
Abstract
Mucopolysaccharidoses (MPSs) are lysosomal storage disorders characterized by progressive accumulation of glycosaminoglycans (GAGs) in various tissues. Enzyme replacement therapy (ERT) for several MPSs is available to date. However, the efficacy of ERT is limited, in particular in compartments such as bone, cartilage, the brain, and the eyes. We selected a rodent model of an MPS, with no central nervous system storage, to study the impact, on systemic features of the disease, of various stable levels of exogenous enzymes produced by adeno-associated viral vector (AAV)-mediated liver gene transfer. Low levels (6% of normal) of circulating enzyme were enough to reduce storage and inflammation in the visceral organs and to ameliorate skull abnormalities; intermediate levels (11% of normal) were required to reduce urinary GAG excretion; and high levels (≥50% of normal) rescued abnormalities of the long bones and motor activity. These data will be instrumental to design appropriate clinical protocols based on either enzyme or gene replacement therapy for MPS and to predict their impact on the pathological features of MPS.
Introduction
Clinical features vary within each MPS type, ranging from milder forms, compatible with a normal life span, to more severe types that are fatal during the first months of life (Neufeld and Muenzer, 2001).
Bone marrow transplantation (BMT) (Johnson and Orchard, 2002; Malatack et al., 2003) and enzyme replacement therapy (ERT) (Desnick and Schuchman, 2002; Rohrbach and Clarke, 2007) are the current treatments for several MPSs (Hoogerbrugge et al., 1995; Li et al., 1996; Gullingsrud et al., 1998; Rohrbach and Clarke, 2007; Wang et al., 2008), relying on the ability of lysosomal enzymes to be secreted and taken up by most cells via the mannose 6-phosphate receptor pathway (Dahms et al., 1989).
However, BMT is associated with substantial morbidity and mortality, and ERT with weekly infusions of costly recombinant enzymes. In addition, despite amelioration of the visceral phenotype because of the significant reduction in tissue GAG storage (Hoogerbrugge et al., 1995; Yano et al., 2009), both approaches show limited efficacy in ameliorating several MPS features, such as ocular pathology (Gullingsrud et al., 1998; Herskhovitz et al., 1999; Koseoglu et al., 2008; Pitz et al., 2009) and skeletal abnormalities (Field et al., 1994; Herskhovitz et al., 1999; Vellodi et al., 1999; Arora et al., 2007). Moreover, because lysosomal enzymes are not expected to cross the blood–brain barrier, both therapeutic approaches show limited efficacy (or no efficacy at all) concerning neurological manifestations of the disease (Hopwood et al., 1993; Shapiro et al., 1995; Vellodi et al., 1999; Desnick and Schuchman, 2002).
Few data are available on the levels of circulating enzymes in patients and animal models after BMT or during ERT. Serum half-lives, in the range of a few min/hours, have been reported for several lysosomal enzymes after intravenous infusion (Shull et al., 1994; Crawley et al., 1996; Jones et al., 1998; Harmatz et al., 2005; Tomatsu et al., 2008), with blood concentrations rapidly decreasing over time after each administration (Harmatz et al., 2005; Tomatsu et al., 2008), in association with enzyme uptake and persistence in peripheral tissues up to 1 week after infusion (Harmatz et al., 2005). BMT results in serum enzyme levels that are barely increased (Bergstrom et al., 1994; Vellodi et al., 1999); higher activities are rarely observed (Li et al., 1996). The low and/or transient serum enzyme levels achieved by these therapies may not be sufficient to ensure the delivery of enough enzyme to compartments such as the brain, bone, and cartilage (Desnick and Schuchman, 2002), or the eye (Pitz et al., 2009), which appears to be refractory to treatment.
Gene transfer to a “factory” organ, such as the liver, may provide high and stable levels of circulating lysosomal enzymes after a single, life-long vector administration. Expression of therapeutic levels of lysosomal enzymes from the liver and/or other target tissues, engineered via viral vector-mediated somatic gene transfer, has been reported in several MPS animal models (for a review, see Biffi and Naldini, 2005). This has resulted in amelioration of the MPS phenotype. Data collected in these studies suggest that a better therapeutic outcome is obtained when higher serum enzyme levels are achieved (Xu et al., 2002; Liu et al., 2005; Ma et al., 2007); however, a clear correlation between the levels of circulating enzymes and their effect on the various aspects of the MPS phenotype has not been established yet.
This would be helpful to improve the current therapeutic approaches for MPS as well as to improve our knowledge of the disease pathophysiology.
Given the inability of lysosomal enzymes to efficiently cross the blood–brain barrier, the impact of various serum enzyme levels on the various systemic aspects of MPS would be best evaluated in an MPS model with no central nervous system involvement. Indeed, in such a model, motor activity would be independent of CNS impairment.
Deficiency in the enzyme N-acetylgalactosamine-4-sulfatase (arylsulfatase B, ARSB) causes the autosomal recessive MPS VI (OMIM 253200), characterized by dermatan sulfate (DS) lysosomal storage (Neufeld and Muenzer, 2001); despite physical disabilities and a wide range of musculoskeletal problems, patients with MPS VI typically have normal intelligence (Hopwood and Morris, 1990; Neufeld and Muenzer, 2001).
We reported that adeno-associated viral (AAV) vector-mediated gene transfer to the liver of newborn MPS VI rats, bearing a null mutation in the ARSB gene, resulted in long-term ARSB secretion from transduced hepatocytes, after a single intravenous administration of vector (Tessitore et al., 2008). However, below-normal serum ARSB levels and limited therapeutic efficacy on the skeletal phenotype were observed. This was associated with the development of neutralizing humoral immune responses to the recombinant ARSB produced by hepatocytes transduced with AAV (Tessitore et al., 2008). Anti-ARSB antibodies developed independently of the transgene type used, presumably because MPS VI rats, bearing a null mutation, do not produce ARSB protein (Tessitore et al., 2008). This was unexpected because AAV-mediated, liver-restricted gene transfer has been reported to induce tolerance to secreted proteins in null animals (Mingozzi et al., 2003; Franco et al., 2005; Passini et al., 2007). Nevertheless, the nature of the transgene, as well as the specific disease model, might influence tolerance induction in this context.
Inhibition of immune response against the supplied enzyme has been reported to improve the therapeutic outcome in MPS (Dickson et al., 2008). Thus, to obtain higher levels of ARSB activity and to determine whether this could improve therapeutic efficacy in MPS VI rats, we tested various immunosuppressive (IS) drugs previously used to blunt the immune response (Fields et al., 2001; Herzog et al., 2001; Ponder et al., 2006; Mingozzi and High, 2007) and to help achieve long-term transgene expression. MPS VI rats were coadministered AAV and IS drugs, and levels of ARSB and their impact on various aspects of the MPS VI phenotype were evaluated.
Materials and Methods
Animal colony
MPS VI rats were obtained from T. Kunieda (Okayama University, Okayama, Japan) and maintained at the Cardarelli Hospital Animal House (Naples, Italy) (Tessitore et al., 2008).
