
Abstract
Our studies and those of many others have implicated hepatocyte necrosis and apoptosis mediated by fibrinogen-like protein-2 (fgl2) prothrombinase and tumor necrosis factor receptor (TNFR) in the development of fulminant viral hepatitis, a disease with a mortality rate greater than 80% in cases lacking immediate organ transplantation. This study was designed to explore the efficacy of dual short hairpin RNA (shRNA) interference with fgl2 and TNFR1 in the treatment of murine hepatitis virus strain 3 (MHV-3)-induced fulminant hepatitis in mice. Plasmids p-mfgl2shRNA and p-mTNFR1shRNA, complementary to the sequences for mfgl2 and mTNFR1, were constructed. Plasmids pEGFP-mfgl2 and pEGFP-mTNFR1 expressing mfgl2–EGFP (enhanced green fluorescent protein) and mTNFR1–EGFP fusion proteins were also constructed to screen the inhibitory effect of p-mfgl2shRNA and p-mTNFR1shRNA on mfgl2 and mTNFR1 expression. Cotransfection of individual shRNA plasmids and pcDNA3.0-mfgl2 and pcDNA3.0-mTNFR1 expression constructs into Chinese hamster ovary (CHO) cells significantly inhibited mfgl2 and mTNFR1 gene expression, as evidenced by fluorescence microscopy, reverse transcription-polymerase chain reaction, and Western blotting. In vivo hydrodynamic delivery of dual-interference shRNA plasmids for mfgl2 and mTNFR1 significantly decreased mfgl2 and mTNFR1 expression; markedly ameliorated fibrin deposition, hepatocyte necrosis, and apoptosis; and prolonged survival against fulminant viral hepatitis induced by MHV-3 in BALB/cJ mice compared with mfgl2 or TNFR1 single-gene interference. These results indicate that in vivo interference with genes for more than one key target provides superior treatment efficacy compared with single-gene interference.
Introduction
RNA interference (RNAi), an evolutionarily conserved gene-silencing mechanism present in a variety of eukaryotic species, has proven to be an extremely potent and versatile tool for specifically decreasing the expression of targeted genes (Ma et al., 2007). The shRNA (small hairpin RNA) is a type of small interfering RNA (siRNA). RNAi uses short, double-stranded RNA to trigger the degradation or translational repression of homologous RNA targets in a sequence-specific manner (Khvorova et al., 2003). To date, RNAi has been used as a novel and effective tool in functional genomics studies and has displayed great potential in treating human diseases, including human genetic and acquired disorders such as cancer and viral infection. Research reports have also demonstrated the potential for use of siRNAs as therapeutic agents. The siRNA method can be surprisingly potent; for example, transfections performed with subnanomolar concentrations of RNA have been reported to achieve a 90% reduction in mRNA level (Kim et al., 2005).
Results from our previous study showed that an mfgl2 antisense plasmid inhibited mfgl2 expression and ameliorated MHV-3-induced fulminant hepatitis in BALB/cJ mice (Zhu et al., 2006). In the present study, we constructed mfgl2 and mTNFR1 shRNA expression plasmids to investigate the effect of dual interference with both targets on liver disease in the same animal model.
Materials and Methods
Virus
The MHV-3 virus was obtained from the American Type Culture Collection (ATCC, Manassas, VA), plaque-purified on monolayers of delayed brain tumor cells, and titered on L2 cells according to a standard plaque assay.
Animals
All animal experiments were carried out according to the guidelines of the Chinese Council on Animal Care and approved by the Tongji Hospital (Tongji Medical College, Huazhong University of Science and Technology, Wuhan, China) Committees on Animal Experimentation. The research protocol was reviewed and approved by the institutional review board of Tongji Hospital. Female BALB/cJ mice (6–8 weeks of age, 18–20 g) were purchased from the Hubei Provincial Institute of Science and Technology (Wuhan, China). Gene transfection was achieved by hydrodynamic injection over 4 to 5 sec, via the tail vein, of 100 μg of p-mfgl2shRNA or p-mTNFR1shRNA or control plasmid, dissolved in 2 ml of normal saline (18 animals in each group). The injection was repeated 24 hr later. In the intervening period, mice received 20 plaque-forming units of MHV-3 intraperitoneally to induce fulminant viral hepatitis. Liver sections and serum samples from separate sets of animals were sampled at the hours indicated after MHV-3 infection.
Creation of fgl2 and TNFR1 eukaryotic expression plasmids and GFP fusion expression plasmids
Mouse genomic DNA was extracted from mouse liver and used as a template for polymerase chain reaction (PCR) amplification to obtain complete genes for mfgl2 and mTNFR1. Two pairs of primer sets, f1/f2 and T1/T2 (f1, 5′-TTT GGG GCA TGG ACA TTG AC-3′; f2, 5′-CCC ACC TTA TGA GTC CAA GG-3′; T1, 5′-TTA ATT AAG CTT ATG GGT CTC CCC ACC GTG-3′; and T2, 5′-ATA TAT GGA TCC TCG CGG GAG GCG GGT CGT-3′), were used to amplify 1300 bp of the mfgl2 gene and 1365 bp of the mTNFR1 gene, respectively, and the complete genes for mfgl2 and mTNFR1 were cloned into the HindIII/BamHI (for mfgl2) or HindIII/KpnI (for mTNFR1) sites of the PMD18-T cloning vector (Takara, Tokyo, Japan), and the sequences were confirmed by sequence analysis. Purified DNA fragments were subcloned into the pcDNA3.0(+) vector (Invitrogen, Carlsbad, CA) at the restriction sites mentioned previously to construct the expression plasmids pcDNA3.0-mfgl2 and pcDNA3.0-mTNFR1. With the use of the EGFP-N2 vector, mfgl2-GFP and mTNFR1-GFP fusion plasmids were also constructed (pEGFP-mfgl2 and pEGFP-mTNFR1).
