
Abstract
Hepatocellular carcinoma (HCC) is one of the most common cancers worldwide, but effective therapies are still needed. The liver has been identified as an important immune organ and is heavily populated with various lymphocyte subsets known to play important roles in cancer immunosurveillance. We hypothesized that activation of hepatic lymphocytes by interleukin (IL)-15, a cytokine known for its ability to trigger proliferation and activation of natural killer (NK) cells, natural killer T cells, and memory CD8+ T cells, might offer an alternative therapy for HCC. We employed hepatotropic adeno-associated virus serotype 8 (AAV8) to deliver an IL-15 superagonist (IL-15-IL-15RαS), consisting of IL-15 covalently linked to the N-terminal sushi domain of the IL-15 receptor α chain, to achieve local sustained cytokine expression in the liver environment. We observed that a single injection of AAV8 expressing IL-15-IL-15RαS, but not IL-15 alone, greatly expanded the number of hepatic mononuclear cells, mainly NK cells, for at least 21 days. AAV8/IL-15-IL-15RαS treatment generated potent antitumor activity in a liver metastatic murine HCC model (BNL cells), and significantly prolonged the survival time of treated animals. The antitumor effect depended mainly on NK cells, not on CD8+ and CD4+ T cells, because AAV8/IL-15-IL-15RαS treatment greatly enhanced the cytolytic activity of hepatic NK cells and depletion of NK cells abrogated the therapeutic effect. Importantly, no apparent liver toxicity was observed during AAV8/IL-15-IL-15RαS treatment. Together, our data demonstrate that AAV8-delivered IL-15-IL-15RαS provides an effective and safe therapy against metastatic HCC.
Introduction
CD8+ T cells and natural killer (NK) cells play important roles in cancer immunosurveillance by directly killing malignant cells (Street et al., 2004; Zitvogel et al., 2006; Waldhauer and Steinle, 2008). Many studies demonstrate that interleukin (IL)-15 is a powerful cytokine for triggering the proliferation and activation of NK, natural killer T (NKT), and memory CD8+ T cells (Fehniger et al., 2002; Budagian et al., 2006; Waldmann, 2006), suggesting that IL-15 might be a potential candidate for cancer immunotherapy. Indeed, IL-15 could increase the antitumor activity of adoptively transferred CD8+ T cells or the chemotherapeutic agent cyclophosphamide through NK cell- and T cell-dependent mechanisms, but when administered alone IL-15 showed only marginal antitumor effects (Evans et al., 1997; Chapoval et al., 1998; Klebanoff et al., 2004; Teague et al., 2006). The failure of IL-15 therapy is likely due to the limiting availability of in vivo IL-15 receptor α chain (IL-15Rα), which is present on antigen-presenting cells (dendritic cells and macrophages) and is required for trans-presenting IL-15 to neighboring lymphocytes expressing the IL-15Rβ/γc receptors (Dubois et al., 2002; Koka et al., 2003; Burkett et al., 2004; Sandau et al., 2004). By mimicking trans-presentation of IL-15 by cell-associated IL-15Rα, several groups have reported that the serum half-life and biological activity of soluble IL-15 is greatly improved after preassociating with a recombinant fusion protein IL-15Rα-Fc, containing the whole extracellular domain (amino acids 1–173) of IL-15Rα and IgG Fc (Rubinstein et al., 2006; Stoklasek et al., 2006). The effect of IL-15Rα-Fc was dependent on the presence of both the IL-15Rα and IgG Fc (Dubois et al., 2008). As expected, the soluble IL-15/IL-15Rα-Fc complex demonstrated a much stronger antitumor effect in several experimental tumor models, including B16 melanoma and pancreatic tumor models (Stoklasek et al., 2006; Dubois et al., 2008; Epardaud et al., 2008).
Because the liver is heavily populated by various lymphocyte subsets, including T cells, NK cells, and NKT cells (Racanelli and Rehermann, 2006; Crispe, 2009), we hypothesize that activation of these hepatic lymphocytes by IL-15 might offer an alternative therapy against HCC. To test this hypothesis, we used the murine HCC cell line BNL to establish an orthotopic liver tumor model and the adeno-associated virus serotype 8 (AAV8) vector as a gene delivery vehicle to express a simpler form of an IL-15 superagonist, which contains covalently linked IL-15 and the N-terminal part of the extracellular domain (amino acids 1–76, called the “sushi” domain) of murine IL-15Rα (Mortier et al., 2006). The novel IL-15 agonist, designated as IL-15-IL-15RαS hereafter, was reported to be more than 100-fold more active than IL-15 alone in stimulating the proliferation of cells bearing IL-15Rβ/γc receptors (Mortier et al., 2006). AAV8 has a high liver transduction rate and low immunogenicity (Gao et al., 2002; Davidoff et al., 2005; Nakai et al., 2005), and thus may allow sustained cytokine expression in the liver to maximally activate the various hepatic lymphocyte populations. Our results demonstrate that a single treatment of AAV8 expressing IL-15-IL-15RαS led to a sustained increase in the number and cytolytic activity of hepatic mononuclear cells, mainly the NK cell subset, and, more importantly, significantly suppressed metastatic liver tumor growth, resulting in prolonged survival of treated animals.
Materials and Methods
Cell lines and mice
Murine BNL 1ME A.7R.1 (BNL; American Type Culture Collection [ATCC], Manassas, VA) is a methylcholanthrene-transformed hepatocellular carcinoma cell line derived from BALB/c mice (Patek et al., 1978). A highly liver metastatic subline of BNL (denoted hereafter BNL-h1) was generated by in vivo passage and recovery from hepatic metastases after intrasplenic inoculation of BNL cells in BALB/c mice. 3T3 (CCL-163; ATCC) is a murine fibroblast cell line derived from BALB/c mice. Cell lines were maintained in Dulbecco's modified Eagle's medium (DMEM), 10% heat-inactivated fetal calf serum, 2 mM
Construction and expression of IL-15 and IL-15-IL-15RαS
Modified IL-15 cDNA encoding the murine Igκ leader sequence fused to the IL-15 mature sequence was generated by overlapping polymerase chain reaction (PCR). Replacement of the native IL-15 leader peptide, which has a negative regulatory function (Onu et al., 1997; Bamford et al., 1998), with the Igκ leader peptide could potentially increase the secretion and biological functions of IL-15 (Kutzler et al., 2005). The DNA fragment encoding IL-15-IL-15RαS was assembled by two-step overlapping PCR with a 26-amino acid linker (SGGGSGGGGSGGGGSGGGGSGGGSLQ) (Mortier et al., 2006) covalently bridging the IL-15 and IL-15RαS domains (see Fig. 1A). These PCR fragments were subcloned into the pAAVEMBL plasmid (Wang et al., 2003) to generate pAAVEMBL/IL-15 and pAAVEMBL/IL-15-IL-15RαS. The control pAAVEMBL/GFP plasmid encoding the green fluorescent protein has been described (Wang et al., 2003). Recombinant AAV8 vectors were produced by a standard triple-plasmid transfection method, purified by two rounds of CsCl centrifugation, and quantitated by real-time PCR as described previously (Xiao et al., 1998).
