptors, truncated outer segments lacking membrane disks were observed at the outer tips of the connecting cilia (Fig. 10A). On the other hand, in the transduced areas correctly formed ROS with parallel membrane disks typical of rod photoreceptor cells attached to the connecting cilium were present. The lengths of the ROS were found to be 9.52 ± 1.08 μm (n = 9). Note that the average length of the ROS in wild-type mouse retinas is 23.8 ± 1.32 μm (Nickell et al., 2007). The ROS of the rescued photoreceptors connected to the RPE (Fig. 10C) and disk membrane stacks shed from the ROS tips appeared to be phagocytosed by cells of the RPE (Supplementary Fig. 2 at
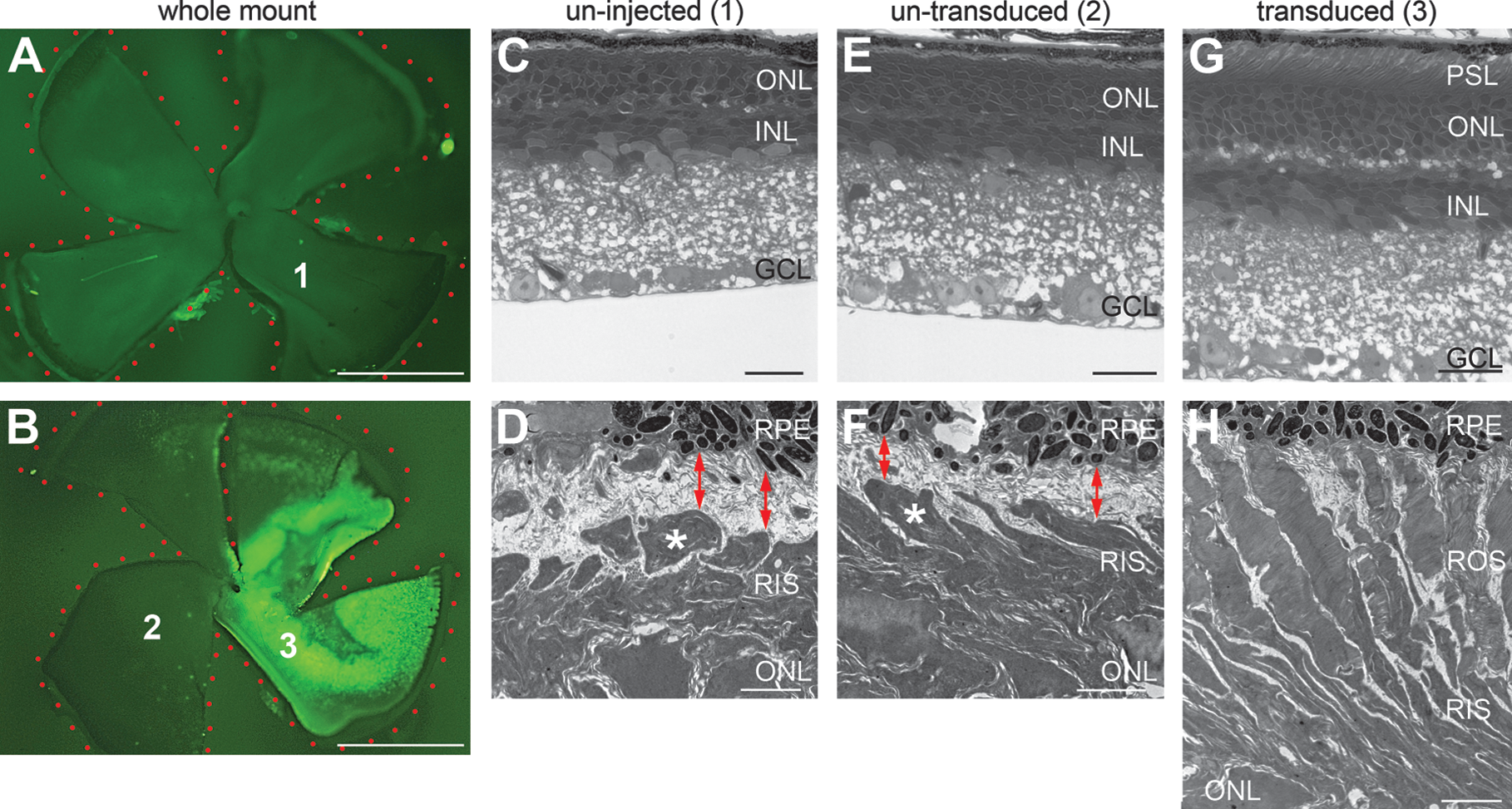
Ultrastructural analysis of Rho–/–
retinas transduced with AAV-BB24. The right eyes of newborn Rho–/–
mice (

Photoreceptor rescue in Rho–/–
retinas transduced with AAV-BB24. Transmission electron micrographs of longitudinal ultrathin sections through parts of rod photoreceptors of untransduced (
Discussion
GPCRs constitute an enormous family of transmembrane molecules involved in signal transduction and are fundamental to a multiplicity of cellular processes (Palczewski, 2006). Rhodopsin represents one of the most extensively studied GPCRs; investigations were facilitated by the relatively large quantities of protein found in the disk membranes of rod photoreceptor cells. The continual replenishing of ROS, in which rhodopsin represents 90% of the protein content, suggests that extensive renewal with de novo rhodopsin molecules is required for optimal visual function (Palczewski, 2006; Winkler, 2008). The high level of rhodopsin expression in ROS, although advantageous for early studies focused on delineating the function of the protein in phototransduction, presents a significant challenge in terms of gene therapy, where the principal objective is to recapitulate endogenous levels of rhodopsin expression from an exogenously delivered vector. Therefore the focus of the current study was to explore ways to achieve replacement RHO expression from AAV at appropriate levels for in vivo biological functionality.