Construction of mfgl2 and mTNFR1 shRNA plasmids
Interference shRNA plasmids for mfgl2 and mTNFR1 were constructed with the retroviral vector pMSCVneo. The shRNA sequences of the target genes were as follows: mfgl2 shRNA, 5′-GCA GTG GAC AGT CTG AAG A-3′; and mTNFR1 shRNA, 5′-GGA AAG TAT GTC CAT TCT A-3′. The mouse U6 promoter was amplified from mouse genomic DNA, and the U6 promoter upstream of the hairpin sequences of the target genes was cloned into pMSCVneo to form shRNA-interfering plasmids p-mfgl2shRNA and p-mTNFR1shRNA. In the same vector, shRNA plasmids containing the same lengths of irrelevant sequence were constructed as controls. Sequences for the mfgl2 and mTNFR1 shRNA plasmids were also verified to be correct.
Transient transfection
The 1.3- and 1.36-kb fragments of complete mfgl2 and mTNFR1 cDNA were, respectively, cloned into pEGFP-N2 (Clontech, Mountain View, CA) upstream of the GFP gene at restriction sites HindIII/BamHI for mfgl2 and HindIII/KpnI for mTNFR1. The Chinese hamster ovary (CHO) cell line was cultured in 6-well plates to 90–95% confluence. The shRNA plasmid or irrelevant shRNA plasmid, along with pEGFP-mfgl2 and pEGFP-mTNFR1 in serum-free F12-Dulbecco's modified Eagle's medium (DMEM), was mixed with Lipofectamine 2000 (2 μg/μl; Invitrogen) according to the manufacturer's protocol. After incubation at room temperature for 20 min, this mixture was distributed into duplicate wells of CHO cells, and transfection was performed at 37°C in 5% CO2. The medium was replaced with fresh complete medium 5–6 hr after transfection. At 48 hr after transfection, direct observation of the effect of shRNA on the target genes was performed by fluorescence microscopy. Cells were subsequently harvested for reverse-transcription (RT)-PCR and Western blotting.
Western blot analysis
For Western blotting, protein extracts from CHO cells 48 hr after transfection were lysed in lysis buffer containing 150 mM NaCl, 50 mM Tris (pH 7.4), 5 mM ethylenediaminetetraacetic acid (EDTA), 1% Nonidet P-40, and protease inhibitors for 30 min on ice, and whole-cell lysates were obtained by centrifugation. Samples were boiled for 5 min and cooled on ice. Proteins were subjected to 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE) and transferred to nitrocellulose membrane. Membranes were blocked in 5% nonfat milk and 0.05% Tween 20 in phosphate-buffered saline (PBS) for 2 hr at room temperature with shaking, and then incubated with polyclonal antibody against fgl2 (diluted 1:500; Zhu et al., 2006) and specific monoclonal antibody against TNFR1 (diluted 1:200; Abzoom, Wuppertal, Germany). Membranes were rinsed twice in PBS and washed three times in 0.05% Tween 20 in PBS for 5 min, with shaking. Membranes were then incubated with horseradish peroxidase-labeled goat anti-rabbit secondary antibody (Santa Cruz Biotechnology, Santa Cruz, CA) for 1 hr, with shaking, before being rinsed twice in PBS and washed three times in 0.05% Tween 20 in PBS for 5 min.
Real-time fluorescence quantitative PCR
Total RNA was extracted from mouse liver with TRIzol reagent (Invitrogen) and reverse-transcribed into cDNA (Fermentas, Burlington, ON, Canada) according to the manufacturer's standard protocol. Real-time fluorescence quantitative PCR was performed with PCR mix (Toyobo, Tokyo, Japan) according to the manufacturer's standard protocol to detect mfgl2 and mTNFR1 expression at the mRNA level. The upstream primer for mfgl2 was 5′-ACT GTG ACA TGG AGA CCA TG-3′, and the downstream primer was 5′-TCC TTA CTC TTG GTC AGA AG-3′. The upstream primer for mTNFR1 was 5′-GGT CTT TGC CTT CTA TCC TT-3′, and the downstream primer was 5′-TTC CAG CCT TCT CCT CTT T-3′. The upstream primer for glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was 5′-CCTGGAGAAACCTGCCAAGTATGA-3′, and the downstream primer was 5′-GGTGGAAGAGTGGGAGTTGCTGT-3′.
Measurement of serum alanine aminotransferase
Plasma or serum alanine aminotransferase (ALT) level was measured with a clinical chemistry analyzer (Aeroset system; Abbott Laboratories, Abbott Park, IL).
Immunohistochemical staining
Immunohistochemical staining was performed with a rabbit polyclonal antibody generated against fgl2 (diluted 1:200) and a monoclonal antibody generated against TNFR1 (diluted 1:100; Abzoom). Liver slices were blocked in 10% normal goat serum in PBS at room temperature for 2 hr. The slices were incubated with primary antibody at room temperature for 2 hr. Subsequently, the slices were incubated with immunoperoxidase-conjugated goat IgG fraction to rabbit IgG Fc (Dako, Glostrup, Denmark) at room temperature for 40 min, followed by three washes in PBS containing 0.05% Tween 20. Slices were then air dried, visualized with a microscope, and photographed.