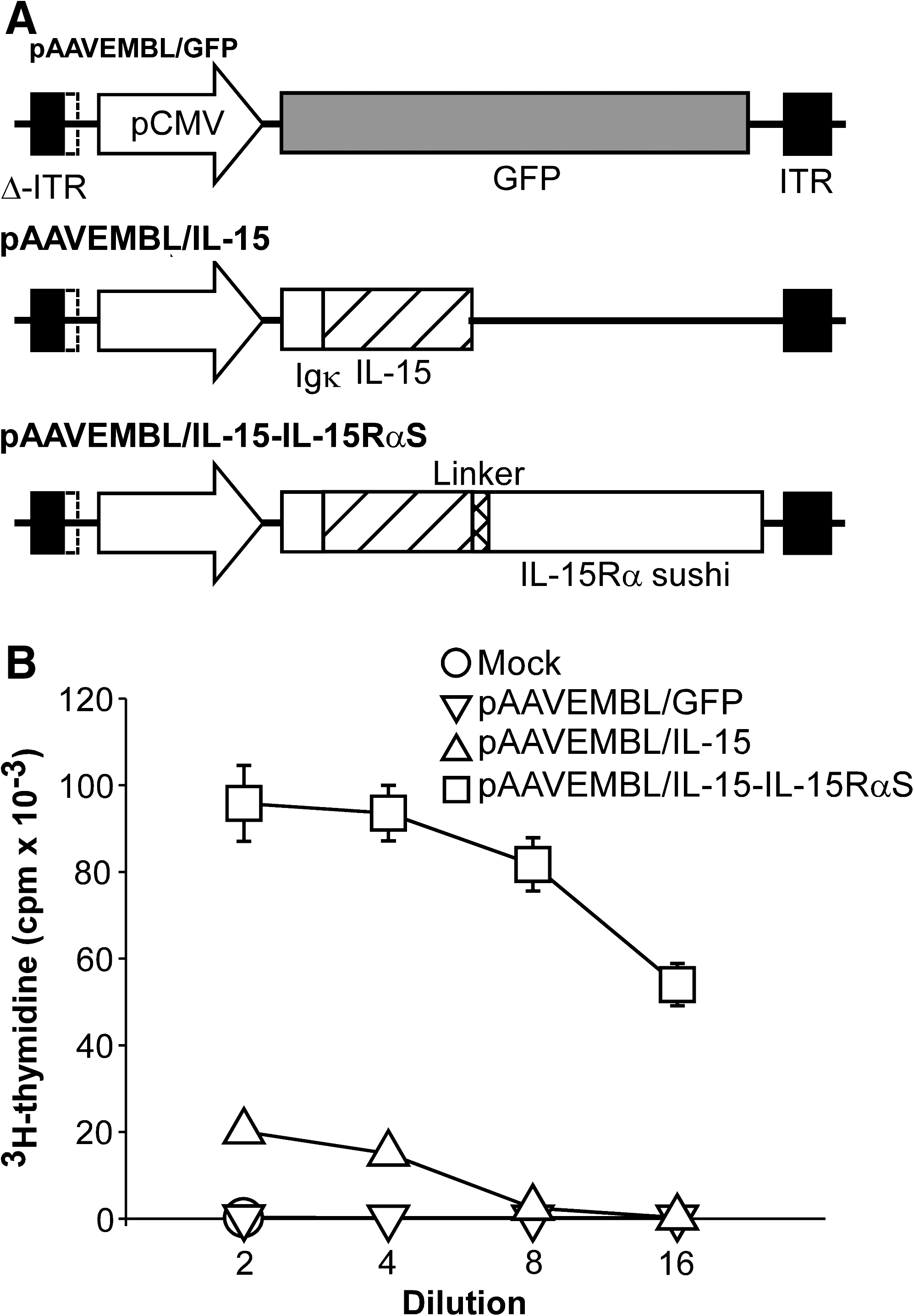
Construction and characterization of pAAVEMBL plasmids encoding IL-15 or IL-15-IL-15RαS. (
HT-2 cell proliferation assay
To examine IL-15 and IL-15-IL-15RαS expression and function, 3T3 cells were transiently transfected with 5 μg of pAAVEMBL plasmid, using Lipofectamine 2000 (Invitrogen, Carlsbad, CA), and 48 hr later cell-free supernatants were collected to stimulate proliferation of HT-2 cells, a murine IL-2/IL-15-dependent T cell line, as previously described (Liu et al., 1998).
Flow cytometry
Splenocytes and hepatic mononuclear cells were isolated on a Percoll density gradient as described (Tamaki et al., 2005). Briefly, the mice were killed and bled by heart puncture. The liver was removed and gently passed through a stainless steel mesh, and the cells were washed with phosphate-buffered saline (PBS). Hepatic mononuclear cells were separated from parenchymal cells by centrifugation at 50 × g for 5 min. Hepatic cells prepared from one mouse were suspended in 20 ml of 33% Percoll gradient solution (GE Healthcare, Piscataway, NJ) and centrifuged at 754 × g for 18 min. The pellet was treated with red blood cell lysis buffer, washed with PBS, and used as hepatic mononuclear cells. The surface phenotype of the mononuclear cells was characterized by flow cytometric analysis. Cells were preincubated for 20 min on ice with anti-CD16/32 monoclonal antibody (mAb) (2.4G2; ATCC) to block nonspecific binding of antibodies to Fc receptors, and then were incubated for 30 min on ice in the dark with the following mAbs diluted in PBS containing 0.1% bovine serum albumin: fluorescein isothiocyanate (FITC)-conjugated anti-mouse CD3ɛ (145-2C11), phycoerythrin (PE)-conjugated anti-mouse CD8 (53-6.7), FITC-conjugated anti-mouse CD4 (RM4-4), allophycocyanin (APC)-conjugated anti-mouse pan-NK (DX5), or an isotype-matched control mAb. All mAbs were purchased from either BD Biosciences (San Jose, CA) or BioLegend (San Diego, CA). After washing with PBS, the stained cells were analyzed with a FACSCanto (BD Biosciences) and the data were processed with FlowJo V.7.2.5 software (Treestar, Ashland, OR).
Tumor treatment studies
To generate hepatic metastases, 6- to 8-week-old female BALB/c mice were injected intrasplenically with 1 × 106 BNL-h1 cells, a dose consistently yielding hepatic metastases in 100% of animals. In the preventive model, mice were injected intravenously with AAV8/IL-15-IL-15RαS at 1 × 1010 or 1 × 1011 vector genome (VG) per mouse or with AAV8/GFP at 1 × 1011 VG per mouse, followed, 10 days later, by intrasplenic injection of BNL-h1 cells. In the therapeutic model, mice were first injected intrasplenically with 1 × 106 BNL-h1 cells, which generated an average of 11 ± 3 disseminated metastatic loci, as viewed under a dissecting microscope. These mice were then injected intravenously with 3 × 1011 VG per mouse of AAV8/IL-15-IL-15RαS or AAV8/GFP or saline. To identify the contribution of lymphocyte subpopulations to the antitumor mechanism, the tumor-bearing mice were depleted of CD4+ T cells, CD8+ T cells, or NK cells by intraperitoneal injection of anti-CD4 mAb (GK1.5, rat IgG2b), anti-CD8 mAb (53-6.72, rat IgG2a), or rabbit anti-asialo-GM1 antiserum (Wako Pure Chemical, Osaka, Japan), respectively, at 1, 4, 6, 8, and 15 days after tumor inoculation, and received a single AAV8/IL-15-IL-15RαS treatment 3 days after tumor inoculation. Our previous data showed that more than 95% of the respective lymphocyte populations were depleted by this treatment dose and schedule (Lo et al., 2003). Mice treated with the same dose and schedule of a monoclonal normal rat IgG (HAA, rat IgG2a), derived from a hybridoma clone isolated from a naive rat, and normal rabbit serum (BioWest, Nuaille, France) were included as controls. All mice were killed 3 weeks after tumor cell inoculation, a few days before the tumor-bearing mice began to die, and the number of hepatic metastases were counted under a dissecting microscope. In some experiments, mice were not killed but were monitored for long-term survivors.
