
Abstract
Melanoma differentiation-associated gene-7/interleukin-24 (mda-7/IL-24), identified by subtraction hybridization in the mid-1990s, is a potent gene therapeutic for cancer. Using a replication-deficient adenovirus as vector, it provokes apoptosis in diverse cancer cells without harming normal cells or tissues. To exploit the anticancer capability of IL-24 to the best, in this study, we generated a novel gene-virotherapy agent MUD55-IL-24, utilizing a replication-competent oncolytic adenovirus MUD55 as the gene delivery vector. It was documented that MUD55-IL-24 exhibited much stronger antitumor activity on gastric carcinoma both in vitro and in vivo, and its safety was comparable to the replication-deficient adenovirus Ad-IL-24. The unique properties of IL-24, including apoptosis induction, antiangiogenesis, and antimigration, were all significantly enhanced in MUD55-IL-24. After looking into the underlying mechanism, we found that intracellular ROS (Reactive Oxygen Species) generation may have caused the induction of apoptosis, mitochondrial dysfunction, and the activation of caspases in MUD55-IL-24-infected SG-7901 cells. Taken together, these results suggest that MUD55-IL-24 may be able to provide a potential strategy for future treatment of human gastric carcinoma.
Introduction
Gene therapy is one of the hot topics in cancer treatment. Scientific breakthrough in our understanding of the important genes in cancers has endued it great promise. The effect of gene therapy depends on the efficiency of therapeutic genes and gene delivery vehicle. Melanoma differentiation-associated gene 7/interleukin-24 (mda-7/IL-24) is a secreted cytokine that was identified by subtractive hybridization in the mid-1990s. It is not expressed or presents at very low levels in tumor cells compared with nontransformed cells (Ekmekcioglu et al., 2001). IL-24 represents considerable therapeutic potential for its ability to suppress cell growth and induce cell death in a broad spectrum of cancers independent of the status of p53, pRB, p21, and additional tumor suppressor genes (Jiang et al., 1996; Su et al., 1998, 2001; Madireddi et al., 2000; Saeki et al., 2000; Lebedeva et al., 2002), without obvious cytotoxicity in normal cells (Lebedeva et al., 2003; Sauane et al., 2004). Besides, IL-24 has antiangiogenesis, radiosensitizing, and immunological enhancement effects (Kawabe et al., 2002; Fisher et al., 2003; Yacoub et al., 2003; Sarkar et al., 2005). It has shown potent antitumor properties in vitro, in vivo, and in phase I clinical trial (Cunningham et al., 2005; Tong et al., 2005; Eager et al., 2008; Xue et al., 2008).
There are many vectors, including viruses and nonviral agents, and methods for gene therapy. Adenovirus is one of the most widely studied vectors and shows a lot of virtues. It is more efficient in gene transfection than nonviral methods, and safer than other viruses, such as retrovirus, because the foreign gene will not be inserted into the genomic DNA of the host cell. Replication-deficient adenovirus has been commonly chosen for gene therapy in the past. Recently, however, oncolytic adenoviruses, which are replication competent, have also been used and showed better gene delivery efficiency, because the foreign gene can replicate with the viral DNA specifically in the target tumor cells. Besides, it combines the antitumor effect of the viral vector and the therapeutic gene together.
Previously, we have utilized this strategy to construct the gene-virotherapy agent ZD55-IL-24, which exhibited potential antitumor activity on colorectal cancer. In this study, we used a double-regulated oncolytic adenovirus MUD55 as the gene delivery vehicle and generated a novel agent MUD55-IL-24. MUD55 has been proved to be a safer adenovirus targeting adenocarcinomas (Wei et al., 2009). As 85% cases of gastric cancer are adenocarcinomas that occur in the lining of the stomach, MUD55-IL-24 is expected to have potent antitumor efficacy on gastric carcinoma.
Materials and Methods
Cell culture
The human embryonic kidney cell line HEK293 was obtained from Microbix Biosystems, Inc. (Toronto, Canada). SG-7901 (human gastric cancer cell line) and MRC-5 (human lung fibroblast cell line) were obtained from the Institute of Biochemistry and Cell Biology (Shanghai, China). All these cells were cultured in RPMI 1640 or Dulbecco's modified Eagle's medium (Gibco-BRL, Grand Island, NY) supplemented with 10% heat-inactivated fetal bovine serum (Gibco-BRL), 4 mM glutamine, 50 U/ml penicillin, and 50 μg/ml streptomycin at 37°C in a humidified air atmosphere with 5% CO2. Human umbilical vein endothelial cells (HUVECs) were purchased from ATCC (Rockville, MD) and cultured in M199 supplemented with 20% fetal bovine serum. We established short-term cultures of primary gastric carcinoma cells T051 and normal gastric cells N052 using the tissues obtained from patient with stomach cancer undergoing surgery. These surgically removed human gastric tissues were collected by our collaborators at the Huadong Hospital. All human tissue samples were obtained and handled in accordance with the approved Institutional Review Board application. The primary cells were cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum, 10 μM insulin, and 1 μM hydrocortisone.