Detection of hepatocellular apoptosis
The terminal deoxynucleotidyltransferase (TdT)-mediated dUTP nick-end labeling (TUNEL) method (TUNEL kit; Bosted, Wuhan, China) was used to detect apoptosis in hepatocytes. Liver slices were dewaxed and then treated with 3% H2O2 and subjected to proteinase K digestion. TdT and digoxin–dUTP were added overnight at 4°C, followed by incubation for 30 min with biotinylated anti-digoxin antibody at 37°C and then binding with streptavidin–peroxidase. The slices were then washed with PBS, and color development was subsequently carried out with diaminobenzidine (DAB). Hepatocytes with condensed brown particles within the nucleus were considered to have undergone apoptosis. The apoptotic index (AI) was calculated on visualization by light microscopy: The number of apoptotic cells in 10 randomly selected high-power fields was determined, and the AI was calculated as (apoptotic cell count/total cell number) × 100%.
Statistical analysis
Quantitative data are expressed as means ± standard deviation. The Student t test for unpaired samples (two-tailed) was used to analyze the data with SPSS 12 statistical software (SPSS, Chicago, IL), and a p value less than 0.05 was considered statistically significant.
Results
Construction of mfgl2 and mTNFR1 expression plasmids, GFP fusion expression plasmids, and corresponding shRNA plasmids
The mfgl2 and mTNFR1 eukaryotic expression plasmids and GFP fusion expression plasmids were successfully constructed as evidenced by restriction enzyme mapping with HindIII/BamHI for mfgl2 and HindIII/KpnI for mTNFR1 (Fig. 1A) and confirmed by sequence analysis. The corresponding shRNA plasmids p-mfgl2shRNA and p-mTNFR1shRNA were also constructed (Fig. 1B).
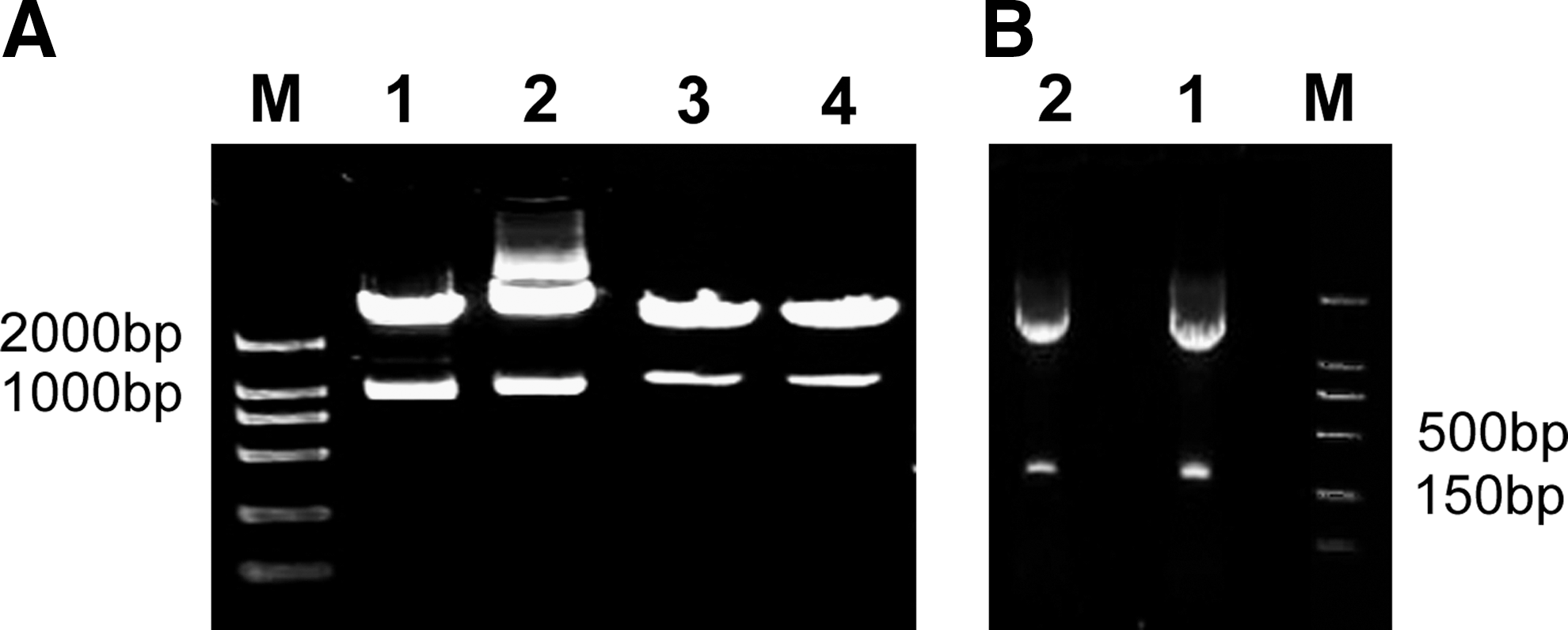
Construction of mfgl2 and mTNFR1 eukaryotic expression plasmids, GFP fusion expression plasmids, and shRNA plasmids. The expression vectors were successfully constructed as evidenced by restriction enzyme mapping and confirmed by sequence analysis. (
shRNA plasmids inhibited expression of mfgl2 and mTNFR1 in CHO cells
The numbers of green fluorescent cells and their fluorescence intensity in CHO cultures treated with shRNA were significantly decreased 48 hr after cotransfection compared with those of cultures treated with irrelevant shRNA plasmid or with no treatment (Fig. 2A). The shRNA plasmids significantly inhibited the expression of mfgl2 and mTNFR1 not only at the mRNA level, as determined by RT-PCR (Fig. 2B), but also at the protein level, as assessed by Western blot (Fig. 2C).
