
Abstract
To assess the feasibility of a gene therapeutic approach to treating choroidal neovascularization (CNV), we generated a recombinant adeno-associated viral (AAV) vector (type 8) encoding soluble Flt-1 (AAV-sflt-1), and determined its ability to inhibit angiogenesis. When we treated human umbilical vein endothelial cells (HUVECs) with the supernatant of cells transduced with AAV-sflt-1 or AAV-EGFP (control), we found that tube formation was significantly inhibited by the former but not the latter (area: 25,121 ± 557 vs. 68,628 ± 1357 pixels [p < 0.01]; length: 4811 ± 246 vs. 10,894 ± 297 pixels [p < 0.01]). CNV was induced in C57BL/6 mice by making four separate choroidal burns around the optic nerve in each eye, using a diode laser. Thereafter, 2 μl (5 × 1011 vector genomes/ml) of AAV-sflt-1 (n = 11) or control AAV-LacZ (n = 12) was injected into the subretinal space, and 2 weeks later the eyes were removed for flatmount analysis of CNV surface area. Notably, subretinal delivery of AAV-sflt-1 significantly diminished CNV at the laser lesions, as compared with AAV-LacZ (555 ± 304 vs. 1470 ± 1000 μm2; p = 0.007). These results suggest that there was diffusion of the secreted sFlt-1 across the retina and that long-term suppression of CNV is possible through the use of stable rAAV-mediated sflt-1 expression. In vivo gene therapy thus appears to be a feasible approach to the clinical management of CNV in conditions such as age-related macular degeneration.
Introduction
The eye has unique advantages as a target organ for the development of gene therapies. First, the eye is easily accessible using standard minor surgical procedures, making delivery of a vector a relatively simple procedure; second, the eye is a fairly small organ with a highly compartmentalized anatomy into which it is possible to precisely deliver small volumes of adeno-associated viral (AAV) vectors and to obtain efficient and stable transduction; and third, the eye enjoys a relatively immune-privileged status due to a low abundance of antigen-presenting cells and the presence of factors that actively suppress T cell activation, which facilitates stable expression of the transgene (Bennett and Maguire, 2000). AAV-mediated gene therapy is thus a promising strategy for the treatment of eye disease. To date, most studies of gene therapeutic approaches to treating eye disease have used type 2 AAV vectors, but transduction efficiency may be somewhat greater with type 5 AAV, particularly for photoreceptors (Yang et al., 2002). In addition, Lebherz and colleagues showed that types 7 and 8 AAV vectors enable better long-term transduction of retinal structures than types 1, 2, and 5 vectors (Lebherz et al., 2008). In the present study, we compared the efficacies of the type 5 and 8 AAV vectors for transduction with that of our murine CNV model and then assessed the effect of the soluble flt-1 (sFlt-1) transgene when delivered by AAV type 8 in the same model.
Materials and Methods
Generation of AAV vectors
To construct the pAAV-FLK plasmid, cDNA encoding the soluble extracellular domain of the human soluble vascular endothelial growth factor receptor-1 (hsFlt-1) was purchased (pBLAST45-hsFLT-1s7; InvivoGen, San Diego, CA). A 2.2-kb NcoI–EcoRI fragment containing the hsflt-1 transgene was excised, treated with Klenow, and ligated into the PvuII site of pSP72 (Promega, Madison, WI), yielding p72-FLT. The XhoI–SalI fragment of p72-FLT was then cloned into the PvuII fragment of the pSP72 plasmid containing the cytomegalovirus early enhancer/chicken β-actin (CAG) promoter (Noro et al., 2004). Finally, the SalI fragment was cloned into the XhoI–SalI site of the AAV plasmid pAAV.CAaGBE (Takahashi et al., 2002). As controls we used pAAV-EGFP or pAAV-LacZ, which contain the green fluorescent protein (GFP)-encoding or lacZ gene, respectively, driven by the CAG promoter. AAV vectors (AAV-sflt-1, AAV-EGFP, and AAV-LacZ) were generated with an adenovirus-free system. The virions were purified by discontinuous iodixanol gradient centrifugation as described previously (Kurai et al., 2007), after which the particle titers of the purified virions were determined by quantitative slot-blot analysis.