Virus construction
The IL-24 expression cassette was released from pZD55-IL-24 (Zhao et al., 2005) and introduced into pMUD55 (Wei et al., 2009) to produce pMUD55-IL-24. Adenovirus MUD55-IL-24 was generated by homologous recombination using pBHGE3 and pMUD55-IL-24 with Effectene Transfection Reagent (Qiagen, Hilden, Germany). The construction of the replication-defective virus Ad-IL-24 and the oncolytic adenovirus MUD55 were reported previously (Zhao et al., 2005; Wei et al., 2009).
Western blot assay
Cells were seeded in 6-cm dishes and treated with phosphate-buffered saline (PBS) or different agents. For determination of the secreted IL-24, conditional media (CM) were collected at the indicated times. Cells were harvested from the plates and lysed in lysis buffer (50 mM Tris-HCl [pH 8.0], 150 mM NaCl, 0.02% sodium azide, 100 μg/ml PMSF (phenylmethanesulfonyl fluoride), 1 μg/ml aprotinin, and 1% Triton X-100). Total protein (40 μg) was electrophoretically separated on 8%–12% SDS-PAGE and transferred to a PVDF (polyvinylidene fluoride) membrane. Primary antibodies for E1A (Santa Cruz, Santa Cruz, CA), E1B-55K (Oncogene, Cambridge, MA), IL-24 (GenHunter, Nashville, TN), β-actin (Santa Cruz), caspase-9 (Santa Cruz), caspase-3 (Santa Cruz), and PARP (Santa Cruz) were used to detect the expression of E1A, E1B-55K, IL-24, β-actin, and the cleavage of caspase-9, caspase-3, and PARP [Poly(ADP-ribose)polymerase], respectively. After incubation with the secondary antibodies (all purchased from Santa Cruz), the membranes were revealed with the ECL (enhanced luminol-based chemiluminescence) detection reagents (Pierce Biotechnology, Rockford, IL).
Cytopathic effect assay
For the crystal violet staining, cells were seeded in 24-well plates, treated with PBS or different viruses at the indicated multiplicity of infections (MOIs), and stained with 2% crystal violet solution at 5 days after infection. For the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay, cells were treated with PBS or different viruses at an MOI of 5. After the indicated times, MTT solution was added, and the absorbance was read at 595 and 630 nm with a DNA microplate reader (Bio-Rad, Hercules, CA). To detect the cytopathic effect (CPE) of MUD55-IL-24 in primary cells, the morphology of cells treated with PBS or MUD55-IL-24 was monitored with phase-contrast microscope and documented by photographs at 72 hr posttreatment.
Viral replication assay
To determine virus progeny, different cells were infected at a MOI of 5. After 2 days, viruses were released by three freeze–thawing cycles and determined by plaque assay in HEK293 cells.
In vivo animal experiments
All the male BALB/c nude mice obtained from Animal Research Committee of the Institute of Biochemistry and Cell Biology (Shanghai, China) were 3–4 weeks old and maintained in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals. SG-7901 cells (3 × 106) were injected subcutaneously into the right flank of the nude mice. When tumors reached sizes of 100–150 mm3, mice were randomly divided into four groups (six mice/group) and treated with five consecutive daily intratumoral injections of PBS or diluted viruses (4 × 108 pfu/day). Tumor sizes were measured twice per week. The volume was calculated according to the following equation: V (mm3) = width2 × length/2. Tumors were removed, weighed, and placed in formalin when the mice were dead or euthanized. Besides, the livers were harvested and embedded in paraffin for the following hepatotoxicity analysis.
Histopathology and immunohistochemistry analysis
For histopathology analysis, the paraffin sections of livers or tumors were stained with hematoxylin and eosin (H&E). For immunohistochemistry analysis, sections were treated with anti-hexon (Chemicon, Temecular, CA) or anti-IL-24 (GenHunter) antibody. Goat or mouse ABC staining systems (Santa Cruz) were used in staining steps. Terminal deoxynucleotidyltransferase-mediated dUTP-biotin nick end-labeling (TUNEL) method was performed using the In Situ Apoptosis Detection Kit (R&D Systems, Minneapolis, MN).
Measurement of apoptosis in vitro
For the flow cytometry analysis, cells after treatment were trypsinized, washed once with PBS, and stained with fluorescein isothiocyanate-labeled annexin V and propidium iodide (BioVision, Mountain View, CA). Fluorescence was analyzed using a FACSort flow cytometer (Becton Dickinson, Franklin Lakes, NJ). The N-acetyl-
Assessment of ROS generation
To determine the intracellular ROS generation, cells were trypsinized, washed with PBS, stained with dichlorohydrofluorescein diacetate (10 μM; Beyotime) for 30 min at 37°C, and analyzed by flow cytometry immediately after staining.