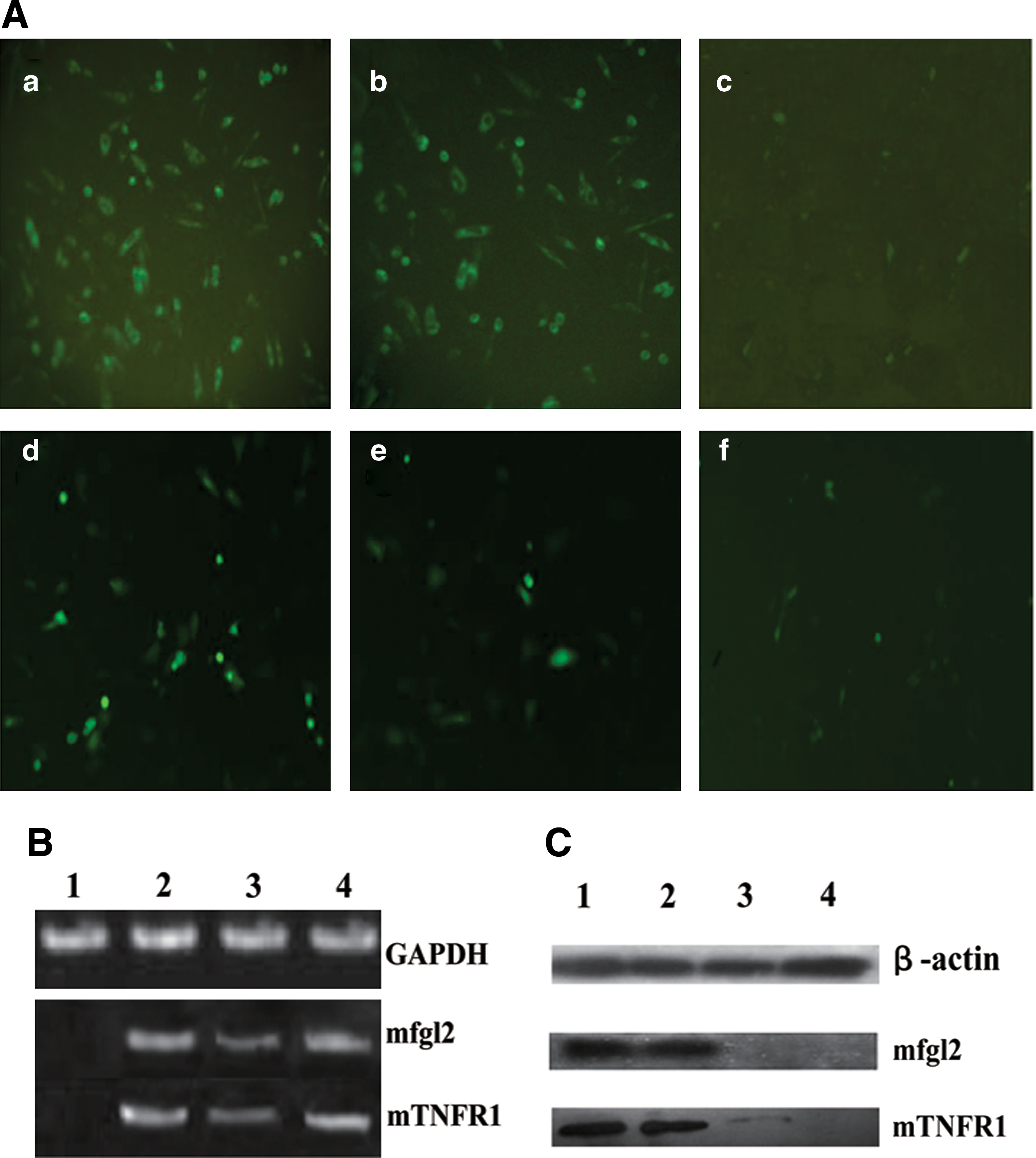
mfgl2 and mTNFR1 shRNA plasmids significantly inhibited target gene expression in CHO cells. (
Dual interference with mfgl2 and TNFR1 resulted in increased survival of mice with MHV-3-induced fulminant viral hepatitis
To investigate the biological effect of p-mfgl2shRNA and p-mTNFR1shRNA plasmids in vivo, shRNA plasmids were delivered twice by hydrodynamic injection into 18 MHV-3-infected BALB/cJ mice. In the p-mfgl2shRNA and p-mTNFR1shRNA dual interference group, all 18 mice with shRNA plasmids were alive on day 3 after infection, and 6 of 18 mice (33.3%) recovered from fulminant viral hepatitis. Two of 18 mice (11.1%) recovered with p-TNFR1shRNA treatment alone, and 2 of 18 (11.1%) mice with p-mfgl2shRNA plasmid were alive on day 7 after infection. In contrast, no mice in the control plasmid-treated group survived to day 4 (Fig. 3).