In vitro tube formation assay
In vitro tube formation was assayed as reported previously (Igarashi et al., 2003). Briefly, assays of tube formation were conducted in triplicate in 24-well dishes, using an angiogenesis kit (Kurabo, Okayama, Japan) according to the manufacturer's instructions. Human umbilical vein endothelial cells (HUVECs) cocultured with human fibroblasts were treated with 100 μl of supernatant from type 8 AAV-sflt-1- or control AAV-EGFP (5 × 1011 vector genomes/ml)-transduced C2C12 (murine myoblast) cells, or left untreated (mock), and were then cultured in medium containing VEGF (10 ng/ml). The medium was changed every 3 days. After 10 days, the dishes were washed with phosphate-buffered saline (PBS) and the cells were fixed with 70% ethanol at 4°C. The fixed cells were then rinsed three times with PBS, after which they were incubated for 60 min with mouse anti-human CD31 (Kurabo) in PBS containing 1% bovine serum albumin (BSA), washed three times with 1% BSA–PBS, and then incubated with alkaline phosphatase-conjugated goat anti-mouse IgG (Kurabo). Metal-enhanced 3,3′-diaminobenzidine-tetrahydrochloride (DAB) was used as the substrate, and the reaction yielded a dark reddish-brown insoluble end-product. Finally, the cells were washed five times with PBS and viewed under an Olympus microscope.
Generation of murine laser photocoagulation-induced CNV model
To generate the model, 2-month-old male C57BL/6 mice were first deeply anesthetized by intramuscular injection of ketamine hydrochloride and intraperitoneal injection of pentobarbital sodium. In addition, anesthetic eye drops were applied, after which the pupils were dilated with one drop of 0.5% tropicamide (Mydrin-M; Santen Pharmaceutical, Osaka, Japan) for photocoagulation (PC). The beam from a diode laser (532 nm; Lumenis, Yokneam, Israel) was shone onto to the retina through a slit lamp biomicroscope, using a 22-mm coverslip as a contact lens. The treatment parameters were chosen to produce a cavitation bubble in the choroid without hemorrhage (spot size, 100 μm2; intensity, 120 mW; duration, 100 msec). Mice with obvious media opacities (i.e., in the cornea and/or lens) were excluded. Four laser burns were made at the 3, 6, 9, and 12 o'clock positions of the posterior pole, around the optic nerve in both eyes. The vector was delivered by subretinal injection at the posterior limbus, using a Hamilton syringe with a 33-gauge needle.
Histological analysis
Two weeks after subretinal injection of type 5 or type 8 AAV-EGFP into untreated C57BL/6 mice (PC–) or laser photocoagulation-treated CNV model mice (PC+), the eyes were enucleated, fixed overnight at 4°C, and sequentially transferred to 10 mM PBS containing 10, 20, or 30% sucrose every 2 hr. The eyes were then frozen in O.C.T. compound on dry ice, and 2-μm-thick cryostat sections were cut in a plane parallel to the vertical meridian of the eye. After mounting with medium containing 4′,6-diamidino-2-phenylindole (DAPI; Vector Laboratories, Burlingame, CA), the specimens were observed under an Olympus fluorescence microscope (Olympus, Tokyo, Japan).
Detection of CNV by fluorescein angiography
Two weeks after type 8 AAV-sflt-1 or AAV-LacZ injection, anesthetized mice were killed by cardiac perfusion of 40 ml of PBS, 5 ml of 4% paraformaldehyde in PBS, and 2 ml of a mixture of fluorescein isothiocyanate (FITC)-conjugated high molecular weight dextran (molecular weights, 2 × 106 and 4 × 104 Da in a proportion of 2:1 at a concentration of 10 mg/ml; Sigma-Aldrich, St. Louis, MO). The eyes were then enucleated and fixed in 4% paraformaldehyde overnight at 4°C. A technique for visualizing FITC–dextran-perfused vessels within CNV was modified to enable simultaneous visualization of cell nuclei within the lesions. The anterior segment and the neurosensory retina were removed, and four radial relaxing incisions were made in the remaining sclera–choroid–retinal pigmental epithelial (RPE) complex. Green FITC fluorescence from within the neovascular complexes was visualized with a fluorescence microscope equipped with narrow bandpass interference-blue (NIBA) filters. The surface area of the CNV was determined after outlining the margins of the lesion defined by FITC–dextran fluorescence, using image analysis software (Photoshop; Adobe Systems, San Jose, CA).