Cytofluorimetric analysis of mitochondrial membrane potential (ΔΨ m )
The variation of mitochondrial transmembrane potential ΔΨm was measured using 5,5′,6,6′-tetrachloro-1,1′,3,3′-tetraethylbenzimidazolcarbocyanine iodide (JC-1; Beyotime). Cells were harvested, incubated in culture medium with JC-1 for 20 min at 37°C in a humidified atmosphere, washed twice, and analyzed by flow cytometry.
Tube formation assay
SG-7901 cells were infected with viruses at an MOI of 5. Three days postinfection, the CM were heated at 56°C to inactivate the viruses. A 96-well plate coated with Matrigel (Becton Dickinson) was incubated at 37°C for 30 min. HUVECs were seeded onto its surface with the CM above. After 30 min, 10 ng/ml rhVEGF (recombinant human vascular endothelial growth factor) was added to the medium. The tube formation was photographed after 12 hr and quantified.
Scratch wound assay
SG-7901 cells were grown to confluence in a 24-well plate. A single scratch wound was created in each well using the Essen Woundmarker (Ann Arbor, MI). The CM collected in the tube formation assay were added to the wells and the plate was then inserted into the Incucyte instrument (Ann Arbor, MI). The wound area in each well was imaged every 3 hr.
Statistical analysis
The statistical significances were assessed with Student's t test. p < 0.05 was considered statistically significant.
Results
Construction of oncolytic adenovirus MUD55-IL-24 and its selective IL-24 expression
Previously we constructed the adenocarcinoma-selective oncolytic adenovirus MUD55, in which E1A was under the control of MUC1 promoter and E1B-55K gene was deleted (Wei et al., 2009). In this study, therapeutic gene IL-24 was inserted into MUD55 and a gene-virotherapy agent MUD55-IL-24 was generated. The replication-deficient virus Ad-IL-24 with a deletion of E1 region was used as control (Fig. 1A). To identify these viruses, SG-7901 cells were treated with PBS, Ad-IL-24, MUD55, and MUD55-IL-24, respectively, and the expression of E1A and E1B-55K were examined. E1A expressed by MUD55 and MUD55-IL-24 was similar to the positive control, whereas all the viruses did not express E1B-55K (Fig. 1B).