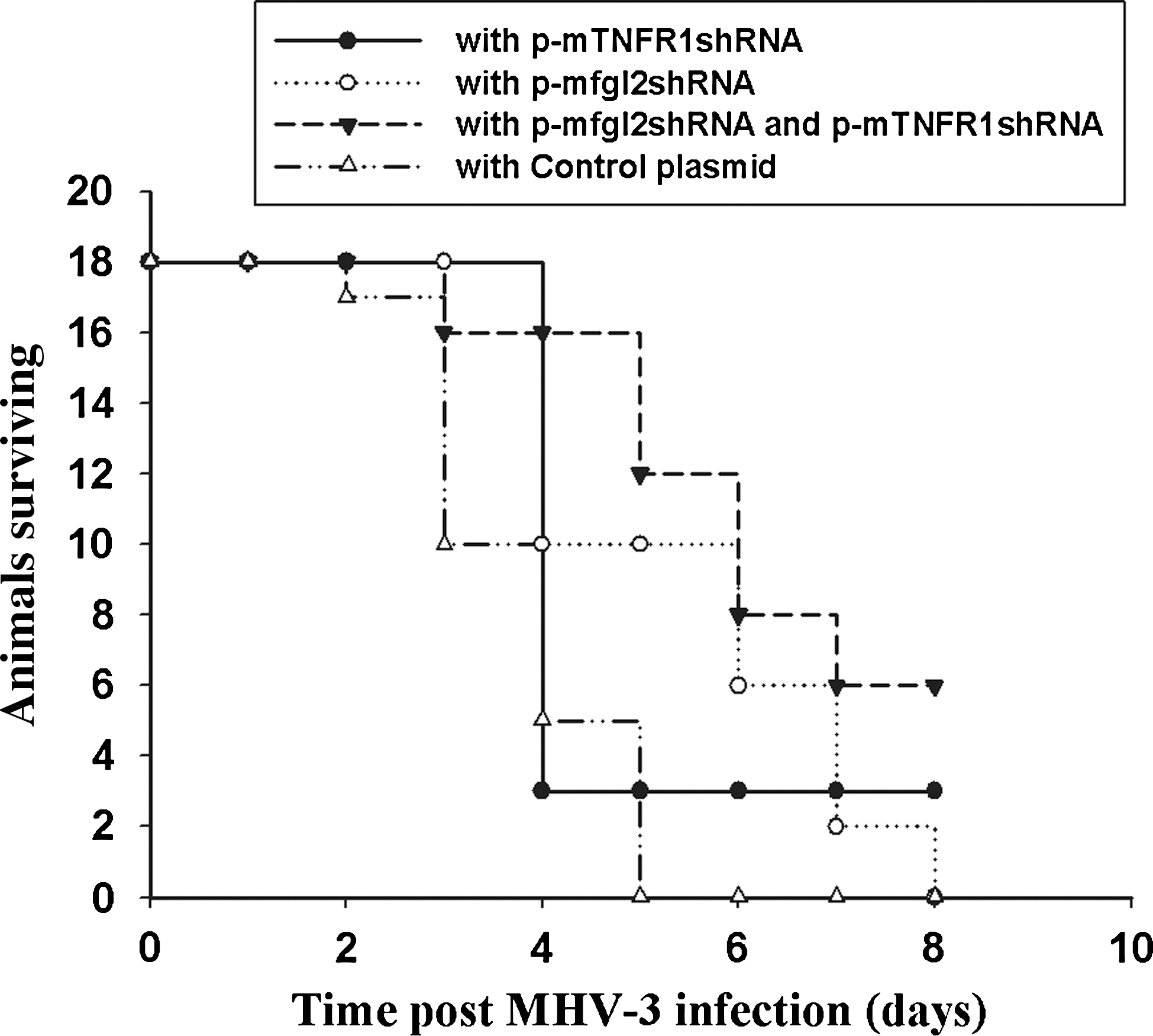
Dual interference with mfgl2 and TNFR1 increased the survival of MHV-3-infected BALB/cJ mice. mfgl2 and/or mTNFR1 shRNA plasmids or control plasmid were introduced into BALB/cJ mice and the mice were then infected as described in Materials and Methods. Survival data are presented.
Dual interference with mfgl2 and TNFR1 resulted in improved liver function and histological recovery in mice with MHV-3-induced fulminant viral hepatitis
To better understand the mechanism of the biological effects of p-mfgl2shRNA and p-mTNFR1shRNA dual interference, a time course study of liver function (serum ALT level) was performed, and liver histology in MHV-3-infected mice was evaluated. The serum ALT level started to increase 24 hr after infection in both the p-mfgl2shRNA- and p-mTNFR1shRNA-treated groups and in the control group. However, by 48–60 hr after infection ALT levels had significantly decreased in targeted gene interference mice, particularly in dual shRNA-treated mice compared with the control plasmid group (Fig. 4A). Liver histology was examined by hematoxylin and eosin (H&E) staining at the indicated times. By 60 hr after MHV-3 infection, only mild inflammatory cell infiltration, with no evidence of hepatocyte necrosis, was found in dual p-mfgl2shRNA- and p-mTNFR1shRNA-treated mice (Fig. 4B, panel b) compared with the focal, evenly distributed necrosis and massive inflammatory cell infiltration in control plasmid-treated mice (Fig. 4B, panel a). Single-gene interference with p-mfgl2shRNA or p-mTNFR1shRNA also markedly ameliorated hepatic inflammatory infiltration (Fig. 4B, panels c and d). These data strongly suggested that dual interference with p-mfgl2shRNA and p-mTNFR1shRNA significantly improved liver function and pathology in MHV-3-induced fulminant viral hepatitis.