Homozygous affected rats were crossed with wild-type Wistar rats (Harlan, S. Pietro al Natisone, Italy) to obtain heterozygous animals. Heterozygous rats, in turn, were bred to produce normal, heterozygous, and affected offspring. Genotype analysis was performed by polymerase chain reaction (PCR) on genomic DNA, as previously described (Kunieda et al., 1995), and the presence of the mutation was detected by sequencing (PRIMM, Naples, Italy).
AAV vector production and administration
Therapeutic AAV2/8-TBG-hARSB and control AAV2/8-TBG-eGFP vectors were produced by the AAV Vector Core of the Telethon Institute of Genetics and Medicine (TIGEM, Naples, Italy) by triple transfection of 293 cells and purified by passage through CsCl gradients (Xiao et al., 1999). Physical titers of the viral preparations (genome copies [GC] per milliliter) were determined by real-time PCR (Applied Biosystems, Foster City, CA) (Gao et al., 2000) and dot-blot analysis.
MPS VI rats were injected with 4 × 1013 GC of AAV2/8-TBG-hARSB or AAV2/8-TBG-eGFP vector per kilogram, either via the temporal vein (postnatal days 3 to 5 [P3 to P5]) or the femoral vein (P30).
Immunosuppressive drug administration
Doses and timing of IS administration are reported in Supplementary Table 1 (at
CTLA4-Ig (15 mg/kg; Bio X Cell, West Lebanon, NH) was administered intraperitoneally, alone or in combination with AAV vectors. Cyclosporine A (25 mg/kg; LC Laboratories, Woburn, MA) was injected intraperitoneally. Rapamycin (20 mg/kg; LC Laboratories) was administered via gavage; FK506 (2 mg/kg; LC Laboratories) was injected intraperitoneally; mycophenolate mofetil (MMF, 25 mg/kg) was coadministered with cyclosporine A, FK506, or rapamycin via gavage, with each IS drug administered according to the same schedule. Cyclosporine A and FK506 levels were measured by W. Utech (Department of Transplant Immunodiagnostics, Antonio Cardarelli Hospital, Naples, Italy). Rapamycin levels were measured by A. Gentile (Department of Clinical Pharmacology, Federico II University of Naples, Naples, Italy). All measurements were performed according to procedures used to dose drugs in clinical samples.
Blood and tissue collection
Blood was collected every month from treated and control rats via eye bleeding and centrifuged at 10,000 × g in an Eppendorf centrifuge for 10 min at 4°C to obtain serum.
The rats were killed at the end of the study (6–7 months after AAV administration) by cardiac phosphate-buffered saline (PBS) perfusion and the liver, kidney, spleen, and knee joints were collected.
Half of each tissue sample was fixed in methacarn solution (30% chloroform, 60% methanol, 10% acetic acid) for 24 hr and half was frozen in dry ice (for ARSB activity and GAG quantitative assays). Knee joints were fixed in 10% formalin (Sigma-Aldrich, Milan, Italy) for 24 hr.
ARSB activity assay and protein levels
Tissues were homogenized in water and protein concentrations were determined with the bicinchoninic acid (BCA) protein assay reagent (Pierce Protein Research Products/Thermo Fisher Scientific, Rockford, IL).
The ARSB assay was performed as described previously (Chang et al., 1981; Tessitore et al., 2008). Briefly, 30 μg of protein or 20 μl of serum were incubated with 40 μl of the fluorogenic substrate, 4-methylumbelliferyl sulfate (12.5 mM; Sigma-Aldrich), for 3 hr at 37°C in the presence of 40 μl of silver nitrate (0.75 mM; Carlo Erba, Milan, Italy). The reaction was stopped by adding 2 ml of carbonate stop buffer (0.5 M NaHCO3, 0.5 M Na2CO3; pH 10.7), and the fluorescence of the 4-methylumbelliferone liberated was measured with a fluorimeter (Bio-Rad, Milan, Italy) at 365 nm (excitation) and 460 nm (emission). Enzyme activities were calculated with a standard curve of the fluorogenic product, 4-methylumbelliferone (Sigma-Aldrich). For tissue lysates the activity is expressed as nanomoles per milligram of protein per hour. For serum samples, enzyme activity is calculated as the percentage of activity measured in serum from normal untreated (NR) rats in each assay (reported as 100%). Average serum ARSB is then calculated as an average of the percent activity measured for each rat every month.
ELISA for detection of anti-ARSB IgG antibodies
The presence of circulating antibodies against the hARSB protein in MPS VI rats was determined in a plate-binding assay, as described previously (Fazeli et al., 2006; Tessitore et al., 2008). Briefly, 96-well microtiter plates (Costar, Cambridge, UK) were coated with human ARSB (Naglazyme [galsulfase]; BioMarin Europe, London, UK), obtained from the Department of Pediatrics of the II Policlinico Hospital of Naples (Naples, Italy), in 100 μl of PBS overnight at 4°C. Plates were blocked for 2 hr at room temperature with 200 μl of 100% fetal bovine serum (FBS), washed five times with PBS containing 0.05% Tween 20 (washing buffer), and incubated with 100 μl of 1:50 dilutions of rat serum in PBS–0.05% Tween–10% FBS for 2 hr at room temperature. The washing step was repeated and 100 μl of biotinylated anti-rat IgG antibody (Vector Laboratories, Peterborough, UK) in PBS–0.05% Tween–10% FBS was added and incubated for 1 hr at room temperature. Binding was revealed by the addition of ExtrAvidin–horseradish peroxidase (Sigma-Aldrich) followed by o-phenylenediamine dihydrochloride (OPD) substrate (Sigma-Aldrich). Absorbance at 450 nm (optical density) was determined with a Titertek Multiskan ELISA reader (Titertek Instruments, Huntsville, AL). The optical density, determined in an assay, represented a qualitative measure of antibody affinity. The assay sensitivity level was defined as the average optical density measured in animals that never received AAV2/8-TBG-hARSB vector.
Quantitative analysis of GAG accumulation in tissues and urine
Urine samples from treated and control rats were collected in metabolic cages every month after treatment and were diluted in water 10-fold for measurement of GAG content.
Fifty microliters of diluted urine or 250 μg of protein extract from liver, spleen, and kidney were used for the GAG assay, as previously described (de Jong et al., 1989; Tessitore et al., 2008). GAG concentrations were determined on the basis of a dermatan sulfate standard curve (Sigma-Aldrich). Tissue GAGs are expressed as micrograms of GAG per milligram of protein. Urine GAGs were normalized to creatinine content. Creatinine was measured with a creatinine assay kit (Quidel, San Diego, CA). Urinary GAGs are expressed as micrograms of GAG per micromole of creatinine and are reported as an average value measured each month for each rat.
Alcian blue staining and immunohistochemistry
After methacarn fixation, all the tissues (liver, spleen, kidney, and heart) were embedded in paraffin and sectioned into 7-μm-thick serial sections on a microtome. For the GAG assay, tissue sections were rehydrated and stained with 1% Alcian blue (Sigma-Aldrich) in hydrochloric acid. Counterstaining was performed for 2 min with nuclear-fast red (Sigma-Aldrich). Heart valve storage was assessed in the mitral, aortic, and tricuspid valves.