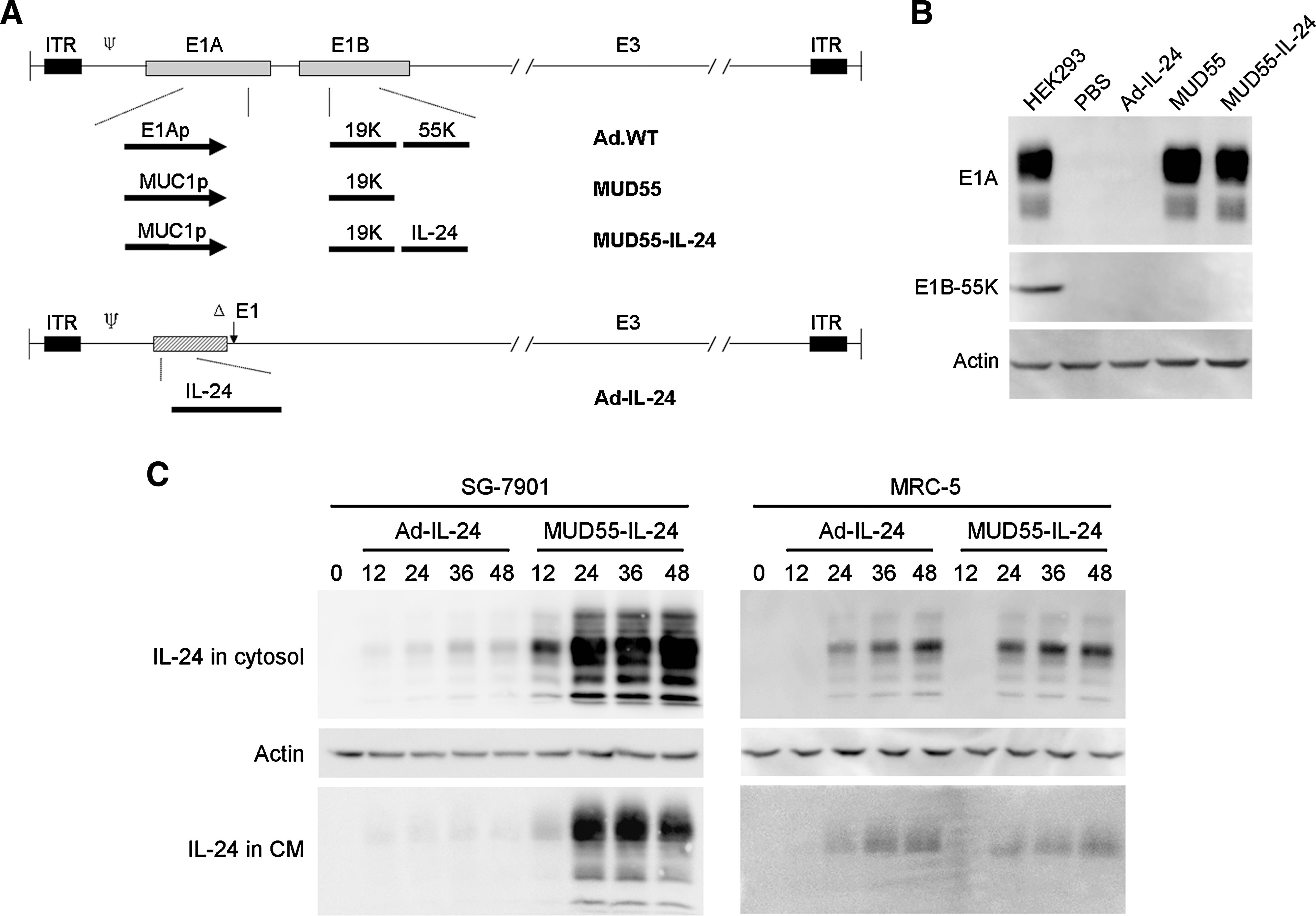
Characterization of adenoviruses. (
The replication-competent adenovirus MUD55-IL-24 was expected to express IL-24 more efficiently than the replication-deficient adenovirus Ad-IL-24. This was confirmed by the Western blot assay. The kinetics of MUD55-IL-24- and Ad-IL-24-mediated IL-24 expression in the cytosol and IL-24 secretion in the CM were both detected. MUD55-IL-24 expressed IL-24 comparable to Ad-IL-24 in the normal cell line MRC-5, but much more efficiently than Ad-IL-24 in the gastric carcinoma cell line SG-7901 (Fig. 1C).
Gastric tumor-specific CPE of MUD55-IL-24 in vitro and in primary culture systems
The CPE of MUD55-IL-24 was first evaluated via crystal violet staining. In SG-7901 cells, the cytotoxicity caused by MUD55-IL-24 was about 100-fold greater than that of MUD55 or Ad-IL-24. In contrast, in MRC-5 cells, MUD55-IL-24 gave no obvious cytotoxicity (Fig. 2A). Similar results were obtained in the MTT assay. The difference in cell viability of the SG-7901 cells treated with Ad-IL-24, MUD55, and MUD55-IL-24, respectively, became apparent at 48 hr after infection. Only 20% of MUD55-IL-24-infected cells were alive, whereas over 70% of MUD55- or Ad-IL-24-infected cells remained viable at 96 hr after treatment (Fig. 2B).
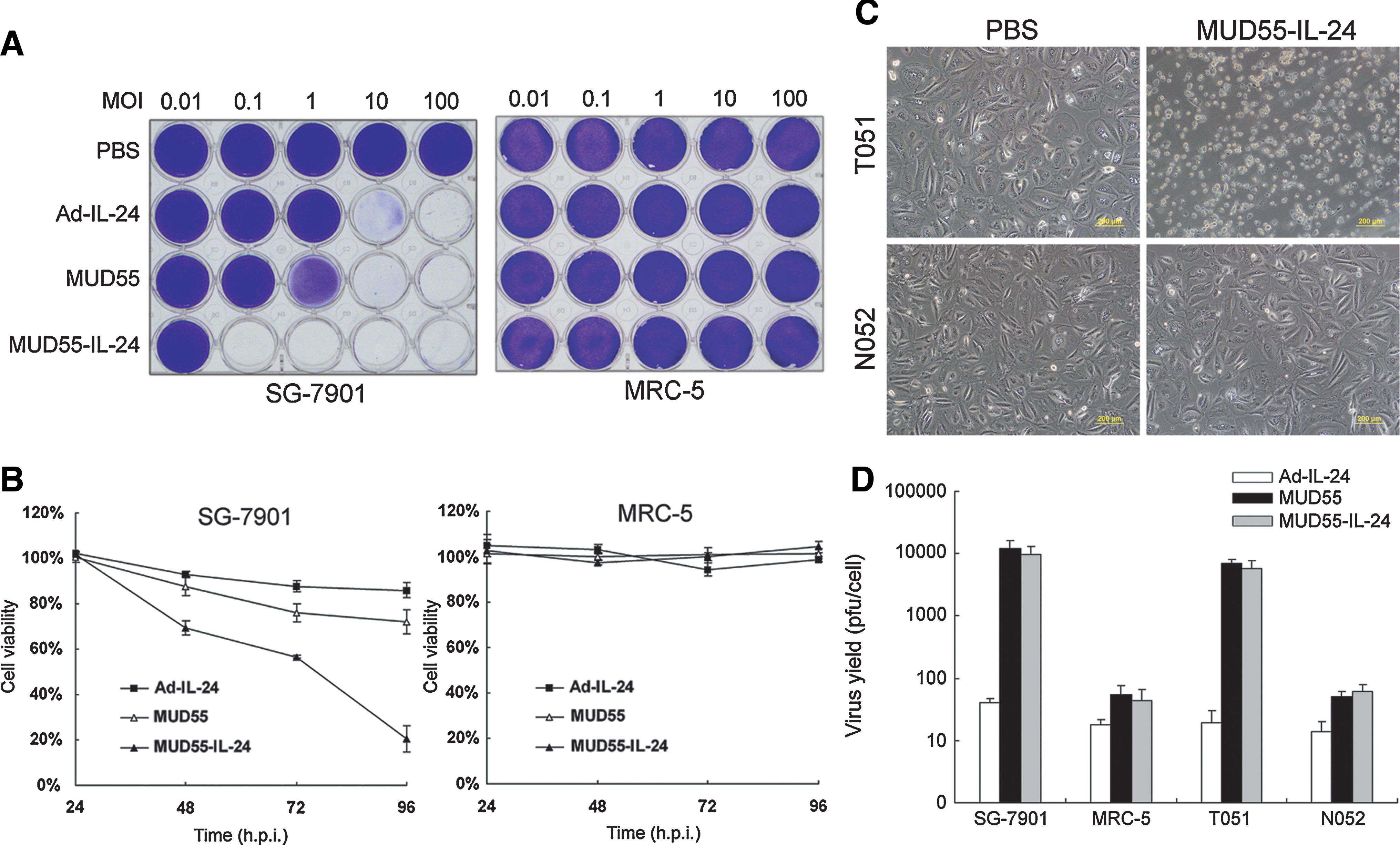
Gastric tumor-specific cytopathic effect of MUD55-IL-24 in vitro and in primary culture systems. (
To further confirm the CPE of MUD55-IL-24 in a broader panel of gastric tissues, we established short-term cultures of primary gastric carcinoma cells T051 and normal gastric cells N052. The morphology of PBS- or MUD55-IL-24-treated cells was recorded at 72 hr after treatment. There was evident CPE in MUD55-IL-24-treated T051 cells, whereas no obvious pathological changes were found in N052 cells (Fig. 2C).