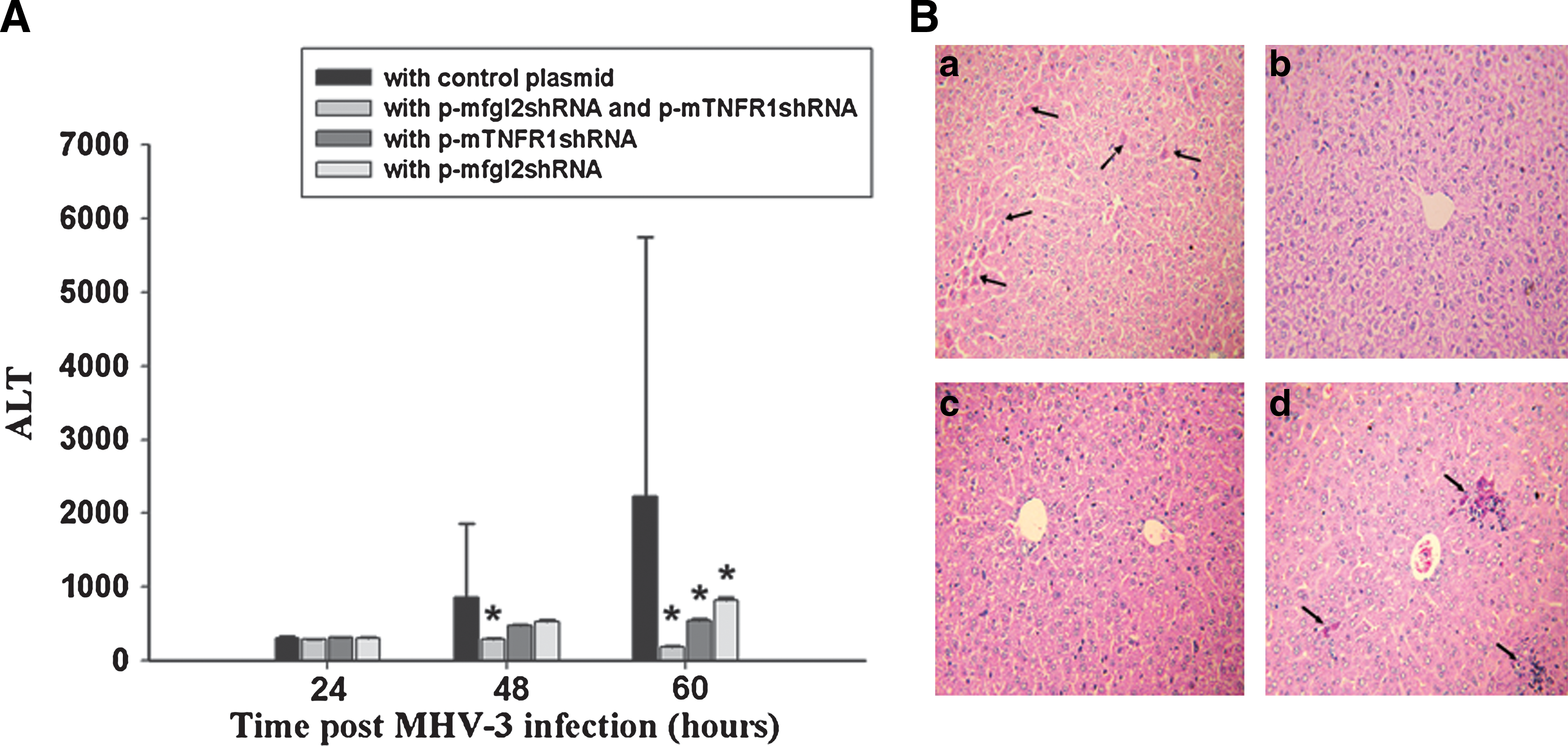
Dual interference with mfgl2 and TNFR1 improved liver function and histopathology in MHV-3-infected BALB/cJ mice. Each group of shRNA plasmids was dissolved in 2 ml of PBS and introduced into BALB/cJ mice by hydrodynamic injection twice. Mice then received 20 PFU of MHV-3 intraperitoneally to develop fulminant viral hepatitis. Serial serum ALT levels (
Effect of p-mfgl2shRNA and p-mTNFR1shRNA on target gene expression and related fibrin deposition and intrahepatic apoptosis in MHV-3-infected mice
To evaluate the inhibitory effects of p-mfgl2shRNA on mfgl2 expression and of p-mTNFR1shRNA on mTNFR1 expression in vivo, a time course study was performed by real-time fluorescence quantitative PCR, using liver samples from MHV-3-infected mice 24, 48, and 60 hr after MHV-3 infection. The inhibitory effect of the targeted gene shRNA plasmids began 24 hr after MHV-3 infection, with a maximal inhibitory effect 60 hr after infection (Fig. 5A and B). These results were confirmed by immunohistochemistry with a specific polyclonal antibody against mfgl2 and a monoclonal antibody against mTNFR1. Little hepatic mfgl2 expression in p-mfgl2shRNA-treated mice (Fig. 5D), mTNFR1 expression in p-mTNFR1shRNA-treated mice (Fig. 5G), and no mfgl2 or mTNFR1 expression in dual interference (Fig. 5E and H) was evidenced, compared with mice treated with control plasmid (Fig. 5C and F). There were significant decreases in fibrin deposition (Fig. 5I–K) and hepatocellular apoptosis (Fig. 5L–N) in liver tissue from mice treated with p-mfgl2shRNA (Fig. 5J) and p-mTNFR1shRNA (Fig. 5M) or dual interference with both shRNA (Fig. 5K and N) compared with that in mice treated with control plasmid. There was a significant decrease in apoptosis in liver from mice treated with p-mTNFR1shRNA (72 hr postinfection) or with both p-mTNFR1shRNA and p-mfgl2shRNA (48 hr postinfection), as shown by the apoptosis index (AI) (Fig. 5O).