Reverse transcription-polymerase chain reaction for detection of sflt-1 expression
RNAs were extracted from eyes 2 weeks after subretinal injection of type 8 AAV-sflt-1 or AAV-Luc. Reverse transcription-polymerase chain reaction (RT-PCR) was performed as described previously (Miyake et al., 1993), using human sflt-1-specific primers (sense, 5′-CATAGATGTCCAAATAAGCAC-3′; and antisense, 5′-CGAGTCAAATAGCGAGCAGAT-3′). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was also amplified as a control (sense, 5′-CATCACTGCCACCCAGAAGA-3′; and antisense, 5′-ATGTTCTGGGCAGCC-3′).
Quantitative analysis
Quantitative analysis of GFP-positive areas was done with a computerized image analyzer consisting of a personal computer running Lumina Vision software (Mitani, Fukui, Japan) interfaced with an Olympus digital charge-coupled device (CCD) camera (DP70) mounted on a BX60 microscope (Olympus). GFP-positive area was calculated with Photoshop software (Adobe Systems).
Statistical analysis
Morphometric data for different lesions in each eye were averaged to provide one value per eye. The mean and SD for these measures for each group was calculated and probabilities (t test) were determined by computer (Prism 3.0; GraphPad Software, San Diego, CA). p < 0.05 was considered statistically significant for all forms of statistical analysis used.
Results and Discussion
Most evaluations of ocular gene therapies have employed a type 2 AAV vector with an ocular disease model. Among these studies, three clinical trials stand out as landmarks in the development of ocular gene therapy (Bainbridge and Ali, 2008). These three trials are similar in that they all investigated the efficacy of using AAV type 2 vector-mediated gene transfer to treat Leber's congenital amaurosis, a severe inherited form of early-onset retinal degeneration (Maguire et al., 2008). However, it has been reported that more efficient gene transfer could be achieved with other types of AAV vector (Yang et al., 2002; Lebherz et al., 2008). This prompted us to compare the efficacy of ocular gene transfer in our murine CNV model, using type 5 or 8 AAV vector expressing enhanced green fluorescent protein (AAV-EGFP). We initially injected 2 μl (5 × 1011 vector genomes/ml) of type 5 or 8 AAV-EGFP into the subretina of laser photocoagulation-treated (PC+) CNV model mice and normal C57BL/6 mice (PC–) mice. Examination under a fluorescence microscope 2 weeks after the subretinal injections revealed that photoreceptors in the outer nuclear layer (ONL) and retinal pigmental epithelial (RPE) cells could be transduced with either AAV serotype, whereas ganglion cells (with cell bodies in the ganglion cell layer [GCL]) were transduced only by the type 8 AAV-EGFP vector in the absence of laser photocoagulation (Fig. 1A). We quantified the GFP-positive area in each of the layers of the retina in both the PC-treated and untreated conditions for AAV5 and AAV8 (Fig. 1B). Overall, the transduction efficiency of type 8 AAV-EGFP was significantly higher than that of type 5 AAV-EGFP. Indeed, we obtained high levels of transgene expression in the outer segment of the neural retina when using the type 8 AAV vector. Moreover, transduction efficiency was higher with laser photocoagulation than without it (Fig. 1A and B). Taken together, these findings suggest that the type 8 AAV vector is the most useful for gene transfer to retinal and choroidal tissue.