Cytotoxicity assay
A chromium release assay was used to measure the ability of liver mononuclear cells to lyse BNL-h1 target cells and NK-sensitive YAC-1 cells (Lo et al., 2003). Briefly, effector liver mononuclear cells and 51Cr-labeled target cells at various effector-to-target (E/T) ratios were incubated for 6 hr at 37°C and 5% CO2, and radioactivity in the supernatant was measured. The percentage of cytotoxicity was calculated from the following formula: [(mean experimental cpm – mean spontaneous cpm)/(mean maximal cpm – mean spontaneous cpm)] × 100. Spontaneous release of 51Cr (incubation of target cells with medium alone) was ∼7% of maximal release (incubation of target cells with 10% Triton X-100) for YAC-1 and ∼15% for BNL-h1.
Histology and immunohistochemistry
Liver sections were fixed in 4% paraformaldehyde in PBS, embedded in paraffin, sectioned (5 μm), and stained with hematoxylin and eosin. The sections were mounted and observed by light microscopy. Immunohistochemical analysis of liver samples was performed as described previously (Lo et al., 2003). Briefly, 10-μm cryostat sections were prepared and fixed in cold acetone and endogenous peroxidase activity was blocked by incubation with 3% H2O2. The slides were then incubated overnight at 4°C with rabbit anti-asialo-GM1 antiserum (Wako Pure Chemical) or normal rabbit serum followed by incubation for 1 hr at room temperatures with horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG antibody. Streptavidin peroxidase (R&D Systems, Minneapolis, MN) was added and the color was developed with a Zymed AEC substrate kit (Zymed Laboratories, San Francisco, CA). The slides were counterstained with hematoxylin and analyzed by light microscopy at ×200 magnification.
Measurement of serum transaminase activity and interferon-γ
Serum alanine aminotransferase (ALT) activity was measured with a Vitros 950 chemical analyzer (Johnson & Johnson, Rochester, NY). Values are expressed as units per liter (U/liter). Interferon (IFN)-γ was measured by ELISA (mouse IFN-γ DuoSet ELISA development system; R&D Systems) according to the manufacturer's instructions.
Statistics
All data were analyzed for significance by the Student t test. p < 0.05 was considered significant.
Results
Construction and characterization of AAV8 expressing IL-15 and IL-15-IL-15RαS
To evaluate the antitumor potential of IL-15 in an orthotopic HCC model, we employed the AAV8 vector to deliver murine IL-15 or the IL-15 superagonist (IL-15-IL-15RαS) to target cytokine expression in the liver environment. In these constructs, the native IL-15 leader peptide was replaced with the murine Igκ leader peptide to increase IL-15 secretion and functions (Kutzler et al., 2005). Expression of IL-15, IL-15-IL-15RαS, and the control GFP protein was under the transcriptional control of the cytomegalovirus (CMV) promoter (Fig. 1A).
We next examined the expression of IL-15 and IL-15-IL-15RαS by transiently transfecting 3T3 cells and assaying the culture medium. ELISAs detected high titers of IL-15 and IL-15Rα produced, respectively, by pAAVEMBL/IL-15 and pAAVEMBL/IL-15-IL-15RαS (see Supplementary Fig. 1 at
The biological function of IL-15 and IL-15-IL-15RαS was evaluated by their ability to stimulate proliferation of murine IL-2/IL-15-responsive HT-2 cells. As shown in Fig. 1B, both IL-15 and IL-15-IL-15RαS clearly stimulated proliferation of HT-2 cells in a dose-dependent manner (Fig. 1B). IL-15-IL-15RαS was much more potent than IL-15 at the same fold dilution. Transfection with the control pAAVEMBL/GFP plasmid did not produce detectable IL-15 or IL-15Rα (Supplementary Fig. 1) and was completely negative in the HT-2 proliferation assay (Fig. 1B).
To evaluate the in vivo biological functions of IL-15 and IL-15-IL-15RαS, the pAAVEMBL/IL-15, pAAVEMBL/IL-15-IL-15RαS, and pAAVEMBL/GFP plasmids were used to generate AAV8 vectors, and were designated, respectively, AAV8/IL-15, AAV8/IL-15-IL-15RαS, and AAV8/GFP. Groups of BALB/c mice were injected intravenously with 3 × 1011 VG of the various AAV8 vectors or saline, and at various time points (days 3, 7, 14, and 21) three mice from each group were analyzed for total cell number and cellular composition of the hepatic mononuclear cells and splenocytes. Treatment with AAV8/IL-15-IL-15RαS significantly increased the number of total hepatic mononuclear cells and splenocytes 3 days after AAV administration, and the elevated cell numbers were maintained across the entire observation period (21 days). The representative day-21 result is shown in Fig. 2A, in which IL-15-IL-15RαS increased the number of hepatic mononuclear cells by 7.2-fold (p = 0.002) and splenocytes by 3.0-fold (p = 0.0004) compared with the saline group. Flow cytometric analysis revealed that NK cells (CD3–DX5+) were the main subpopulation activated by IL-15-IL-15RαS, with a 63.1-fold increase in the liver (p = 0.001 vs. the saline group) and a 46.9-fold increase in the spleen (p = 0.0002 vs. the saline group) (Fig. 2A). In fact, treatment with AAV8/IL-15-IL-15RαS quickly elevated the relative abundance of NK cells from 11 to 76% in the liver and from 4 to 40% in the spleen within 7 days after injection, and maintained such a high level for at least 21 days (Fig. 2B). A comparable expansion of NK cells was also observed in the peripheral blood (data not shown). AAV8/IL-15-IL-15RαS also significantly, but to a lesser extent, increased NKT cells (CD3+DX5+) by 1.9-fold in the liver (p = 0.046 vs. the saline group) and 2.6-fold in the spleen (p = 0.0003 vs. the saline group), and CD8+ T cells by 1.8-fold (p = 0.004 vs. the saline group) in the spleen, but had no effect on the total number of CD8+ T cells in the liver and CD4+ T cells in both the liver and spleen (Fig. 2A). Compared with the strong activity of IL-15-IL-15RαS, IL-15 alone showed a modest in vivo effect, only slightly increasing the number of NK cells in the liver (2.6-fold; p = 0.013) and in the spleen (1.6-fold; p = 0.001). The control AAV8/GFP treatment had no effect on any of the lymphocyte subpopulation in the liver and spleen.