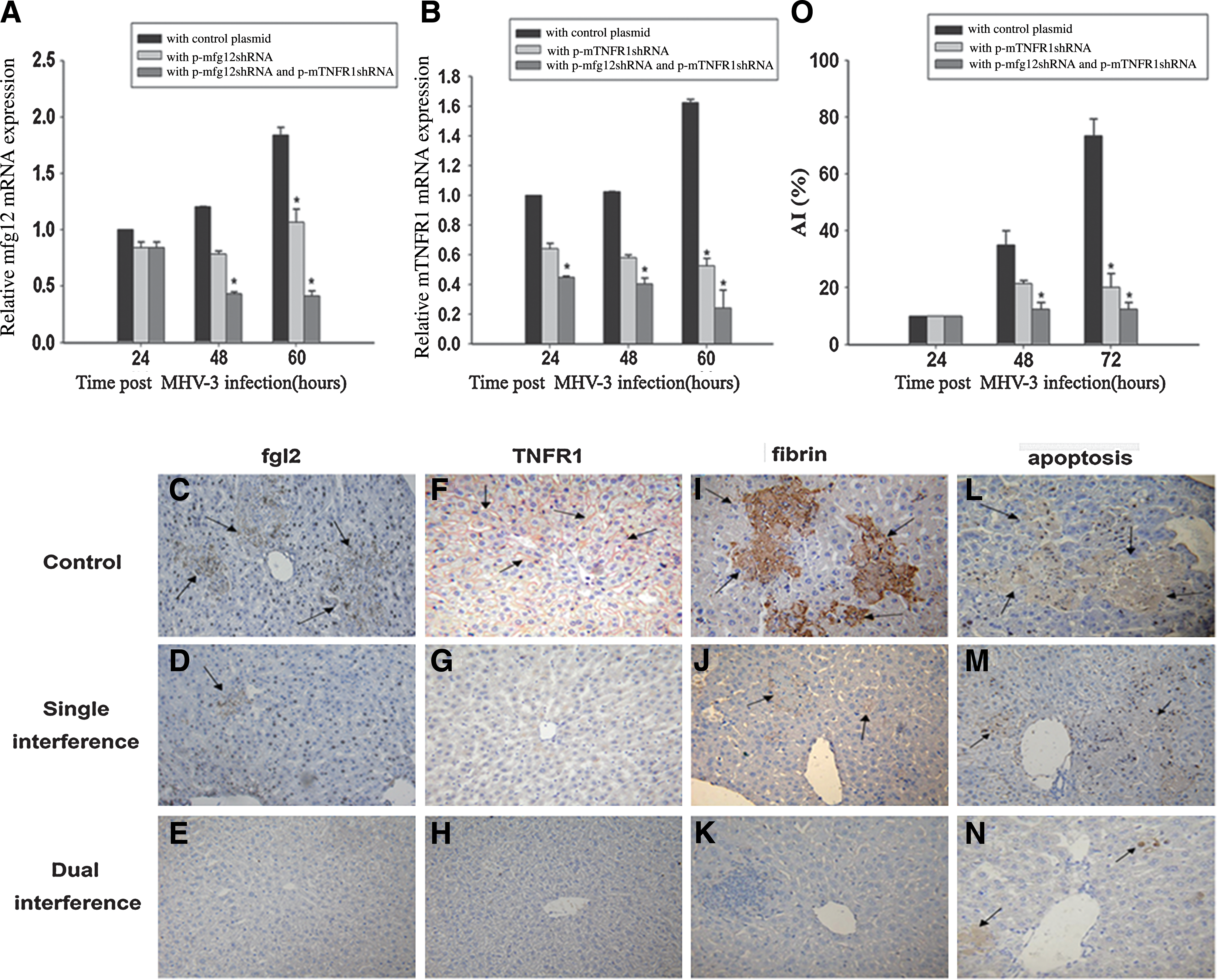
mfgl2shRNA and mTNFR1shRNA plasmids inhibited target gene expression remarkably and decreased related fibrin deposition and intrahepatic apoptosis in MHV-3-infected BALB/cJ mice. Livers from BALB/cJ mice treated with mfgl2shRNA and/or mTNFR1shRNA plasmid and from control plasmid-treated BALB/cJ mice were collected 60 hr after MHV-3 infection, and targeted gene mfgl2 and TNFR1 expression was determined by (
Discussion
Approximately 400 million individuals worldwide have chronic HBV infection, particularly in Asia, where the chronic carrier rate is 10–20% (Pawlotsky, 2002; Cheng et al., 2003). The mortality from fulminant viral hepatitis is greater than 80% in cases lacking immediate liver transplantation (Coleman, 2003; Higuchi and Gores, 2003; Nakamoto and Kaneko, 2003). Therefore, investigation of the mechanism of hepatocyte death in severe hepatitis and the development of gene therapeutic means to block the pathological process have become serious issues in this field. Our studies and those of many others have implicated fgl2 prothrombinase and TNF-α-mediated hepatocyte necrosis and apoptosis in a wide range of liver diseases including viral hepatitis, alcoholic hepatitis, ischemia/reperfusion injury, fulminant hepatic failure, and cancer.
A number of studies have suggested that fgl2 plays a pivotal role in the pathogenesis of experimental and clinical severe viral hepatitis (Li et al., 1992; Ding et al., 1998; Levy et al., 2000; Marsden et al., 2003). Early reports demonstrated hepatic fibrin deposition in patients with chronic hepatitis. Our previous reports in an animal model of fulminant viral hepatitis induced by MHV-3 and in patients with acute or chronic HBV infection have shown a critical role for fgl2 prothrombinase in the pathogenesis of fulminant viral hepatitis. Taking into consideration these results, together with those of many others, we propose that fgl2 may offer an important new therapeutic approach for hepatitis virus-induced disease.