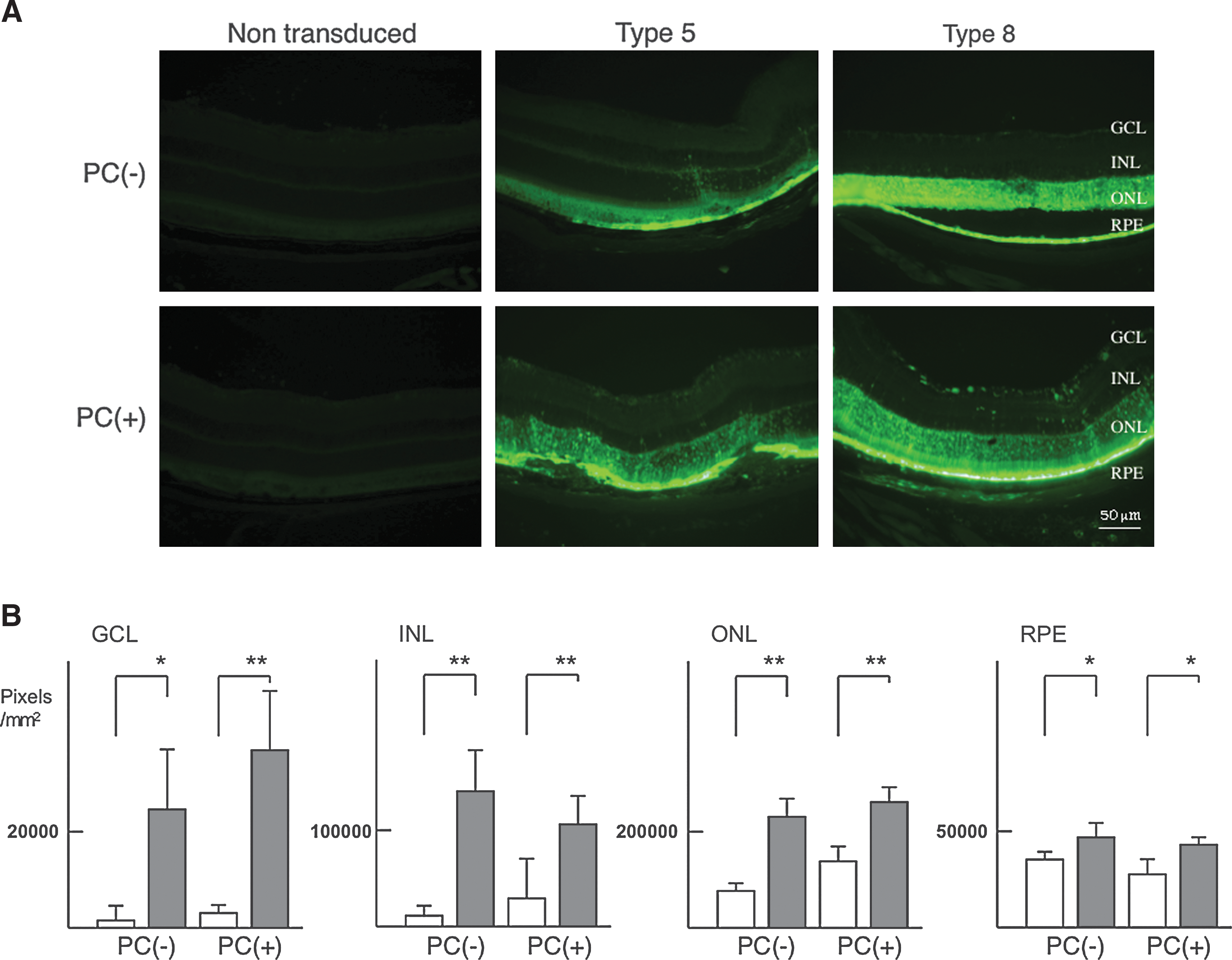
Efficient gene transfer into the subretina by type 8 AAV-EGFP. (
To treat CNV in our murine model, we generated a type 8 AAV vector (AAV-sflt-1) harboring sflt-1, which encodes a soluble, secretable form of Flt-1 that functions as an antiangiogenic effector. To investigate the biological effects of AAV 8-mediated expression of sflt-1, we initially carried out a tube formation assay using human umbilical vein endothelial cells (HUVECs) cocultured with fibroblasts. When HUVECs were cultured in the presence of VEGF, efficient tube formation was observed (Fig. 2A), and no significant changes were elicited by treatment with supernatant from AAV-EGFP-transduced C2C12 cells (Fig. 2B). By contrast, when supernatant from AAV-sflt-1-transduced C2C12 cells was added, tube formation was significantly inhibited (Fig. 2C). Determination of tube area and length confirmed that AAV-sflt-1 efficiently inhibited in vitro angiogenesis (area: 25,121 ± 557 vs. 68,628 ± 1357 pixels; p < 0.01; length: 4811 ± 246 vs. 10,894 ± 297 pixels; p < 0.01) (Fig. 2D and E). Thus AAV-sflt-1 appears to effectively mediate the expression and secretion of functional soluble Flt-1.