In vivo effect of AAV8-delivered IL-15 or IL-15-IL-15RαS. BALB/c mice were injected intravenously with saline, or 3 × 1011 VG per mouse of AAV8/GFP, AAV8/IL-15, or AAV8/IL-15-IL-15RαS. On the indicated day, three mice of each group were killed for analysis of hepatic mononuclear cells and splenocytes by flow cytometry. (
Treatment of hepatic tumors
We chose AAV8/IL-15-IL-15RαS for further antitumor study because of its much stronger biological activity as compared with AAV8/IL-15. In the first set of experiments, we determined whether pretreatment with AAV8/IL-15-IL-15RαS could prevent hepatic metastases of BNL-h1 cells, a murine HCC cell line. BNL-h1 cells do not express the IL-2/IL-15Rβ chain (see Supplementary Fig. 2 at
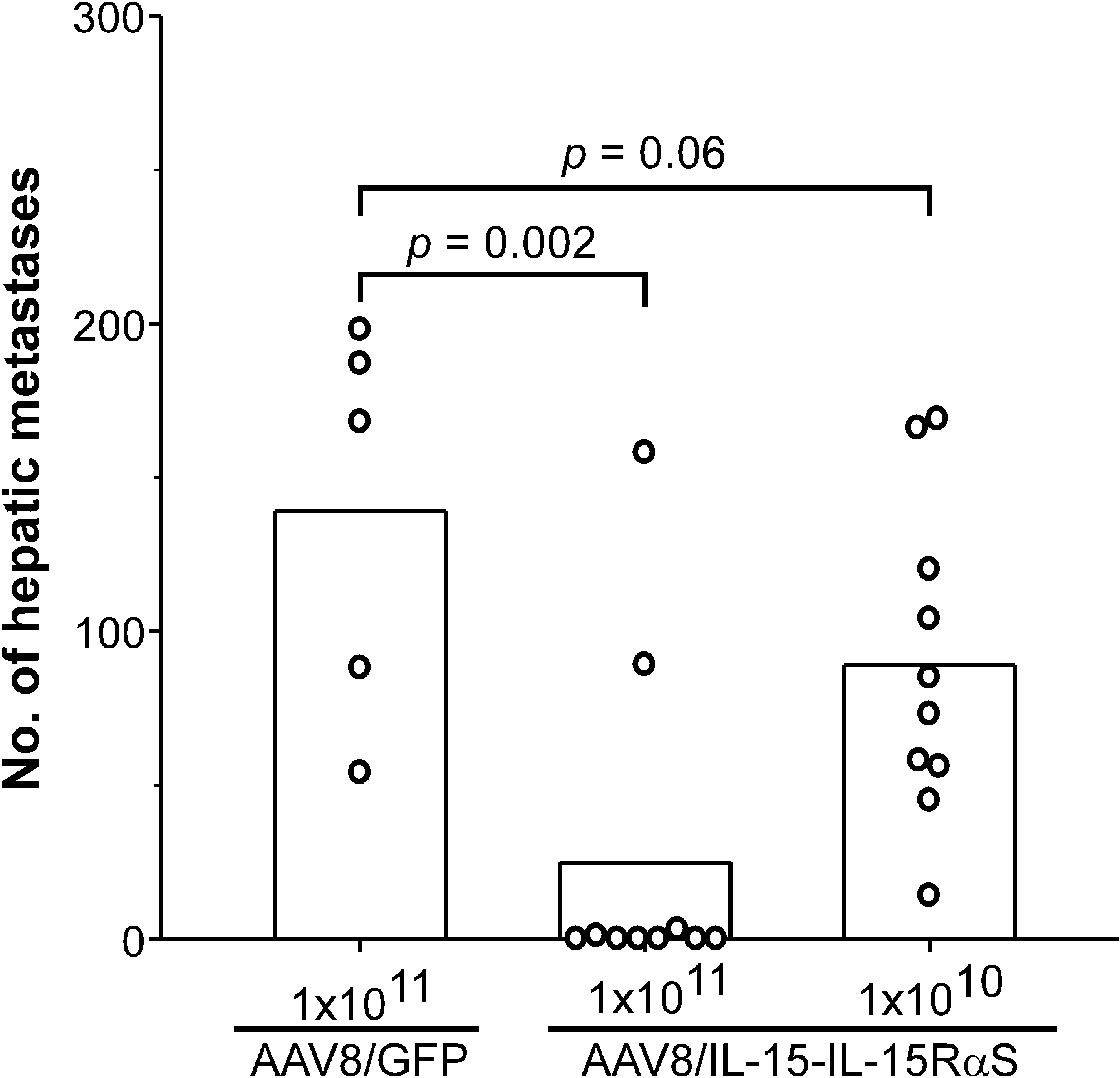
Prevention of hepatic metastases of BNL-h1 by AAV8/IL-15-IL-15RαS treatment. Groups of BALB/c mice (n = 5–10) were injected intravenously with AAV8/IL-15-IL-15RαS at 1 × 1010 or 1 × 1011 VG per mouse or with AAV8/GFP at 1 × 1011 VG per mouse; then, 10 days later, they received an intrasplenic injection of 1 × 106 BNL-h1 cells to induce hepatic metastases. The mice were killed 21 days after tumor challenge and the number of hepatic metastases was counted. Data are presented as the number for the individual mice and the mean of each group. The experiment was repeated twice with similar results.
To examine the therapeutic effect of AAV8/IL-15-IL-15RαS against preestablished hepatic metastases, mice were intrasplenically injected with 1 × 106 BNL-h1 tumor cells and 3 days later treated with, per mouse, 3 × 1011 VG of AAV8/IL-15-IL-15RαS (n = 8) or AAV8/GFP (n = 8) or with saline alone (n = 5). The number of hepatic foci in each individual mouse in each group is shown in Fig. 4A. AAV8/IL-15-IL-15RαS treatment significantly suppressed metastatic tumor growth (mean, 39; range, 0–170; p = 0.00006) as compared with mice injected with saline (mean, 195; range, 145–245). In contrast, treatment with the control AAV8/GFP had not therapeutic benefit (mean, 178; range, 99–260; p = 0.24 vs. the saline group).