The liver is particularly susceptible to TNF-α-mediated cytotoxicity. The TNF-induced apoptotic death of hepatocytes may also be of major importance under conditions of acute liver failure in septic shock (Leist et al.., 1994). In fact, hepatic failure and tissue destruction as a result of endogenously produced TNF have been demonstrated to be mediated by the 55-kDa TNF receptor (TNFR1) (Pfeffer et al., 1993). Hepatocyte apoptosis in the liver plays an important role in the processes of many liver diseases, particularly hepatic failure resulting from various causes. In the process of severe hepatitis, activation of Fas and TNF affects the severity of hepatocyte apoptosis (Ogasewara et al., 1993; Pinkoski et al., 2000). In addition, lipopolysaccharide (LPS) toxicity in response to severe apoptosis induces further liver injury and destruction in the fulminant hepatic failure model induced by LPS/
There are currently no safe and effective antiviral treatments for fulminant viral hepatitis. However, there has been significant interest in the use of gene therapy, which can be applied to evaluate the role and importance of target genes in the pathogenesis of disease (Stevenson, 2004). High levels of gene transfer in mouse liver can be achieved by hydrodynamic injection of a large volume of plasmid DNA solution into the tail vein. This technology has been used extensively in gene therapy for mouse studies (Zhu et al., 2006). There has been a focus on the use of gene therapy in fulminant viral hepatitis. For example, Nakayama and colleagues used adenovirus encoding immunoglobulin against cytotoxic T lymphocyte antigen (CTLA)-4 to suppress liver injury by inhibiting acquired immune responses in a mouse model of fulminant hepatitis induced by injection of Propionibacterium acnes and LPS (Nakayama et al., 2005). In addition, Arvelo and colleagues reported that adenovirus-mediated hepatic expression of A20, an antiapoptotic protein, protected BALB/cJ mice from
We constructed an mfgl2 antisense plasmid and found that therapeutic effects are achievable, including significantly ameliorated inflammatory infiltration, fibrin deposition, and hepatocyte necrosis; prolonged survival time; and increased survival rate of BALB/cJ mice with MHV-3-induced fulminant hepatitis (Zhu et al., 2006). It is frequently questioned whether RNAi has significant advantages over antisense technology. It has been considered that if a sufficient amount of antisense DNA can be delivered to the appropriate subcellular compartment, antisense inhibition of gene expression can be as potent as that by RNAi. However, antisense technology has failed to gain widespread acceptance as a gene knockdown tool, owing largely to the extensive testing that is often required to identify effective target sites for the gene of interest (McCaffrey et al., 2003). It is currently possible to obtain potent RNAi reagents without the need for extensive testing (Khvorova et al., 2003; Amarzguioui and Prydz, 2004). Given that RNAi is thought to trigger the degradation of target mRNA before translation (Cordeiro et al., 2003), it is possible that RNAi could act synergistically to specifically result in more efficient gene silencing.
There are few studies targeting more than one molecule. Heering and colleagues reported that simultaneous loss of the DLC1 (deleted in liver cancer-1) and PTEN (phosphatase and tensin homolog) tumor suppressors enhanced breast cancer cell migration (Heering et al., 2009). In the present study, we used a combination RNAi methodology employed to modulate fgl2 and TNFR1 expression as a novel potential therapeutic strategy for fulminant viral hepatitis. Simultaneous shRNA interference with fgl2 and TNFR1 showed better results than shRNA interference with either target alone. The mechanisms involved in dual interference are not clear. Previous studies have shown that TNF-α induced fgl2 expression in endothelial cells in vitro. In addition, TNF-α enhanced IFN-γ-induced macrophage fgl2 transcription, and in vivo, TNF-α induced hepatocyte apoptosis, which is dependent on induction of fgl2 (Liu et al., 2006). Therefore, dual interference may involve synergistic effects on two key genes in the pathogenesis of this disease. However, although not the focus of this study, the issue concerning whether blocking apoptosis leads to enhanced viral replication and pathogenicity in mice needs to be further explored as a previous study has demonstrated that caspase inhibitors blocked MHV-3-induced apoptosis and enhanced viral replication and pathogenicity. Caspase inhibitors block MHV-3-induced apoptosis and enhance viral replication and pathogenicity (Leibowitz and Belyavskaya, 2001).
In the present study, we constructed shRNAs complementary to the codon sequences and showed significant inhibitory effects on mfgl2 and mTNFR1 expression in vitro and in vivo. To test the specificity of the 370-bp shRNA plasmids, two additional reporter constructs were used. The first was pEGFP, which, like the basic plasmid pcDNA3.0, encodes GFP under the control of the cytomegalovirus promoter. The second construct was an irrelevant shRNA plasmid. These two constructs failed to inhibit gene expression at the level of either transcription or translation. These data provide additional evidence of the specificity of the shRNA plasmids.
Considering the complexity of fulminant viral hepatitis, treatment will be required to address multiple disorders, including enhancement or suppression of immune responses by cytokine administration or increasing hepatic regeneration. In our present study, the overall survival rate in mice receiving dual mfgl2 and mTNFR1 interference was superior to that of mice receiving single-target interference (33.33 vs. 11.1%).
In summary, we have demonstrated that novel, specific shRNA plasmids can be used successfully to block the expression of mfgl2 and mTNFR1 in a cell culture system and in an animal model. This method increased the survival period as well as the survival rate in MHV-3-infected BALB/cJ mice, which normally die of fulminant hepatitis, and ameliorated liver pathology including inflammatory cell infiltration, fibrin deposition, and hepatocellular apoptosis. We further showed that dual interference with mfgl2 and TNFR1 showed superior efficacy compared with single-gene interference. Results of this study may provide an effective therapeutic strategy for targeted intervention in diseases involving fgl2 and TNFR1.
Footnotes
Acknowledgments
This work was supported by the National Key Basic Research Program of China (2007CB512900, 2005CB522901, 2005CB52250), the National Science Fund of China (NSFC30672380), and the National Science Youth Fund of China (NSFC30700702).
Author Disclosure Statement
No competing financial interests exist.