The replication selectivity of viruses was then evaluated by the progeny assay. As shown in Fig. 2D, the progeny production of MUD55-IL-24 was about 1000-fold larger than that of Ad-IL-24 in SG-7901 and T051 cells, whereas only 10-fold more than that of Ad-IL-24 in MRC-5 and N052 cells. There was no obvious difference in viral production between MUD55 and MUD55-IL-24, suggesting that the introduction of IL-24 did not affect viral replication ability.
Collectively, both the in vitro and primary culture experiments showed the highly selective antigastric tumor activity of MUD55-IL-24.
Antitumor effect of MUD55-IL-24 in nude mice
After an SG-7901 human gastric adenocarcinoma xenograft model was established in nude mice, the constructed agents were intratumorally administrated. MUD55-IL-24 exhibited much more significant tumoricidal activity compared with Ad-IL-24 and MUD55, consistent with the results in vitro. About 90% of tumor growth was inhibited by MUD55-IL-24 at the end of this study (Fig. 3A). No animal death was observed in the MUD55-IL-24 treatment group throughout the experiment. In contrast, all the mice treated with PBS were moribund at 4 weeks after treatment and two of them died on days 31 and 33 posttreatment, respectively. When the average tumor volume exceeded 1500 mm3 at 6-week posttreatment, we had to euthanize the mice in the PBS group. In the Ad-IL-24-treated group, three mice died on days 41, 46, and 53, respectively. In the MUD55-treated group, two mice died on days 43 and 54, respectively. Besides, the MUD55-IL-24-treated tumors displayed the least net tumor weight at the end of this experiment (Fig. 3B). In addition, H&E staining of the liver sections from MUD55-IL-24-treated mice showed no obvious hepatotoxicity signs (data not shown).