For CD68 staining, sections were incubated for 1 hr with blocking solution (1 × PBS, 0.5% Tween 20, 0.1% bovine serum albumin, and 10% fetal bovine serum; GIBCO-BRL/Invitrogen, Gaithersburg, MD) and incubated overnight with a polyclonal anti-CD68 antibody (diluted 1:300; Serotec, Oxford, UK). After washing, sections were incubated for 1 hr with biotinylated secondary anti-rabbit IgG (Vector Laboratories, Burlingame, CA). The reaction was developed with a VECTASTAIN Elite ABC-peroxidase kit (Vector Laboratories) followed by 30 min of 3,3′-diaminobenzidine (DAB) staining (Vector Laboratories). Last, sections were counterstained with hematoxylin (Sigma-Aldrich) and mounted with Eukitt (Kaltek, Padua, Italy).
Radiograph analyses
Skull radiographs of 6-month-old treated and control MPS VI rats were taken with a 9600 mobile imaging system (OEC Medical Systems, Milan, Italy). Radiographs were printed, the condylobasal length and the maximal width were measured, and their ratio was calculated.
For assessment of long bone length, radiographs of 6-month-old rats were taken with an Acer X-ray machine at the veterinary clinic L'Arca (Naples, Italy); lengths of the femur, tibia/fibula, humerus, and radius/ulna were measured on radiograph films with a precision caliper (Mitutoyo, Milan, Italy) and reported as a percentage of the length of each bone in normal rats.
Bone histology and immunohistochemistry
Formalin-fixed knee joints were decalcified in 8% formic acid (Sigma-Aldrich), dehydrated, embedded in paraffin, and sectioned into 7-μm sections. For histological analysis, sections were stained with hematoxylin and eosin (Richard-Allen Scientific, Kalamazoo, MI; Sigma-Aldrich) according to standard procedures. For CD68 staining, the sections were rehydrated, digested for 1 hr at room temperature with 0.05% hyaluronidase (Sigma-Aldrich), and stained as reported for the other tissues.
18F Positron emission tomography
[18F]fluoride was produced with an in-house Scanditronix MC17 cyclotron (Scanditronix, Uppsala, Sweden). Five-month-old treated and control MPS VI rats, as well as normal controls, were anesthetized with 2,2,2-tribromoethanol (Avertin), and 20–80 MBq of [18F]fluoride were injected into the femoral vein. After 20 min, the animals were scanned under anesthesia in an ECAT47 clinical PET scanner (Siemens, Erlangen, Germany) in two-dimensional mode with measured attenuation correction at the Department of Nuclear Medicine of the National Institute for Tumors, G. Pascale Foundation (Naples, Italy). Scans were analyzed with e.soft software (version 5.0.3; Siemens) and standardized uptake values (SUVs) were measured by placing three-dimensional regions of interest (ROIs) on both knee joints of each rat. Results are reported as the average maximal SUV measured in the right and left knee joints from each rat, expressed as a percentage of the SUVs measured in normal rats. ROI analysis was also attempted on other segments, such as the spine and the elbow joint, but since measurements in these segments were variable the knee joint was chosen as the reference region for quantitative analysis.
Analysis of motor activity
A total of 63 (39 females and 24 males) adult rats (5 to 6 months old) were used for the behavioral analysis. The behavioral testing room was exposed to constant sound (classical music) and light conditions. Animals were tested during the day between 9.00 am to 6.00 pm. Before each behavioral task, animals were acclimatized to the testing room for at least 30 min.
Motor activity was tested by using the rota-rod apparatus for rats (Ugo Basile, Comerio, Italy) with four rods. On the first day each rat was gently placed on each rod set at a steady slow speed of 4 rpm and trained to remain on the rod for 60 sec. After this habituation trial each animal was exposed to four trials per day for five consecutive days, with an intertrial interval of 15 min. The trial started when all rats moved in the correct direction. The rota-rod was set at increasing speeds ranging from 5 to 40 rpm over 5 min (Monville et al., 2006) and the animals were left on the rod for an additional 3 min. The latency to fall off the rod within this period was recorded.
The latency to fall off the rod was averaged for the four trials and the five training days, and analyzed by one-way analysis of variance (ANOVA) with the variable group (11 levels) as the between-group factor. Correlation between percentage of ARSB (average of three samples analyzed in the last 3 months before the behavioral measurement) and mean latency to fall off the rod was made, using simple regression.
Results and Discussion
High levels of systemic ARSB are obtained by combining immunosuppression with AAV-mediated gene delivery in MPS VI rats
To inhibit the generation of systemic anti-ARSB antibodies after AAV delivery (Tessitore et al., 2008) and to obtain high ARSB serum levels in MPS VI rats, we used IS drugs in combination with somatic gene transfer. MPS VI rats were administered systemically AAV2/8 vectors carrying the human ARSB transgene (hARSB) under the control of the liver-specific thyroxin-binding globulin (TBG) promoter (AAV2/8-TBG-hARSB) at 4 × 1013 genome copies/kg (GC/kg), a dose that has proven to be effective from a previous study (Tessitore et al., 2008). The IS drugs used included the following: the recombinant cytotoxic T lymphocyte antigen 4-immunoglobulin (CTLA4-Ig) fusion molecule (Ponder et al., 2006), which is a costimulation blocker preventing T cell activation (Graca, 2008); and combinations of either cyclosporine A (CsA) (Fields et al., 2001), tacrolimus (FK506) (Fields et al., 2001), or rapamycin (Rap) (Miao et al., 2006) with mycophenolate mofetil (MMF) (Ciron et al., 2006), conventionally used to inhibit lymphocyte proliferation. We selected two ages for treatment: newborn (P3 to P5, indicated by an “n” in front of the group acronym) or juvenile (P30, indicated by a “j” in front of the group acronym). Juvenile animals were included to assess the efficacy of treatment at later stages of the disease and to investigate the efficacy of gene transfer with AAV (the genome of which is mainly in an episomal state in the nucleus of target cells) in the juvenile liver, in which hepatocyte proliferation is lower than in the newborn liver. In addition, the two ages selected, newborn and juvenile, broadly represent the two main ages at which a diagnosis is made in human patients, either because of the appearance of clinical signs or because of a positive family history or prenatal diagnosis. The IS regimen we selected successfully inhibited immune responses to the transgene in gene therapy protocols (Fields et al., 2001; Ciron et al., 2006; Miao et al., 2006; Ponder et al., 2006). In a pilot study we evaluated the toxicity and bioavailability of this regimen in wild-type newborn and juvenile rats; circulating IS levels were measured in serum samples at the time points reported in Supplementary Table 1. IS drug serum levels were found to be in the range corresponding to the human therapeutic range (see Supplementary Table 1). CTLA4-Ig was the only drug that was well tolerated by newborn rats; therefore, although all IS drugs were coadministered with AAV in juvenile MPS VI rats, only CTLA4-Ig was administered to newborns. Control animals included the following: (1) normal untreated rats (NR), (2) MPS VI rats untreated or treated with AAV2/8-TBG-EGFP (AF), and (3) affected newborn and juvenile rats treated with CTLA4-Ig alone (AF + CTLA4-Ig). Serum ARSB activity levels and circulating anti-hARSB antibodies were measured each month for 6–7 months, after which animals were killed for ex vivo analysis of harvested tissues.