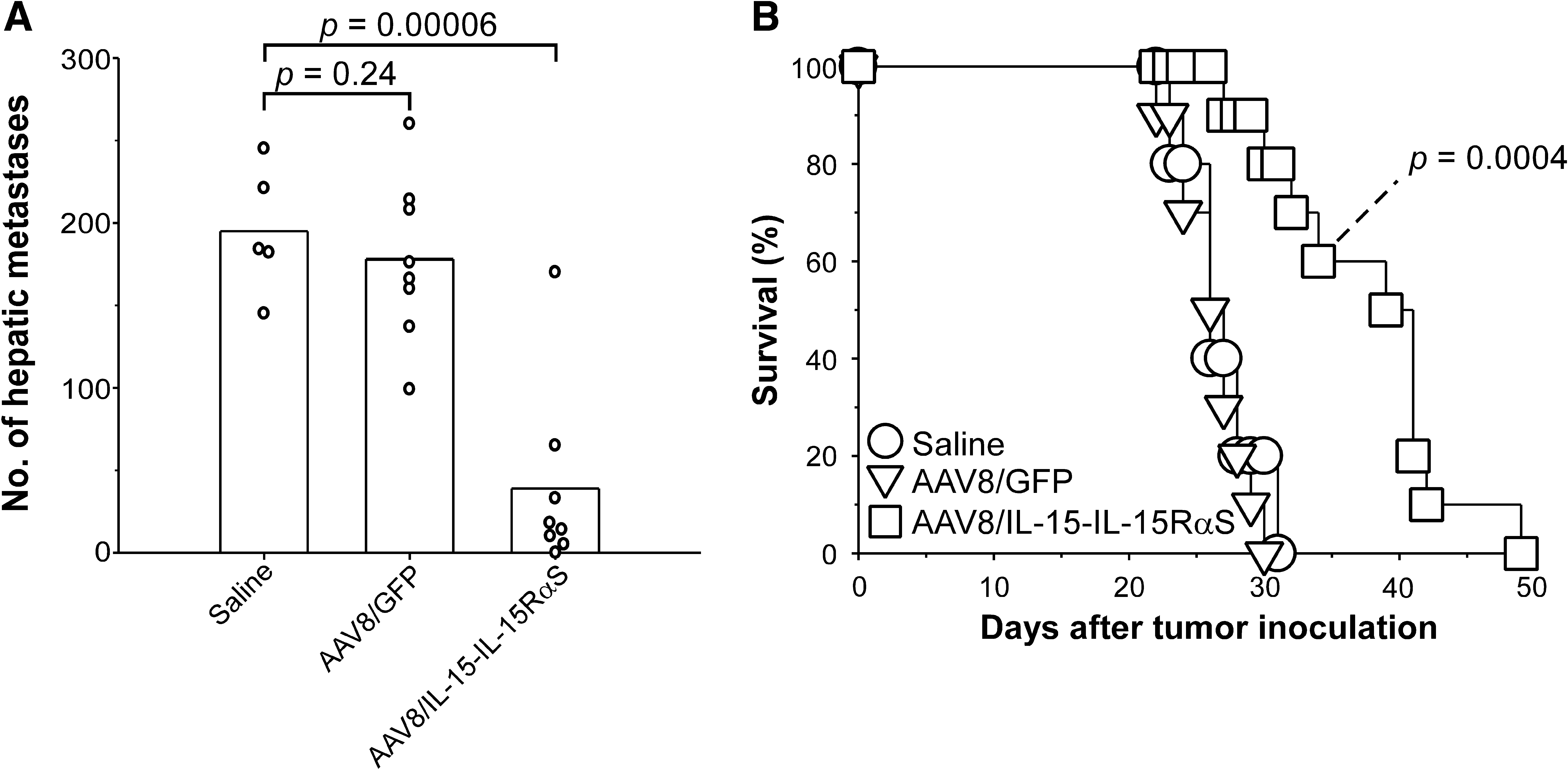
Treatment of established hepatic metastases of BNL-h1 by AAV8/IL-15-IL-15RαS. (
We then investigated whether treatment with IL-15-IL-15RαS could result in a survival benefit for mice with established hepatic metastases. Groups of BALB/c mice (n = 10) with 3-day hepatic metastases were injected with, per mouse, 3 × 1011 VG of AAV8/IL-15-IL-15RαS or AAV8/GFP or with saline. As shown in Fig. 4B, treatment with AAV8/IL-15-IL-15RαS significantly prolonged the mean survival time of the treated mice (38 ± 7 days; p = 0.0004) as compared with the mice injected with saline (27 ± 3 days). Mice treated with AAV8/GFP had no beneficial effect on the mean survival time (26 ± 2 days; p = 0.38 vs. the saline group).
Mechanisms of the antitumor effect of AAV8/IL-15-IL-15RαS
To investigate the antitumor mechanisms of IL-15-IL-15RαS, tumor-bearing BALB/c mice (n = 5) treated with AAV8/IL-15-IL-15RαS, AAV8/GFP, or saline as described previously were killed 21 days after tumor cell inoculation for histological and immunohistochemical analyses. Histological analysis revealed aggregation of BNL-h1 cells with fibrous stroma in the liver of all mice, with a much smaller average tumor size in mice treated with AAV8/IL-15-IL-15RαS (Fig. 5C) compared with AAV8/GFP (Fig. 5B) or saline (Fig. 5A). Intense infiltration of mononuclear cells was detected in the tumor lesions of AAV8/IL-15-IL-15RαS-treated mice, but much less in those treated with AAV8/GFP or saline. There were also many mononuclear cells scattered throughout the liver in the AAV8/IL-15-IL-15RαS-treated mice than in mice from the other two groups. These liver mononuclear cells in the AAV8/IL-15-IL-15RαS-treated mice were mostly NK cells, as revealed by immunohistochemical staining with asialo-GM1-specific antibody (Fig. 5F). An ex vivo cytotoxic assay revealed that liver mononuclear cells from AAV8/IL-15-IL-15RαS-treated mice efficiently lysed prototype NK cell target YAC-1 cells (Fig. 6A) and BNL-h1 tumor cells (Fig. 6B), whereas liver mononuclear cells from the AAV8/GFP group failed to elevate the NK cytolytic activity as compared with those from the saline group. The increase in hepatic NK cells 21 days after AAV administration in AAV8/IL-15-IL-15RαS-treated mice is consistent with a sustained elevated level of IL-15-IL-15RαS in the serum of these mice, ranging between 565 ± 191 pg/ml on day 7 and 339 ± 58 pg/ml on day 21 (see Supplementary Fig. 3 at