When selecting a viral vector for gene delivery various issues must be considered including tropism of the vector for the target cell, the viral titers that can be achieved, the safety and immune tolerance associated with the virus, and available clinical information regarding administration to humans. Given these issues, AAV represents an attractive vector option for photoreceptors (Lebherz et al., 2008; Li et al., 2008; Surace and Auricchio, 2008; Stieger et al., 2009). However, AAV has a limited capacity of approximately 4.5 kb for most serotypes, thereby curtailing the insert size that can be engineered into AAV. Larger inserts of approximately 9 kb have been incorporated into AAV2/5 (Allocca et al., 2008), although levels of expression from these vectors have still to be fully elucidated. In the current study, to explore the feasibility of achieving high levels of expression of a RHO replacement gene, a range of AAV RHO replacement vectors was produced. The human RHO replacement gene used was modified such that it is resistant to the shRNA expressed from H1PshBB but encodes wild-type RHO. The replacement gene characterized in this study may be of value in future human suppression and replacement therapies.
Expression levels provided by CMV-P and the 1.7-kb Rho-P were determined with AAV-CMV-P-EGFP and AAV-Rho-P-EGFP, respectively (Fig. 2). Rho-P was found to result in 5-fold higher levels of RHO expression than CMV-P in murine retinas (Fig. 2) and therefore for reasons of specificity and superior expression, this promoter was progressed further. A subsequent comparative analysis of six AAV Rho-P RHO replacement constructs was undertaken in vivo to assess levels of RHO expression achieved with these vectors. Expression levels of RHO mRNA were determined in RNA samples extracted from whole retinas. Therefore the RHO mRNA expression levels in the transduced areas were expected to be higher.
Notably, RHO promoter-driven expression of EGFP and RHO resulted in a differential subcellular distribution of the two proteins in the adult retina. EGFP was localized in the cell body (Fig. 2) and RIS whereas RHO was detected mainly in ROS (Fig. 4). Evidence suggests that trafficking to ROS represents the default location for photoreceptor membrane proteins such as RHO (Baker et al., 2008) and that membrane proteins locate elsewhere only if they contain specific targeting signals (Baker et al., 2008). In addition, an ROS-targeting signal in the C terminus of RHO has been reported (Tai et al., 1999; Deretic, 2006). On the other hand, EGFP, a soluble protein, localizes in the cytosol including the cell body and RIS but is not evident in ROS. Sequential inclusion/exclusion of various elements and conserved sequences associated with the rhodopsin promoters in AAV vectors served to improve significantly the expression of the RHO replacement gene (Figs. 1 and 3, and Supplementary Table 1). In particular, inclusion of the WPRE sequence or regions of the murine Rho-P served to radically increase rhodopsin expression. RNA results were mirrored by evaluation of RHO protein expression in wild-type (Fig. 4) and Rho –/– mice (Figs. 5 and 6). WPRE was included in AAV-BB13 3′ of the first polyadenylation signal to minimize read-through and aid polyadenylation (Higashimoto et al., 2007). Note that RHO is localized mainly to the ROS in adult retinas (Fig. 4), whereas it is also detected in the cell bodies of rods in the developing retina (Figs. 5 and 6). The mechanisms underlying such improvements in expression may possibly be related to the presence of binding sites for transcription factors such as neural retinal leucine zipper factor (NRL) and cone rod homeobox-containing transcription factor (CRX) in the Rho-P sequences used (Hennig et al., 2008; Oh et al., 2008).