Animals treated with AAV alone as newborns (n-AAV) had significantly higher ARSB levels in serum than did AF animals (Fig. 1a). These levels were on average 11% of NR and reached peaks of 30% (Fig. 1a). The decline in serum ARSB activity over time may be caused by vector dilution due to hepatocyte proliferation occurring in newborns, which is less pronounced in juvenile rats. Rats from the j-AAV group showed only a slight increase in serum ARSB levels (on average, 6% of NR) compared with AF animals (Fig. 1b). Serum neutralizing antibodies (NAb) were concordantly higher in j- than in n-AAV animals (Fig. 1c and d). When IS drugs were coadministered with AAV, serum ARSB levels increased in some but not all treated animals, to levels higher than in animals treated with AAV alone. More specifically, in two of nine n-AAV-CTLA4-Ig (Fig. 1a) serum ARSB levels were on average >60%, peaking up to 75% of NR levels. Similarly, in one of five j-AAV-CTLA4-Ig, one of five j-AAV-FK506, and three of five j-AAV-CsA, circulating ARSB levels were between 50 and 76% of NR levels, and remained stable for the duration of the study (Fig. 1b). The remaining animals in the groups administered AAV + IS drugs had serum ARSB and anti-ARSB antibody levels comparable to those of age-matched littermates treated with AAV alone (data not shown). In the animals in which coadministration of IS drugs with AAV had resulted in high serum ARSB levels, serum NAb levels were concordantly lower than in animals receiving AAV alone (Fig. 1c and d), or AAV + IS drugs, with serum ARSB levels ≤50% of NR (data not shown). Notably, all animals receiving AAV and Rap in combination with MMF on P30 had low serum ARSB levels comparable to those of j-AAV (data not shown). As expected, AF + CTLA4-Ig, whether administered at birth or on P30, had serum ARSB levels comparable to those of age-matched AF littermates (data not shown). We do not have a clear explanation for the variability we observed among AAV animals treated with immunosuppressive drugs both in serum enzyme and NAb levels. One possibility is the heterogeneity in the humoral immune response against the transgene, as suggested for FVIII (Miao et al., 2006); in addition, the mixed genetic background (Ishibashi Hairless [Yoshida et al., 1993] × Wistar) of MPS VI rats may have contributed to the variability observed; further, the IS doses we used may be subtherapeutic, although similar doses were successfully used in other animal models (Fields et al., 2001; Ciron et al., 2006; Miao et al., 2006; Ponder et al., 2006) and IS serum levels in treated rats were maintained in a therapeutic range (Supplementary Table 1) (Napoli et al., 1997; Jacobson et al., 2001; Podder et al., 2001; Maury et al., 2002; Sakai et al., 2004; Kuypers, 2005). Although the combination of CsA and MMF seemed to result in high circulating ARSB levels more consistently than with any other regimen tested, the suggestion of a specific IS regimen to be combined with AAV2/8-mediated liver gene transfer for therapy of MPS VI is beyond the aim of this study and is not possible because of the high variability we observed in IS efficacy.

Immunosuppression results in high levels of circulating arylsulfatase B (ARSB) and low levels of anti-ARSB antibodies. Mucopolysaccharidosis type VI (MPS VI) rats were injected with AAV2/8-TBG-hARSB vector with or without immunosuppressive (IS) drugs at birth (
Taken together, these results suggest that reduction of humoral immune responses to hARSB results in stable circulating ARSB levels higher than 50% of those observed in NR rats. This demonstrates that the levels of AAV-mediated ARSB transduction and serum enzyme activity were influenced by the immune response and that this can be attenuated by immunosuppression, both in newborn and young animals.
The different enzyme levels observed among treated animals correlate with levels of anti-ARSB antibodies measured: MPS VI rats with the highest serum enzyme levels had low circulating anti-ARSB antibodies and vice versa. Indeed, the presence of serum antibodies impacts on mannose 6-phosphate receptor-mediated enzyme uptake and on enzyme tissue distribution and clearance, both mediated by the Fc receptors present on tissue macrophages, which are specifically abundant in reticuloendothelial tissues (Brooks et al., 1997, 1998; Dickson et al., 2008). Therefore, the levels of enzymatic activity measured in the presence of anti-ARSB antibodies should already reflect the potential decrease caused by the immune response. However, we cannot exclude that similar enzyme levels may result in different therapeutic outcomes based on the presence or absence of circulating anti-ARSB antibodies.
Serum ARSB levels as low as 6% of normal are sufficient to reduce MPS VI visceral storage and inflammation
The use of AAV, whether in combination with IS drugs or not, and in the different age classes of treatment, resulted in different ARSB serum levels in MPS VI rats. We studied the impact of the aforementioned therapeutic regimen on the various MPS VI phenotypic characters. To this end, we identified three different groups of animals on the basis of their average serum ARSB levels: (1) j-AAV rats (n = 6), showing the lowest enzyme serum activity (on average, 6% of NR); (2) n-AAV rats (n = 7), in which we observed intermediate levels (on average, 11% of NR); and (3) AAV + IS rats (n = 7), which included animals receiving AAV + IS drugs treated either at birth or on P30, with serum ARSB activity between 50 and 76% of NR levels (Fig. 2a). This grouping, albeit combining animals with different treatments and similar serum enzyme levels, allows to assess the impact of serum enzyme levels on the various MPS VI biochemical, pathological, and skeletal features.

Impact of various serum ARSB levels on urinary glycosaminoglycan (GAG) excretion. Average serum ARSB activity (percentage of NR levels;
Treated animals were killed at 6–7 months of age and ARSB activity was measured in the liver, spleen, and kidney (Table 1). Animals with the highest circulating ARSB activity had the highest enzyme activity levels in the liver as well as in untransduced tissues such as the kidney and spleen (p ≤ 0.01 vs. n- and j-AAV).
Abbreviations: AAV+IS, rats receiving both AAV vectors and immunosuppressive (IS) drugs showing serum ARSB activity ≥ 50% of NR; AF, MPS VI-affected rats; ARSB, arylsulfatase B; j-AAV, rats receiving AAV vectors on P30; MPS VI, mucopolysaccharidosis type VI; n, number of rats analyzed for each group; n-AAV, rats receiving AAV vectors at birth; NR, normal untreated rats.
All tissue measurements were taken at the time of sacrifice (6–7 months).
Means ± SEM.