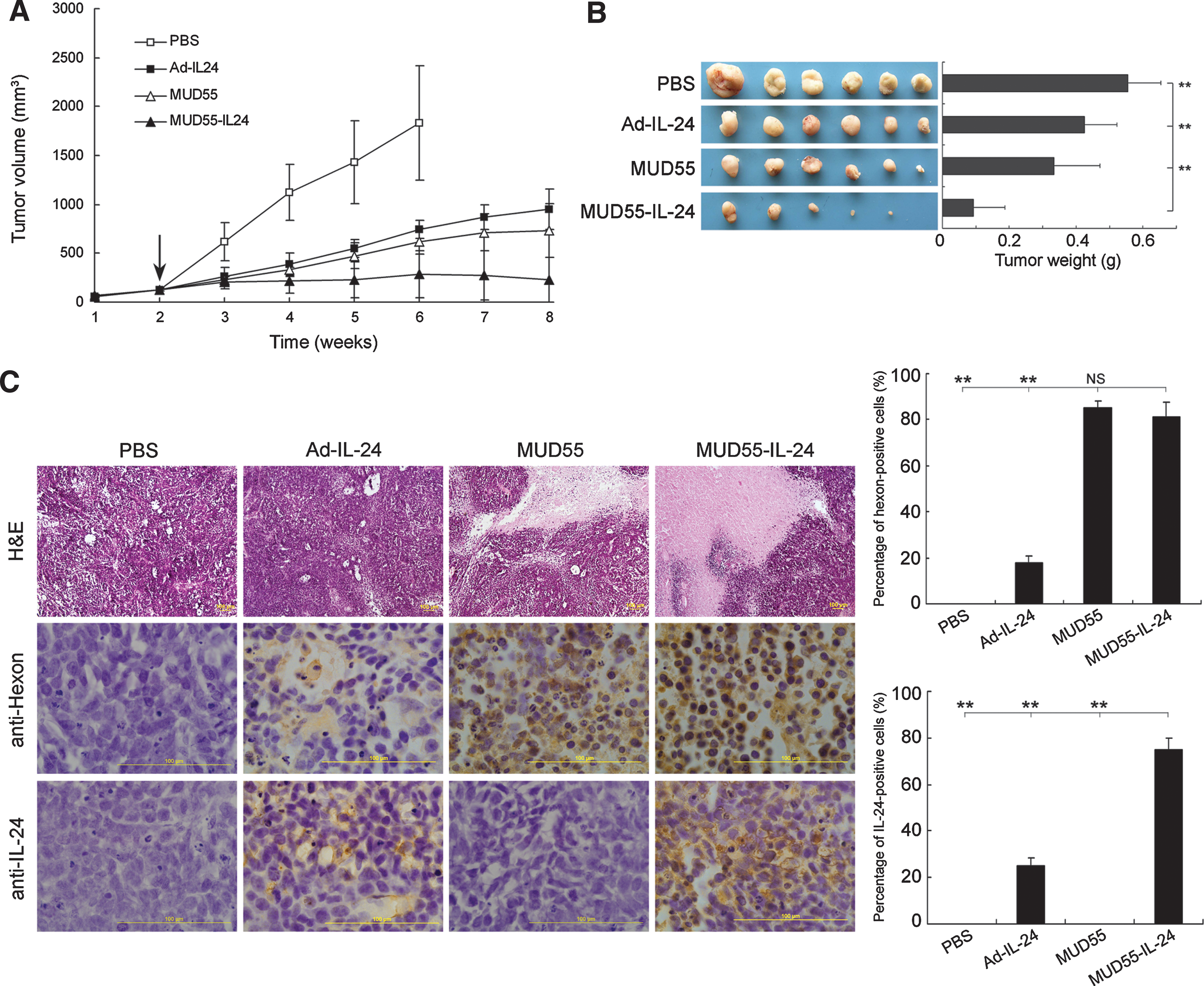
Antitumor activity of MUD55-IL-24 in SG-7901 xenograft model. BALB/c nude mice bearing SG-7901 xenografts were treated with PBS or different viruses by intratumoral administration. (
The tumors were subjected to H&E staining for pathologic examination. Necrotic areas in the MUD55-IL-24-treated tumors were larger than that in the PBS-, Ad-IL-24-, or MUD55-treated tumors. There were over 70% regions positive for hexon and IL-24 staining in the MUD55-IL-24-treated tumors, detected by Ad5 hexon and IL-24 immunohistochemical staining. These results confirmed the oncolytic ability of MUD55-IL-24 in the treatment of gastric adenocarcinoma in vivo. The promising outcome most likely came from the replication of the adenovirus together with the expression of the therapeutic gene (Fig. 3C).
MUD55-IL-24 induced profound apoptosis in gastric carcinoma
IL-24 is known to induce apoptosis in diverse cancers. To look into the mechanisms underlying the significant antitumor activity of MUD55-IL-24, we first evaluated its apoptosis induction effect in tumor sections by TUNEL staining. The percentage of TUNEL-positive cells in MUD55-IL-24-treated tumors was about 90%, whereas this proportion in Ad-IL-24- and MUD55-treated tumors was much less (about 15% and 35%, respectively) (Fig. 4A).