Histological and immunohistochemical evaluation of liver tumors treated with AAV8/IL-15-IL-15RαS. Groups of BALB/c mice (n = 5) bearing 3-day BNL-h1 hepatic tumors were treated with saline or AAV8 vectors expressing IL-15-IL-15RαS or GFP as described in the caption to Fig. 4. Mice were killed 21 days after tumor inoculation and the liver tissues were removed for histological and immunohistochemical analysis. Paraffin sections were stained with hematoxylin and eosin (
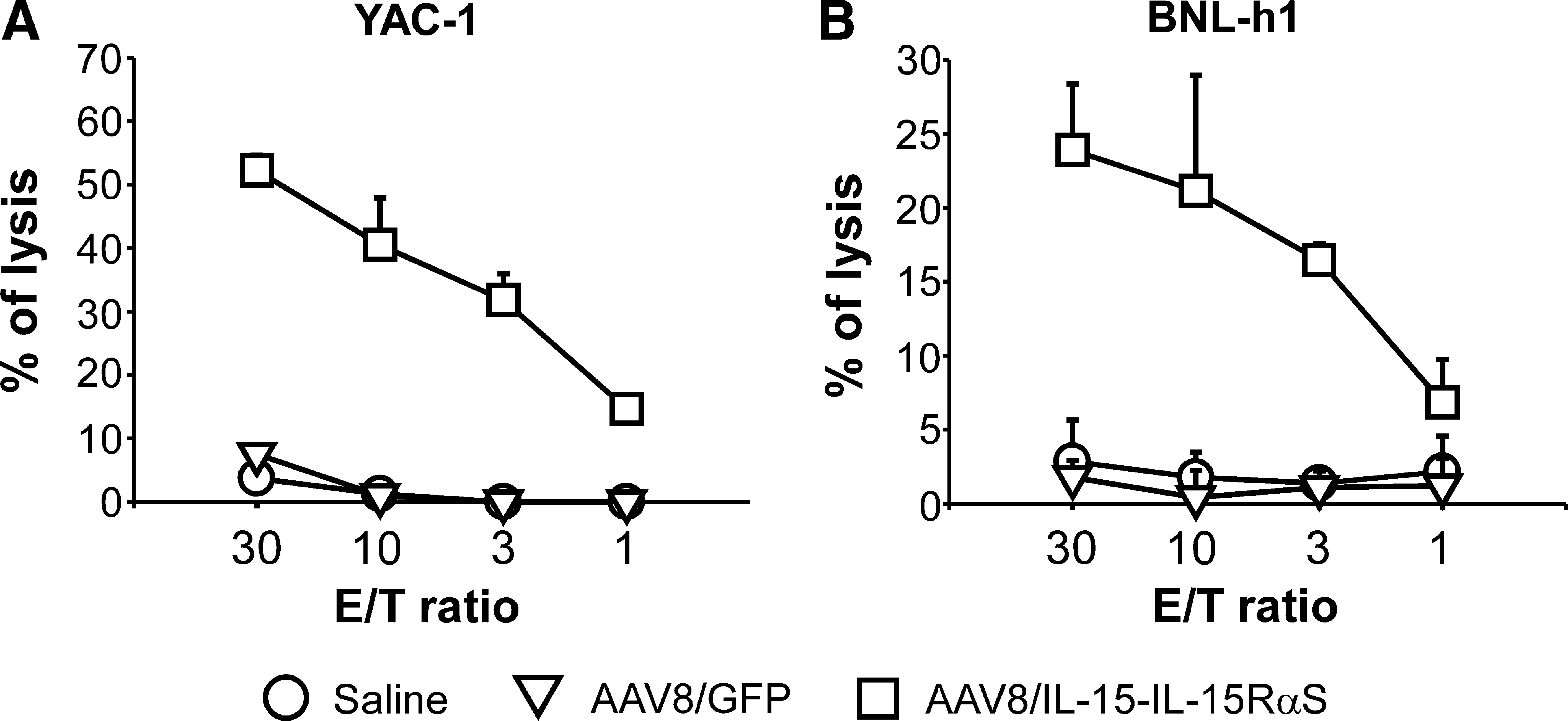
Cytotoxic activity of hepatic mononuclear cells against YAC-1 and BNL-h1 cells. Fresh hepatic mononuclear cells were isolated from the mice described in Fig. 5, and tested on 51Cr-labeled YAC-1 cells (
To further assess the role of the various lymphocyte populations in IL-15-IL-15RαS-mediated antitumor activity, tumor-bearing BALB/c mice were depleted of CD4+ T cells, CD8+ T cells, or NK cells by injection of anti-CD4 mAb, anti-CD8 mAb, or anti-asialo-GM1 antiserum, respectively, before and after AAV8/IL-15-IL-15RαS treatment. Mice treated with an irrelevant rat monoclonal IgG or normal rabbit serum at the same dose and schedule were included as controls. As shown in Fig. 7, depletion of NK cells impaired the antitumor effect of AAV8/IL-15-IL-15RαS, resulting in a significant increase in hepatic metastases (mean, 160; range, 89–253; p = 0.002) as compared with mice receiving normal rabbit serum (mean, 30; range, 5–66). Depletion of CD8+ T cells had no effect on hepatic tumors (mean, 45; range, 27–84; p = 0.37) as compared with the rat IgG control (mean, 38; range, 3–92), whereas depletion of CD4+ T cells tended to decrease the number of hepatic metastases, but the effect was not statistically significant (mean, 6; range, 3–10; p = 0.06).
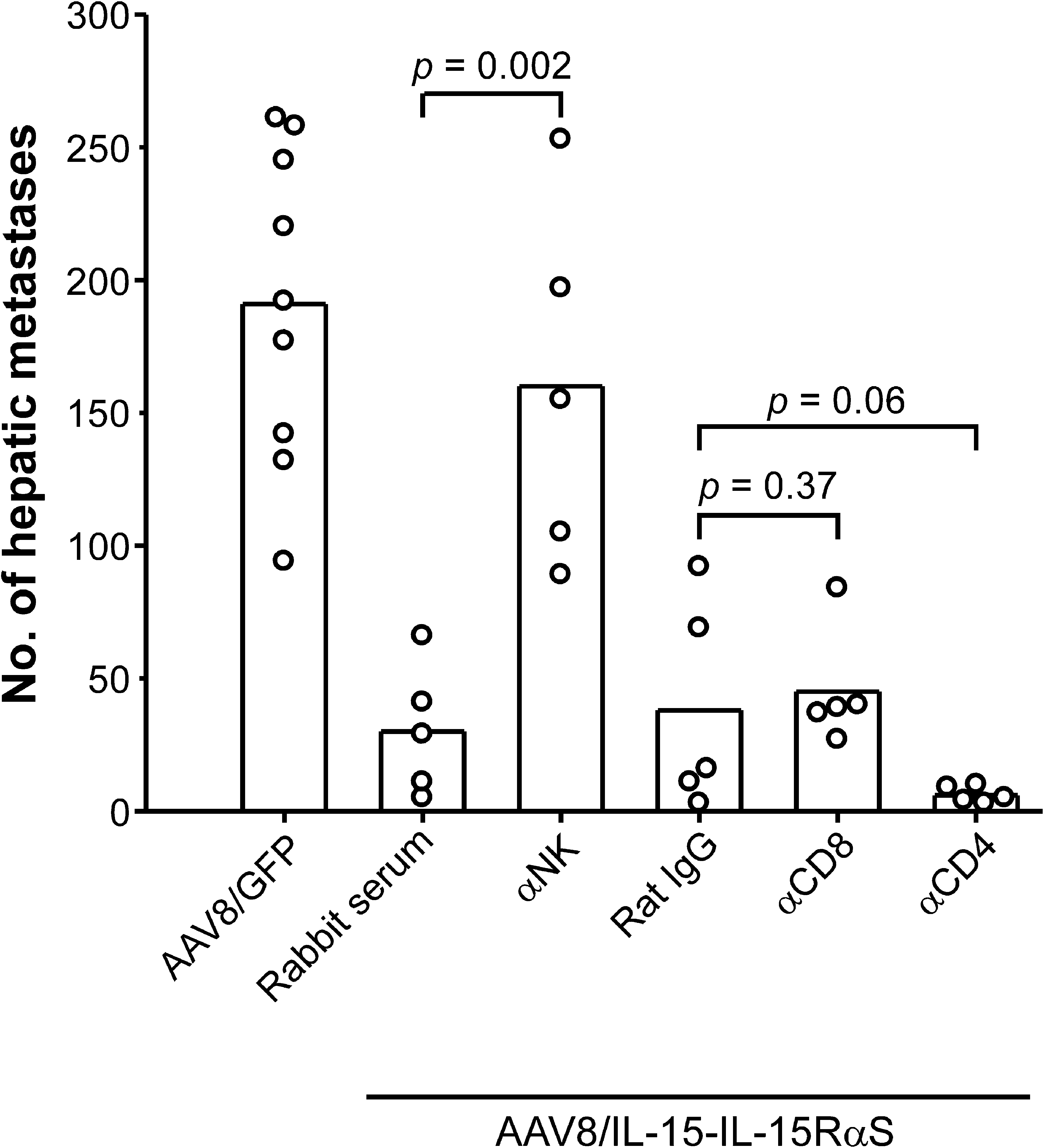
Effect of selective depletion of various lymphocyte populations on AAV8/IL-15-IL-15RαS-mediated antitumor effect. BALB/c mice were injected intrasplenically with BNL-h1 cells on day 0 and then, 3 days later, were treated, per mouse, with 3 × 1011 VG of AAV8/IL-15-IL-15RαS or AAV8/GFP. Mice (n = 5 per group) that were treated with AAV8/IL-15-IL-15RαS received injections of anti-asialo-GM1 antiserum (depletion of NK cells), anti-CD8 mAb, or anti-CD4 mAb on days 1, 4, 6, 8, and 15. Mice treated with the same dose and schedule of a monoclonal normal rat IgG or normal rabbit antiserum were included as controls. Data, the results of one experiment, are representative two performed independently.