We assessed GAG accumulation in both tissues and urine of MPS VI rats from each of the three groups showing different systemic ARSB levels, as well as from controls (Figs. 2b and 3). Tissue GAGs were analyzed in the liver, spleen, kidney, and heart of MPS VI rats at the time of sacrifice by both Alcian blue staining of tissue sections and the quantitative dimethylmethylene blue method (Fig. 3a). All AAV-treated animals had liver, spleen, and kidney GAG levels lower than those of AF animals, with the exception of the AAV + IS group, for which GAG levels were comparable to those under normal conditions (p ≥ 0.3). Interestingly, GAG accumulation was evident in AF heart valves (Fig. 3a) but almost absent in the myocardium (data not shown), as previously reported (Yoshida et al., 1993). Heart valve storage in n- and j-AAV rats was similar to that in AF rats whereas it was cleared in AAV + IS animals. Poor heart valve vascularization (Sacks et al., 2009) may explain the requirement for high circulating ARSB activity levels for GAG clearance.
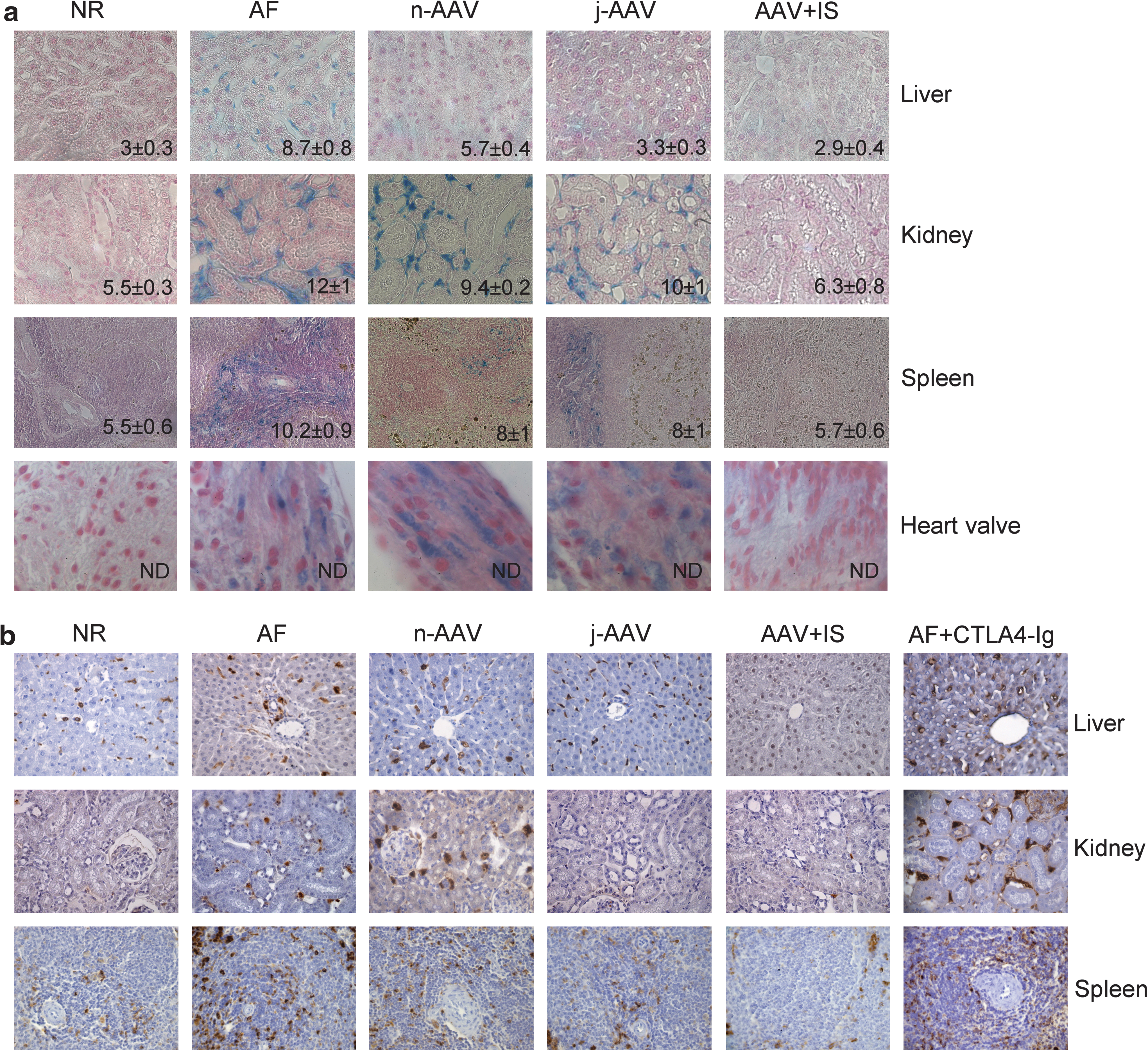
Tissue GAG levels and macrophage infiltration are inversely correlated with serum ARSB levels. (
Tissue inflammation, which is associated with ARSB deficiency and GAG accumulation (Tessitore et al., 2008) and is revealed by the presence of activated CD68+ macrophages, was reduced in j-AAV and n-AAV animals and almost abolished in rats treated with AAV + IS (Fig. 3b). j-AAV-CsA animals showed persistent CD68 staining independently of circulating ARSB levels (data not shown), probably due to CsA side effects (Pernu and Knuuttila, 2001; Ghee et al., 2008). Importantly, in AF animals receiving CTLA4-Ig alone, either at birth or on P30 (AF + CTLA4-Ig), tissue GAG accumulation (GAG levels: in the liver, 7.9 ± 0.3; in the kidney, 11.7 ± 0.7; in the spleen, 11.4 ± 0.3) and macrophage infiltration were similar to those observed in AF animals (Fig. 3b). The average urinary GAG levels measured during the study were inversely correlated with circulating ARSB activities, as expected (Fig. 2b). AF or j-AAV rats had significantly higher urinary GAGs than NR, followed by n-AAV animals and AAV + IS rats, the latter being the only group with urinary GAG levels close to NR and with serum ARSB levels ≥50% of NR.
These results suggest that circulating ARSB activity levels below 11% of normal, such as those obtained in the n-AAV and j-AAV groups, result in low enzyme uptake by untransduced tissues, which is sufficient to reduce GAG storage and inflammation, as previously reported (Tessitore et al., 2008). Nevertheless, systemic enzyme levels ≥50% of NR, such as those obtained in the AAV + IS animals, result in higher peripheral enzyme uptake and clearance of GAG storage and of inflammatory infiltrates.
Levels of systemic ARSB ≥50% of normal are required to rescue abnormalities of the long bones and motor activity in MPS VI rats
One of the major challenges of therapies for MPS VI, as well as for other MPSs, is to improve skeletal and joint abnormalities. To this end, the availability of enzymes for cartilage and bone is critical and often limited by the local microcirculation. Like MPS VI patients, MPS VI rats have a widened and shortened cranium and shortened long bones (Tessitore et al., 2008).