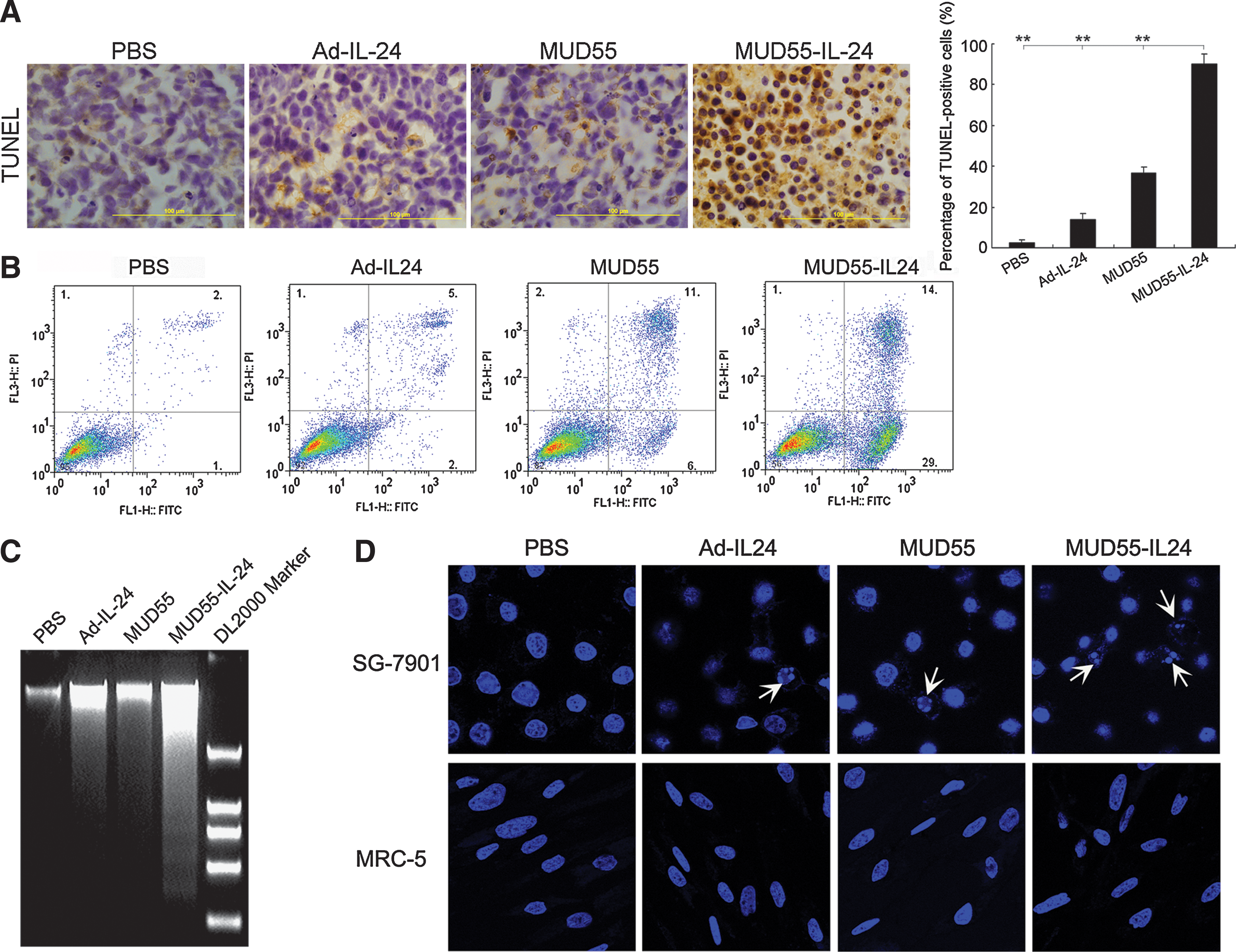
MUD55-IL-24 selectively induced apoptosis in gastric carcinoma both in vitro and in vivo. (
The apoptosis induction capacity of MUD55-IL-24 in vitro was measured with several methods. In the annexin V and propidium iodide staining assay, we found that 43% of MUD55-IL-24-infected SG-7901 cells were annexin V positive, whereas Ad-IL-24- and MUD55-infected cells manifested much less apoptosis (7% and 17%, respectively) (Fig. 4B). Similar findings were observed when apoptosis was evaluated by nucleosomal DNA degradation. MUD55-IL-24 induced DNA ladder more efficiently than Ad-IL-24 and MUD55 in SG-7901 cells (Fig. 4C). Further, Hoechst 33258 staining was used to detect the apoptotic changes in different cell lines. In SG-7901 cells, MUD55-IL-24 induced much more significant nuclear fragmentation and chromatin clumping than Ad-IL-24 and MUD55, whereas in MRC-5 cells, no obvious changes were detected (Fig. 4D). Taken together, all these results indicate that the antitumor activity of MUD55-IL-24 in gastric cancer may have come from its apoptosis induction ability.
MUD55-IL-24 induced ROS generation, mitochondria dysfunction, and caspases activation
ROS (including singlet oxygen, superoxide ions, hydroxide, and hydroxyl radicals) are known to regulate various cellular responses including apoptosis. To clarify the possible pathways by which MUD55-IL-24 induced apoptosis, we evaluated the cellular ROS production using the probe dichlorohydrofluorescein diacetate. Figure 5A shows a significant increase in ROS generation in MUD55-IL-24-treated SG-7901 cells compared with untreated cells, suggesting the involvement of ROS in apoptosis induction. The observation that the percentage of annexin V-positive SG-7901 cells infected with MUD55-IL-24 was greatly reduced when pretreated with antioxidants such as NAC further supported our suggestion (Fig. 5B).