No liver toxicity induced by AAV8/IL-15-IL-15RαS treatment
Last, we evaluated whether liver toxicity was induced by AAV8/IL-15-IL-15RαS treatment. BALB/c mice (n = 5) were injected intravenously with, per mouse, 3 × 1011 VG of AAV8/IL-15-IL-15RαS or AAV8/GFP or with saline, and the serum ALT level was determined at various times after AAV injection. As shown in Fig. 8A, mice treated with AAV8/IL-15-IL-15RαS, like those injected with AAV8/GFP or saline, did not induce apparent liver toxicity as serum ALT levels in these mice remained in the normal range (<40 U/liter) across the entire observation period. In contrast, mice injected intravenously with an adenoviral vector expressing IL-12 (Ad/IL-12), one of the most potent antitumor cytokines, led to a marked elevation of serum ALT at all time points. Because IFN-γ is intimately associated with IL-12-mediated liver injury (Car et al., 1995), we also examined the serum IFN-γ level in mice treated with the AAV vectors. As shown in Fig. 8B, AAV8/IL-15-IL-15RαS treatment did not induce detectable serum IFN-γ, which is in sharp contrast to the quick elevation of serum IFN-γ after Ad/IL-12 administration.
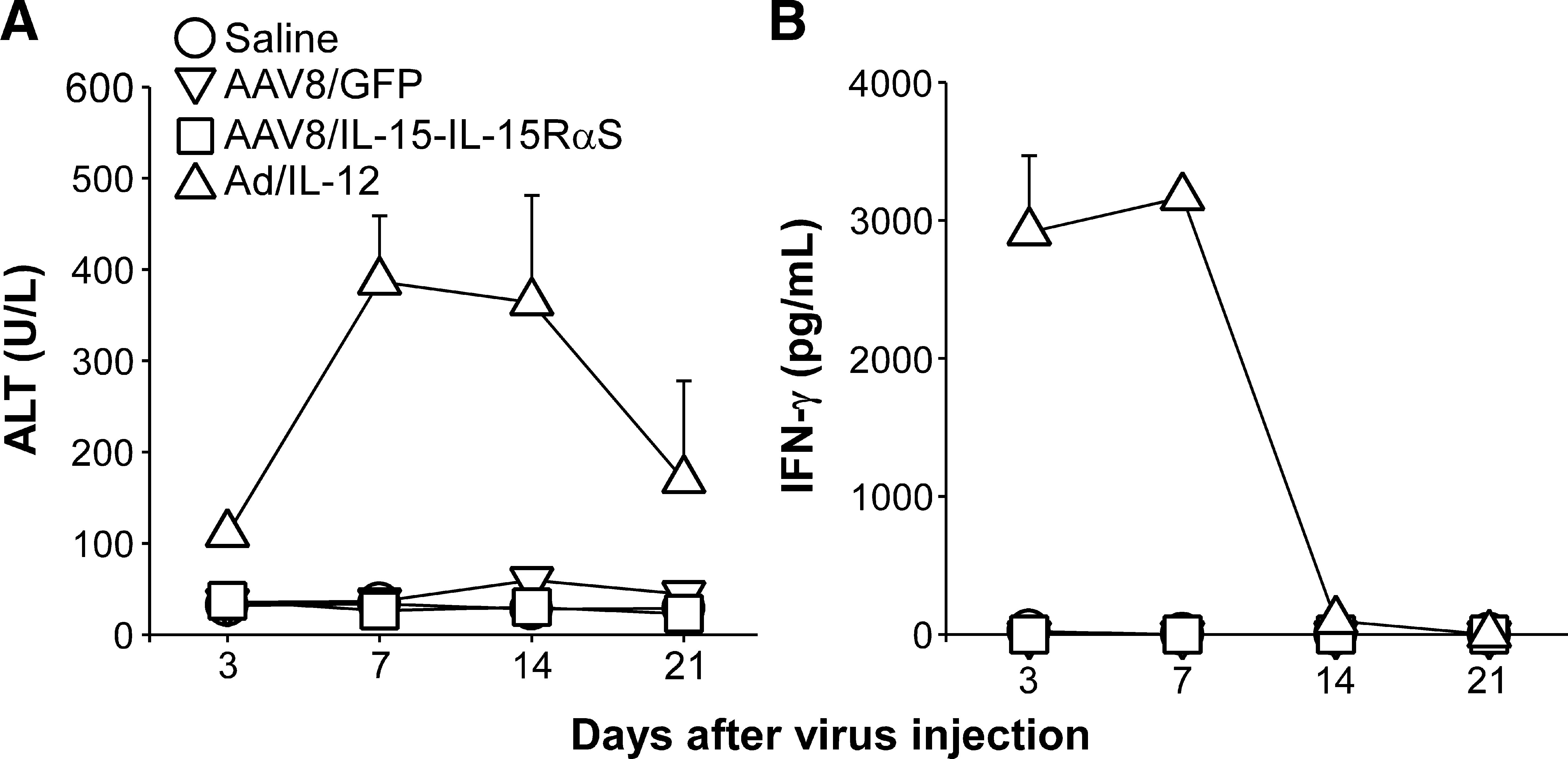
Serum ALT and IFN-γ levels in AAV8/IL-15-IL-15RαS-treated mice. Groups of BALB/c mice (n = 5) were injected intravenously with, per mouse, 3 × 1011 VG of AAV8/GFP or AAV8/IL-15-IL-15RαS. Mice injected with saline or with 1 × 109 plaque-forming units of adenovirus carrying the mouse IL-12 gene (Ad/IL-12) were included as negative and positive controls, respectively. At the indicated time points, serum samples were collected to measure ALT (
Discussion
HCC is one of the most common cancers worldwide, but effective therapies are still needed (Avila et al., 2006; El-Serag et al., 2008; Llovet and Bruix, 2008). To take advantage of the abundant lymphocytes present in the liver environment, we constructed an AAV8 vector encoding IL-15-IL-15RαS, an IL-15 superagonist, to stimulate differentiation and augment functionality of hepatic lymphocytes to treat HCC. Our data demonstrate that AAV8/IL-15-IL-15RαS treatment generated potent antitumor activity against metastatic HCC and significantly prolonged the survival time of treated animals. More importantly, no apparent liver toxicity was observed during the treatment.