To assess the impact of ARSB serum levels on the MPS VI skeletal phenotype, we performed radiographic analyses of the skull and long bones of 6-month-old animals. Skull abnormalities were assessed by measuring the ratio between the condylobasal length (L) and the maximal zygomatic width (W) (Tessitore et al., 2008) whereas the length of the long bones (tibia, femur, humerus, and ulna/radius) was measured and reported as the percentage of NR bone length (Fig. 4). The L/W ratio, which is significantly reduced in AF animals compared with NR age-matched controls, was improved in n- and j-AAV rats (with serum ARSB levels on average as low as 6% of NR) and rescued in AAV + IS animals (Fig. 4a). In contrast to what was observed for the skull, only AAV + IS rats with serum ARSB levels ≥50% of NR presented complete rescue of femur length (Fig. 4b) as well as of all the other long bones analyzed (data not shown). The different systemic ARSB levels associated with improvement of the skull and long bone abnormalities could be related to more efficient diffusion of circulating ARSB in flat (skull) compared with long bones. Importantly, we observed a similar rescue of the skeletal phenotype in rats treated at birth or on P30, with the extent of rescue exclusively correlating with serum ARSB activity levels and not with the age at which treatment was started. This could be expected because we observed by radiographic analysis that the long bone length in NR rats increased steadily until 7 months of age, which corresponds to the last time point in our analysis (data not shown). The skeletal rescue observed in j-AAV rats suggests that improvement of skeletal abnormalities, in individuals treated before skeletal growth is complete, may be feasible.
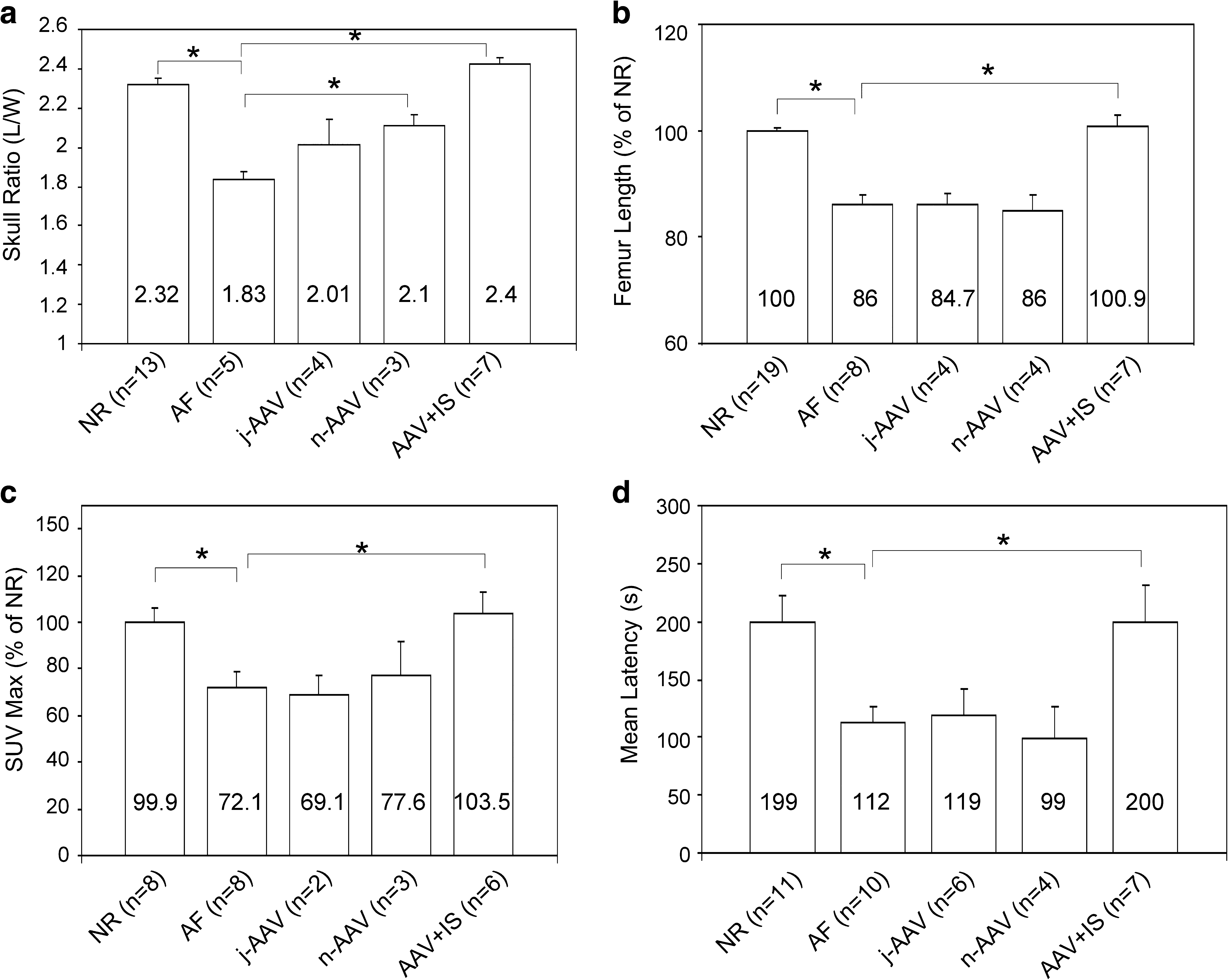
High ARSB levels are required to rescue skeletal abnormalities and motor activity. (
Studies aimed at explaining the basis for bone pathology in MPS VI rats have suggested that defects in endochondral ossification are involved in the reduced growth of long bones (Nuttall et al., 1999; Simonaro et al., 2005). This could be related to defective chondrocyte differentiation (Simonaro et al., 2005) and osteoblast activity in bone metaphysis (Nuttall et al., 1999), a site where chondrocytes and osteoblasts cooperate for deposition of new bone tissue (Bloom and Fawcett, 1994; Nuttall et al., 1999). To assess the impact of gene transfer on endochondral ossification, we used positron emission tomography (PET) with positron-emitting [18F]fluoride to measure osteoblast activity in the knee joint of MPS VI and control rats (Hsu et al., 2007). [18F]fluoride has high affinity for hydroxyapatite (HA) and is captured in regions of high osteoblast activity, where HA concentrates in the newly produced bone matrix (Hsu et al., 2007). Qualitative assessment of [18F]fluoride standardized uptake values (SUVs, expressed as a percentage of NR) was obtained by region of interest (ROI) analysis of the knee joint, which was chosen as the reference region because it yielded the best reproducibility of measurements. MPS VI rats showed significantly reduced SUV values compared with NR rats (Fig. 4c), suggesting that defects in endochondral ossification and reduced osteoblast activity are responsible, at least in part, for MPS VI bone abnormalities. These findings were confirmed in other skeletal segments by visual analysis, although quantitative analysis in other segments such as the spine and elbow could not be performed given the variability observed in these regions. Although n-AAV and j-AAV rats showed no improvement in the SUV value of the knee joints, this was on average normalized in rats treated with AAV + IS (Fig. 4c; and see Supplementary Fig. 1 at