Suppression of tube formation by treatment with supernatant conditioned by type 8 AAV-sflt-1-transduced cells. HUVECs cocultured with human fibroblasts were treated with 100 μl of supernatant from AAV-sflt-1-transduced cells (
We next investigated the efficacy of AAV-sflt-1 in a murine laser photocoagulation-induced CNV model. After injection of 2 μl (5 × 1011 vector genomes/ml) of type 8 AAV-slft-1 or a control type 8 AAV vector harboring the lacZ gene (AAV-LacZ), the ability to inhibit angiogenesis was analyzed by the choroidal flatmount method, and leakage during fluorescein angiography was analyzed as previously described (Mori et al., 2002). We found that leakage due to CNV was inhibited significantly in mice treated with AAV-sflt-1 (Fig. 3B), as compared with mice treated with AAV-LacZ (Fig. 3A) (555 ± 304 vs. 1470 ± 1000 μm2; p = 0.007) (Fig. 3C). Expression of sflt-1 was detected in AAV-sflt-1-injected mice but not in noninjected and AAV-LacZ-injected mice (Fig. 3D). Thus subretinal injection of type 8 AAV-sflt-1 was an effective treatment for CNV in our murine model.
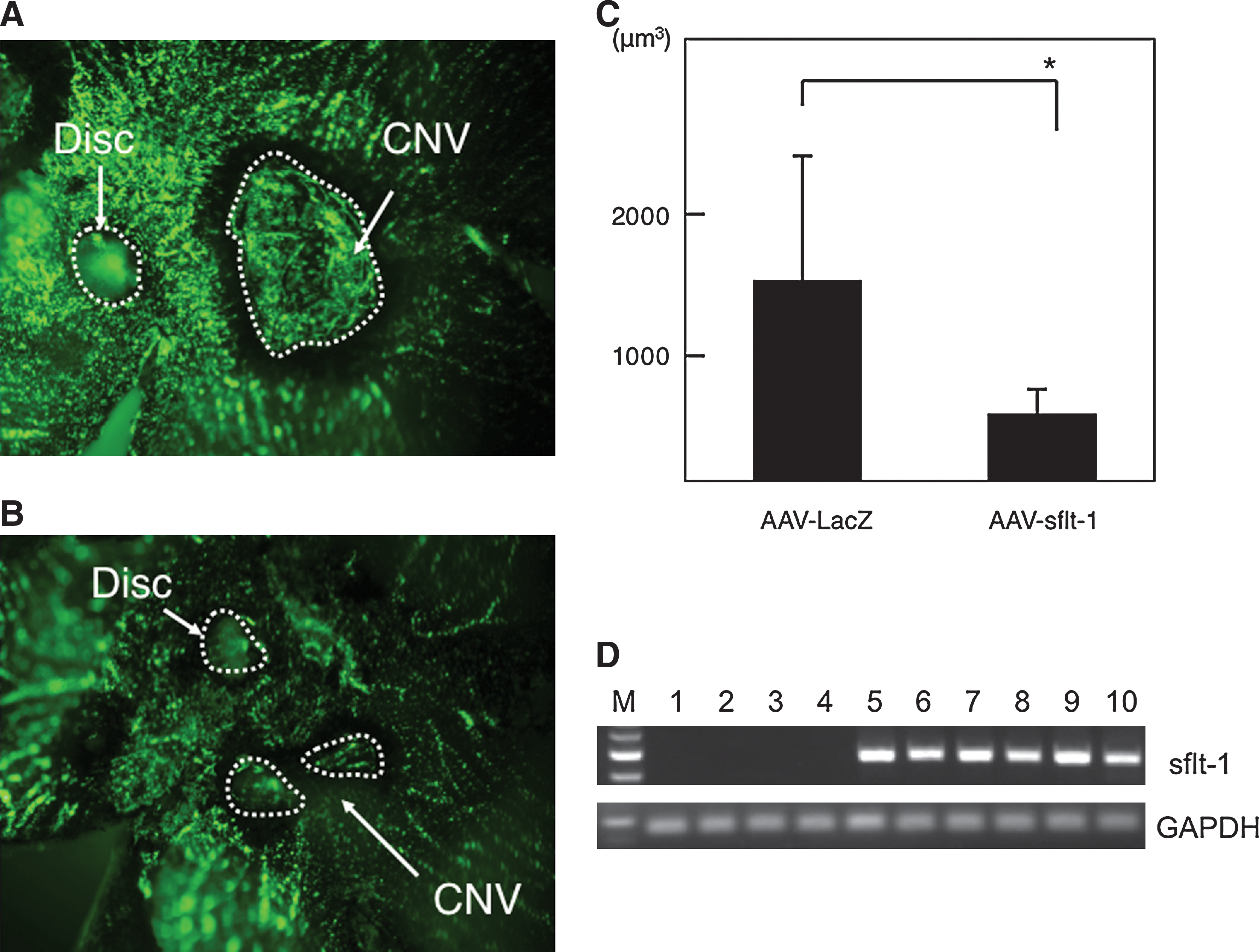
Efficient suppression of CNV in vivo. Two weeks after type 8 AAV-sflt-1 (n = 11) or AAV-LacZ (n = 12) injection, CNVs of the eyes were analyzed by fluorescein angiography. CNV lesions in eyes injected with AAV-sflt-1 (
A number of gene therapeutic approaches have been applied to inhibit ocular angiogenesis, and choroidal neovascularization has been inhibited by viral vectors mediating expression of angiostatin (Igarashi et al., 2003), endostatin (Mori et al., 2001), or pigment epithelium-derived factor (PEDF) (Mori et al., 2002). In particular, angiostatin, which we previously showed to inhibit retinal neovascularization, will be tested in a lentiviral vector system in a phase I clinical trial in 2009 (