The poor in vivo bioactivity of IL-15 can be greatly improved by preassociating IL-15 with a soluble IL-15Rα-Fc protein (Rubinstein et al., 2006; Stoklasek et al., 2006; Dubois et al., 2008) or by covalently linking IL-15 to the sushi domain of IL-15Rα via a flexible peptide linker (Mortier et al., 2006). The superiority of these exogenous IL-15-IL-15Rα complexes over IL-15 alone is likely due to their increased serum half-lives, increasing from ∼30 min for IL-15 to ∼20 hr for IL-15-IL-15Rα-Fc (Stoklasek et al., 2006) and ∼3 hr for IL-15-IL-15RαS (Bessard et al., 2009), and enhanced biological functions, increasing ∼50-fold for IL-15-IL-15Rα-Fc (Rubinstein et al., 2006; Stoklasek et al., 2006; Dubois et al., 2008; Epardaud et al., 2008) and more than 100-fold for IL-15-IL-15RαS (Mortier et al., 2006; Bessard et al., 2009). We decided to use IL-15-IL-15RαS in our study because of its higher activity compared with IL-15-IL-15Rα-Fc and because of its broader target populations, which require expression of IL-15Rβ/γc only, whereas both IL-15Rβ/γc and IgG Fc receptors are required for the optimal function of IL-15-IL-15Rα-Fc (Dubois et al., 2008). We used a hepatotropic AAV8 vector to preferentially express IL-15-IL-15RαS in the liver environment to improve its therapeutic effect against liver cancer, a strategy supported by the finding that a tumor-targeting antibody-conjugated IL-15 displayed better antitumor activity than a nonspecific antibody-conjugated IL-15 (Kaspar et al., 2007).
Our result demonstrated that AAV-mediated expression of IL-15-IL-15RαS, but not IL-15 alone, is powerful in expanding the number of hepatic and splenic lymphocytes in vivo, mainly NK cells but to a lesser extent NKT cells and CD8+ T cells (Fig. 2A). Compared with soluble IL-15-IL-15Rα complexes, which required multiple injections to maintain a long-term biological effect (Stoklasek et al., 2006; Dubois et al., 2008; Epardaud et al., 2008), a single treatment with AAV8/IL-15-IL-15RαS resulted in a high and sustained level of NK cells for at least 21 days, the longest observation period of our experiment (Fig. 2B). These results suggest that AAV8/IL-15-IL-15RαS could be used in controlling infectious and malignant diseases that are sensitive to NK cell-mediated cytotoxicity (Lee et al., 2007; Zamai et al., 2007; Terme et al., 2008).
We demonstrated that administration of a single dose of AAV8/IL-15-IL-15RαS effectively prevented hepatic metastases (Fig. 3) and enhanced the treatment of established tumors (Fig. 4A). Most importantly, AAV8/IL-15-IL-15RαS treatment significantly prolonged the survival time of treated mice (Fig. 4B). A direct effect of IL-15-IL-15RαS on BNL tumor can be ruled out, because BNL-h1 cells do not express a detectable amount of the IL-2/IL-15Rβ chain (Supplementary Fig. 2). Histological analysis demonstrated that mice treated with AAV8/IL-15-IL-15RαS, as compared with those injected with saline or AAV8/GFP, had much smaller tumors and many more mononuclear cells infiltrating the tumor lesions as well as the liver (Fig. 5A–C). Immunohistological staining revealed that these hepatic mononuclear cells were mostly asialo-GM1 cells (mainly NK cells) (Fig. 5F). Phenotypic analysis demonstrated that CD69 was upregulated on the hepatic mononuclear cells isolated from IL-15-IL-15RαS-treated mice as compared with those injected with saline or AAV8/GFP (data not shown), suggesting that these tumor-infiltrating lymphocytes were in an activated state. Indeed, we demonstrated that the isolated hepatic mononuclear cells efficiently lysed NK-sensitive YAC-1 cells and BNL-h1 tumor cells (Fig. 6A). The importance of NK cells in constraining liver tumor growth was further supported by the observation that depletion of NK cells by anti-asialo-GM1 antiserum abrogated the tumor therapeutic effect of AAV8/IL-15-IL-15RαS, whereas depletion of CD8+ or CD4+ T cells had no significant effect on treatment outcome (Fig. 7). Taken together, our data suggest that AAV8/IL-15-IL-15RαS is effective in controlling liver tumor growth through an NK cell-dependent mechanism. Because IL-15 is also known to increase the antitumor activity of cyclophosphamide (Evans et al., 1997; Chapoval et al., 1998), it is likely that a combination therapy of AAV8/IL-15-IL-15RαS and chemotherapeutic agents might provide a more effective therapy for HCC treatment. While we were preparing this paper, we became aware of a report showing that the soluble IL-15-IL-15RαS complex exhibited higher antitumor activity than IL-15 in two other cancer models, metastatic melanoma and colorectal cancer (Bessard et al., 2009). Our work is in agreement with and also expands these studies by providing a more convenient method for targeted cytokine delivery and a more detailed analysis of the cellular mechanisms underlying the antitumor effect of IL-15-IL-15RαS.
Despite induction of massive hepatic mononuclear cells and a great increase in their cytotoxicity, mice treated with AAV8/IL-15-IL-15RαS showed no evidence of liver injury during the entire experimental course. The serum ALT levels of all AAV8/IL-15-IL-15RαS-treated mice remained within the normal range for up to 21 days after treatment (Fig. 8A). Other studies also demonstrated that expression of high levels of IL-15 or both IL-15 and IL-15Rα from plasmid vectors did not induce any apparent toxic effects in the treated animals (Giri et al., 1994; Bergamaschi et al., 2008). However, we did observe enlarged spleens, likely due to an increased number of splenocytes after AAV8/IL-15-IL-15RαS treatment (Fig. 2A), a phenomenon also reported in mice treated with IL-15-IL-15Rα-Fc complex (Rubinstein et al., 2006). It should be noted that IL-15 seems to provide protection in several liver injury models (Li et al., 2006; Suzuki et al., 2006) and against chemotherapy-induced gastrointestinal toxicity (Cao et al., 1998a,b). This protective effect of IL-15 is in sharp contrast to the significant and consistently elevated serum ALT observed in mice treated with adenovirus-delivered IL-12 (Fig. 8A), another potent cytokine for liver cancer treatment (Del Vecchio et al., 2007).
IL-2 is a Food and Drug Administration-approved drug for cancer treatment, but its efficacy is limited by systemic toxicity and induction of peripheral tolerance due to selective promotion of T cell activation-induced cell death (Marks-Konczalik et al., 2000) and expansion of T-regulatory cells (Fontenot et al., 2005; Antony et al., 2006). In contrast, IL-15, which shares with IL-2 two receptor subunits (CD122 and CD132) and induces similar intracellular signaling events (Waldmann, 2006), does not have these adverse effects of IL-2, but instead could inhibit IL-2-induced T cell death. These characteristics make IL-15 an alternative choice for cancer immunotherapy. Because IL-15-IL-15RαS could further improve the biological functions of IL-15 (Fig. 2), together with its unique feature of low hepatic toxicity, we anticipate that this IL-15 superagonist might provide an effective and safe therapy for the treatment of patients with HCC.
Footnotes
Acknowledgments
The authors thank Dr. Nan-Shih Liao (Institute of Molecular Biology, Academia Sinica, Taiwan) for providing murine IL-15 and IL-15Rα cDNA. This work was supported by the Academia Sinica and by grant 98-3112-B-001-030 from the National Research Program for Genomic Medicine, National Science Council, Taiwan.
Author Disclosure Statement
All authors declare no conflict of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.