To analyze GAG storage in the bones and cartilage of MPS VI rats, we examined hematoxylin and eosin (H&E)-stained histological sections of knee joints from 6- to 7-month-old MPS VI rats (Fig. 5). These sections showed extensive vacuolization, representing lysosomal storage, in AF cortical bone osteocytes and in chondrocytes of the articular surface of the tibia (Fig. 5a). Vacuolated chondrocytes were present within the full thickness of the AF tibial growth plate (Fig. 5a), whereas in NR rats chondrocytes of increased size are typically present only in the hypertrophic chondrocyte region. In addition, loss of the normal columnar organization of chondrocytes and disorganization of the tibial growth plate (GP) were evident in AF joints, as previously reported (Abreu et al., 1995; Evers et al., 1996). Animals in the n- and j-AAV groups had histological knee joint abnormalities similar to AF rats (data not shown) whereas the AAV + IS animals showed clearance of osteocyte and chondrocyte storage as well as a normal GP structure. Similar results were observed in femurs from the same animals (data not shown). Because production of proinflammatory cytokines has been reported in the cartilage and bone of MPS VI animals (Simonaro et al., 2005), we investigated the presence of activated macrophages in the aforementioned regions of MPS VI rats by immunohistochemical analysis of CD68, which is also expressed by osteoclasts (Kaneko et al., 2001). As shown in Fig. 5b, AF but not NR rats had CD68 immunostaining in the subchondral bone and primary spongiosa of the knee joint, suggesting that activated macrophages and/or osteoclasts are present in these regions and could be involved in the production of proinflammatory cytokines eventually leading to cartilage/bone erosion (Kaneko et al., 2001). Although no differences in number of CD68+ cells were detected between AF animals, treated with CTLA4-Ig (AF + CTLA4-Ig; Fig. 5b) or with AAV alone (j-AAV and n-AAV; data not shown), those treated with AAV and IS had reduced or absent CD68 staining (Fig. 5b). Interestingly, j-AAV-CsA with high levels of circulating ARSB, despite complete normalization of GP structure and of cell vacuolization, showed persistent CD68 staining (data not shown). CsA administration has been documented to increase the number of osteoclasts on the bone surface (Spolidorio et al., 2007; Nassar et al., 2008) and to induce CD68 expression on macrophage cell lines (Spolidorio et al., 2007; Nassar et al., 2008). Thus, the persistent CD68 staining in bones and tissues of j-AAV-CsA-treated rats may be related to CsA side effects.
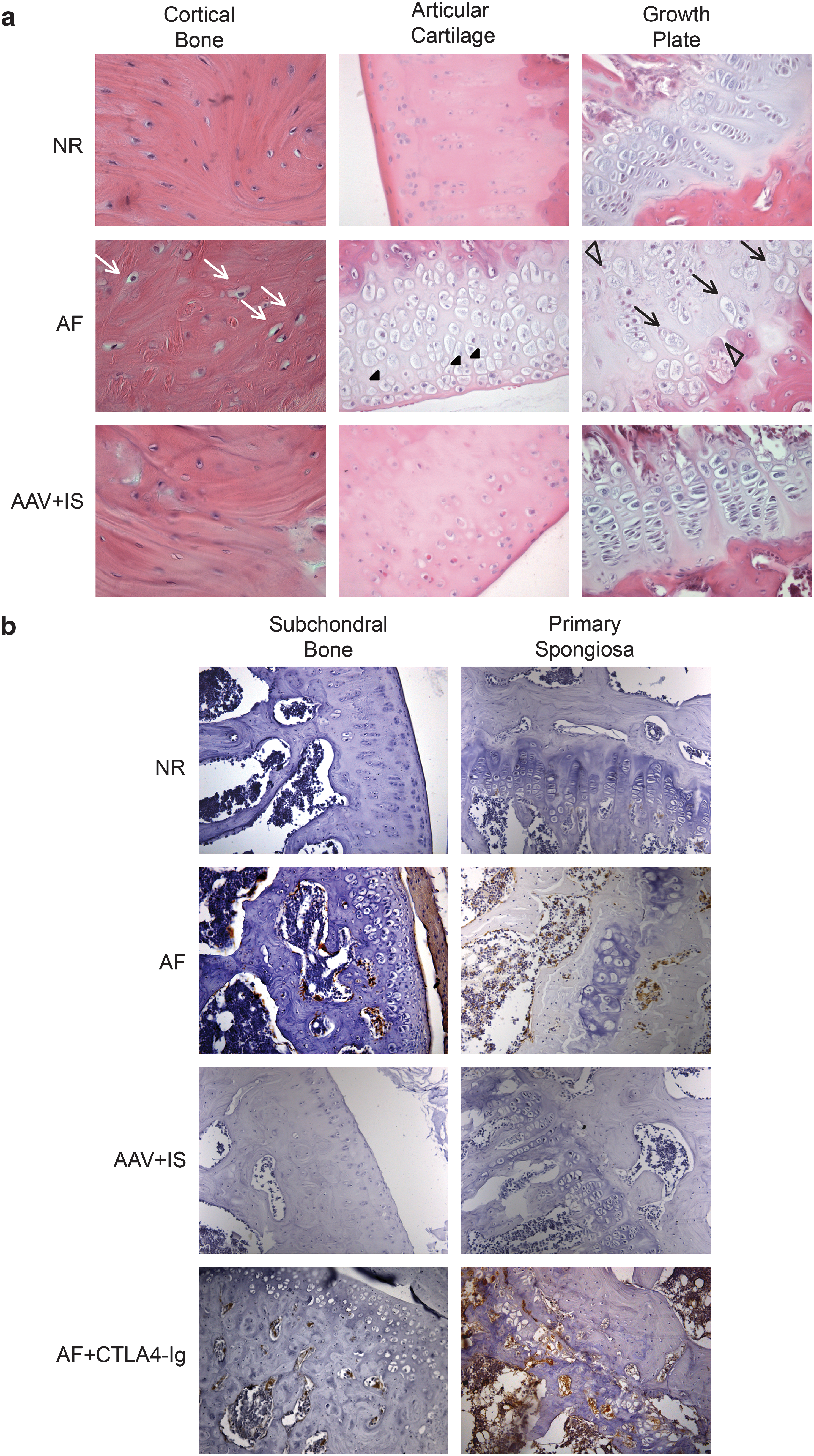
Cartilage and bone abnormalities are rescued in MPS VI animals with high ARSB serum levels. (
These data suggest that a significant amelioration of skeletal abnormalities, including osteoblast activity and long bone length, reduction of inflammation, and improvement in bone and cartilage histology, requires levels of circulating ARSB ≥50% of NR.