There have been several experiments in which sflt-1 was transduced to treat ocular neovascularization (Honda et al., 2000; Lai et al., 2002, 2005; Gehlbach et al., 2003; Rota et al., 2004). Of those, sflt-1 transduced by adenoviral vectors (Gehlbach et al., 2003) and type 2 AAV vectors (Lai et al., 2002, 2005) inhibited choroidal neovascularization. In the present study, we demonstrated that a type 8 AAV vector could be efficiently transduced into the subretina and could consistently control angiogenesis in an experimental model of retinal neovascularization. The high efficiency of type 8 AAV-mediated transduction could reduce the dose of AAV vector needed, resulting in a safer and less expensive vector for use in a clinical trial. In summary, our findings demonstrate the feasibility of an in vivo gene therapeutic approach to the clinical management of CNV in conditions such as AMD.
Footnotes
Acknowledgments
This work was supported in part by grants from the Ministry of Health and Welfare of Japan and the Ministry of Education, Science, and Culture of Japan. The authors thank Dr. James Wilson at the University of Pennsylvania for providing AAV packaging plasmids (pacH, and p5E18-VD2/8). The authors also thank Yukihiko Hirai and Yuko Odagaki for making AAV viral vectors, and Nagisa Asakawa for technical assistance.
Author Disclosure Statement
No competing financial interests exist.