As the inclusion of additional murine Rho-P sequences in AAV-BB24 provided high levels of RHO expression, a detailed analysis was undertaken with this vector in Rho –/– mice. Subretinal delivery of AAV-BB24 to P0 Rho –/– mice followed by ERG 6 and 12 weeks postadministration of virus provided evidence of significant rod photoreceptor-derived ERG responses in Rho –/– mice (Figs. 7 and 8). The rod-derived amplitudes declined with time and this was mirrored by reduced ONL thickness (Fig. 6) at 12 weeks postinjection. However, there was a clearly visible improvement compared with uninjected eyes, where only a single photoreceptor cell layer remained and no recordable rod-ERG response was detected. A compromised cone-ERG response was observed in Rho –/– mice injected with AAV-BB24 or PBS alone on P0 (Supplementary Fig. 1); causative mechanisms will require future investigation in terms of possible surgical trauma. Notably, less of a decrease in b-wave amplitude was observed in mice subretinally injected on P10. TEM provided unequivocal evidence of the presence of ROS in AAV-BB24-treated Rho –/– mouse eyes (Figs. 9 and 10, and Supplementary Fig. 2). Indeed, the lengths of the AAV-BB24-induced ROS were on the order of one-third those present in wild-type mice (Nickell et al., 2007). Although the capability of retinal structures such as ROS for plasticity and recovery has still to be fully elucidated, the current study provides some additional insights into this feature of ROS (Schremser and Williams, 1995; Liang et al., 2004; Wen et al., 2009).
The aim of the study was to develop AAV-RHO vectors that can achieve levels of RHO gene expression that provide functional rods in Rho –/– mice. Although this in itself was a significant achievement, it was also demonstrated that the presence of an RNA interference (RNAi)-based suppressor gene (H1PshBB; targeting wild-type RHO) in some vectors did not influence expression of the sequence-modified RHO-BB replacement gene (Figs. 1 and 3), mirroring the results from a prior study in vitro (O'Reilly et al., 2007). Such RNAi suppressors and RNAi-resistant replacement genes will be required as components of future therapies, in particular for gain-of-function RHO mutations, where potent suppression in conjunction with efficient rhodopsin replacement may provide therapeutic benefit for RHO-adRP.
It is worth highlighting challenges associated with ameliorating the Rho –/– phenotype by gene replacement. Rho expression initiates at approximately P4 postnatally in the murine retina (McNally et al., 1999), increasing in levels of expression until Rho comprises 90% of ROS disk membranes (Palczewski, 2006). To increase the likelihood that the appropriate cellular machinery accompanying Rho expression from P4 is invoked within the correct time frame in AAV-BB24-treated Rho –/– mice, it was deemed optimal to deliver AAV-BB24 early, that is, on P0. Even using P0 injections, initial histologically observable expression from the AAV2/5 serotype occurs approximately 1 week postinjection in murine retinas. The volume, and hence viral dose, that can be injected subretinally into P0 mice is limited to 0.5 μl. Moreover, viral spread in P0 Rho –/– mice is limited (Fig. 9B), thereby also restricting the extent of possible rescue. Because AAV has a modest insert capacity, recapitulating endogenous levels of rhodopsin from AAV vectors is a significant task. However, in the current study we have made considerable strides toward achieving this goal with associated significant structural and functional benefits in Rho –/– mice.
Despite the limitations and challenges outlined previously, this is the first demonstration of substantial rescue of aspects of the retinal degeneration in Rho –/– mice, which lack this highly expressed GPCR. This study also represents significant progress toward the development of gene-based therapies for RHO-adRP, the most common form of RP, by providing a functionally relevant replacement rhodopsin gene.
Footnotes
Acknowledgments
The authors thank Prof. W. Baehr (University of Utah, Salt Lake City, UT) for the original RHO cDNA construct, Prof. T. Giroglou (University of Frankfurt, Frankfurt, Germany) and Prof. T.J. Hope (The Salk Institute, La Jolla, CA) for providing the vectors containing the woodchuck hepatitis virus posttranscriptional regulatory elements, Prof. R.S. Molday (University of British Columbia, Vancouver, BC, Canada) for the two rhodopsin primary antibodies, Dr. Alfonso Blanco Fernandez (Flow Cytometry Core Facility, University College, Dublin, Ireland) for assisting with FACS analysis, Elisabeth Sehn (University of Mainz, Mainz, Germany) for skillful technical assistance, and the staff of the Bioresources Unit (Trinity College Dublin, Dublin, Ireland). This work was supported by Science Foundation Ireland; Foundation Fighting Blindness – National Neurovision Research Institute (USA); European Union RETNET (MRT-CT-2003-504003); EviGenoRet (LSHG-CT-2005-512036); Deutsche Forschungsgemeinschaft (GRK1044/1); and FAUN-Stiftung.
Author Disclosure Statement
J.F. and P.K. are directors of Genable Technologies; S.M.-W., N.C., A.P., and M.O'R. are consultants for Genable Technologies. These authors have competing interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.