Bone and joint abnormalities in MPS VI patients are the cause of motor disabilities. Therefore, we hypothesized that MPS VI rats could have impaired motor activity. To test this we used the rota-rod motor coordination task that forces animals to walk on a high accelerating rod. Motor performance is measured as the latency to fall off the rod. Fisher's PLSD post-hoc analysis demonstrated that the performance of AF animals was significantly (p = 0.006) worse than that of NR rats, therefore demonstrating that MPS VI rats have impaired motor activity (Fig. 4d). The latency to fall off the rod of n-AAV and j-AAV animals was similar to that of AF animals. Interestingly, AAV + IS animals displayed a behavioral phenotype comparable to that of NR animals (Fig. 4d). Rescue of the behavioral phenotype was presumably not due to IS injection per se, as AF + CTLA4-Ig rats showed no improvement in rota-rod tests (data not shown). To confirm that the rescue we observed was correlated with the amount of circulating ARSB, we calculated the linear regression between the percentage of ARSB and the mean latency to fall off the rod. A highly significant correlation (p < 0.0001) was found between the two variables. Within the AAV + IS group, the j-AAV-CsA animals, despite levels of circulating ARSB ≥ 50% of NR, had motor performance that was worse than that of the other AAV + IS animals (data not shown). Interestingly, this was the only group that, despite a normalization of the skeletal parameters, had persistent CD68+ cells, presumably a side effect of CsA administration.
Altogether, these results demonstrate that bone and joint abnormalities in MPS VI rats are accompanied by motor disabilities and that these, in turn, are rescued when the amount of circulating ARSB in affected rats is ≥50% of NR. Importantly, MPS VI represents an ideal model disease in which to study the impact of the various serum ARSB activity levels obtained in therapeutic settings on motor abnormalities. The absence of CNS involvement in MPS VI disease allows direct correlation of serum enzyme activity levels with motor performance, avoiding the bias due to the influence of CNS function impairments, which are evident in other MPS models (Garcia-Rivera et al., 2007; Heldermon et al., 2007).
In conclusion, this study produced a range of circulating ARSB levels in MPS VI rats, from 4 to 76% of NR, allowing evaluation of the impact of serum ARSB levels on various MPS VI pathological parameters.
These data, as summarized in Fig. 6, show that average ARSB levels as low as 6% of those observed in NR animals are sufficient to reduce the visceral lesions in MPS VI rats and to improve skull abnormalities. Intermediate levels (≥11% of NR) result in reduction of urinary GAG storage whereas high levels (≥50% of NR) are required for normalization of heart valve storage, and rescue of the abnormalities of the long bones and motor activity.
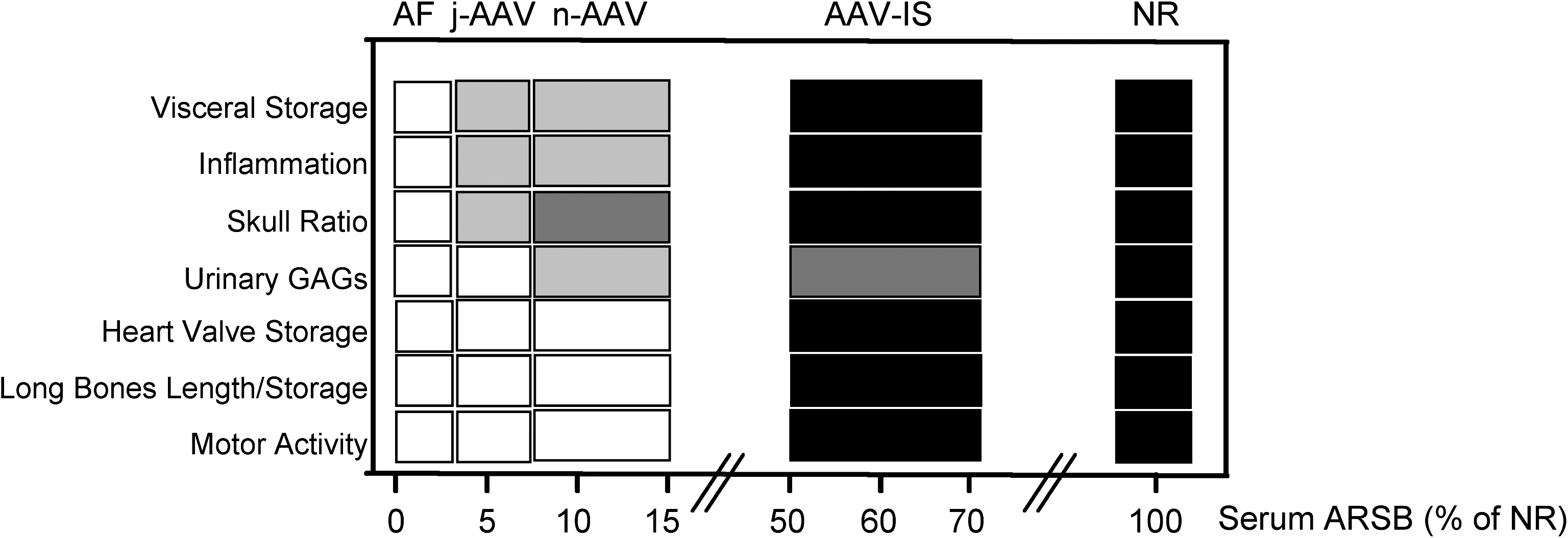
Different serum ARSB levels are required to improve different MPS VI phenotypic aspects. Shown is the correlation between improvement of the various MPS VI systemic features observed and the ARSB serum levels in control and treated MPS VI rats. The different pathological aspects analyzed are reported on the left and serum ARSB levels are reported at the bottom, as a percentage of normal (NR) levels. Affected (AF), NR, and MPS VI rats receiving either AAV alone at birth (n-AAV) and on P30 (j-AAV) or receiving AAV with various IS drugs (AAV + IS) are depicted at the top. Shading (from white to black) is correlated with the level of phenotype improvement. White, no improvement; light gray, a minor improvement; dark gray, a significant improvement; black, the normal phenotype (NR) or the complete rescue of the phenotype (AAV-IS).
Because MPSs, and lysosomal storage diseases in general, share a similar systemic phenotype, including skeletal abnormalities, our results have important implications for the therapies of these diseases, either based on gene or enzyme replacement, suggesting that high stable enzymatic levels are required for complete phenotype rescue and indicating that monitoring serum enzyme levels during therapy could be important to predict therapeutic outcome.
Footnotes
Acknowledgments
The authors are grateful to Drs. Mark Haskins, Giancarlo Parenti, Graciana Diez-Roux, Carmine Settembre, and Nicola Brunetti-Pierri for helpful discussion and for critical revision of the manuscript. Drs. Calogera Simonaro and Francesca Abramo gave useful advice on bone histology. The authors thank the AAV Vector Core for production of the vectors used in this study and the Comparative Pathology Technological Platform, CEINGE (Naples, Italy), for help with tissue embedding. This work was supported by Telethon grant TIGEM P33, grant PRIN 2006064337 from the Italian Ministry of University and Research, grant Regione Campania L.R. n. 5/02, and by the European Union within its Sixth (DiMI contract 512146; Clinigene contract 018933) and Seventh (EUCLYD contract 201678) Framework Programs.
Author Disclosure Statement
The authors do not declare conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.