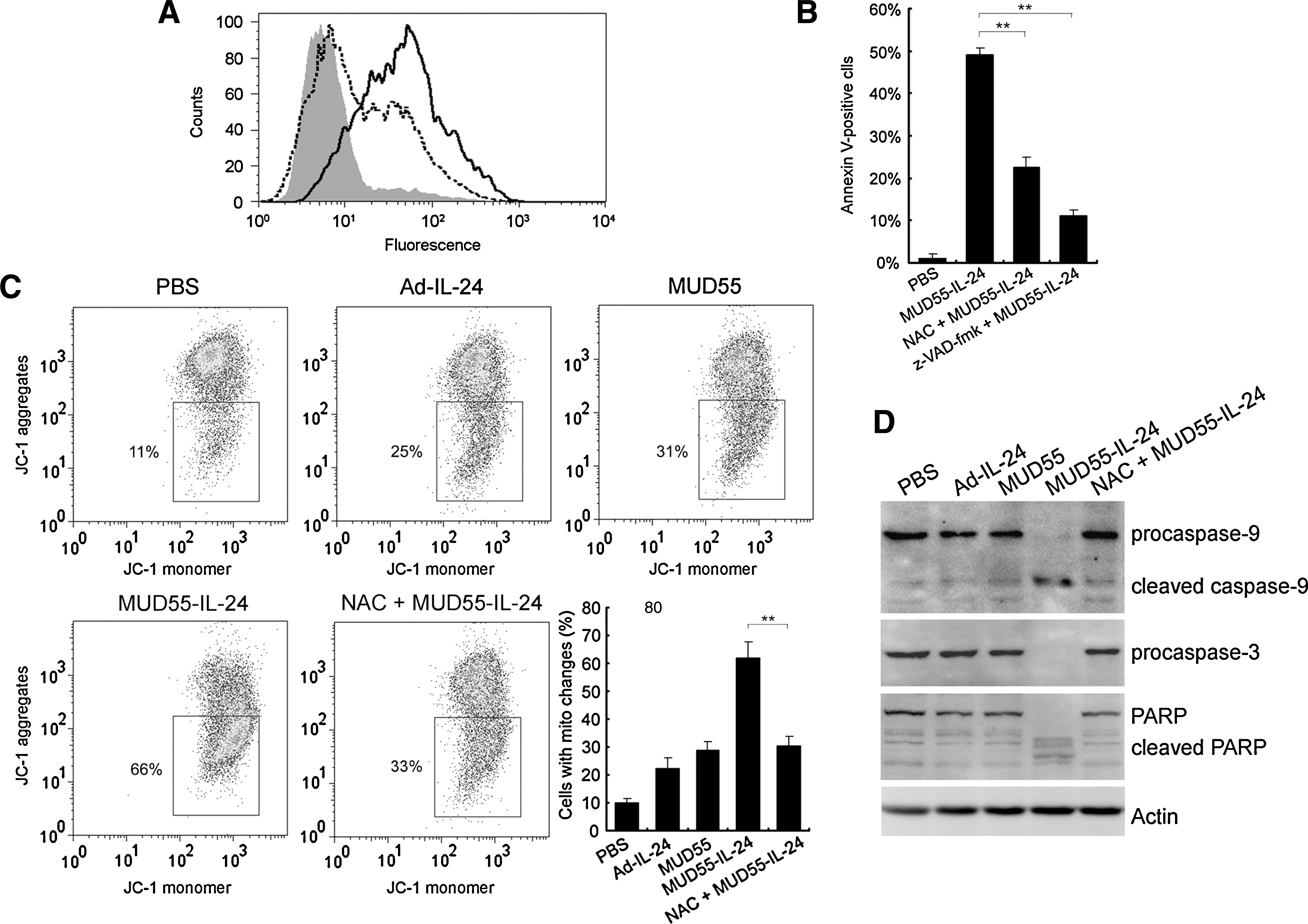
MUD55-IL-24 induced ROS generation, mitochondria dysfunction, and caspase activation in SG-7901 cells. (
We measured the mitochondrial membrane potential (ΔΨm) in SG-7901 cells by JC-1 probe. A marked increase in JC-1 monomers, indicating a drop in ΔΨm, was observed in over 60% of MUD55-IL-24-infected cells, whereas similar phenomena were present in only about 30% of Ad-IL-24- and MUD55-treated cells. Besides, the decrease of ΔΨm in MUD55-IL-24-infected cells was rescued by antioxidants such as NAC, indicating that ROS generation is upstream of mitochondrial dysfunction in the apoptotic pathway (Fig. 5C).
Mitochondrial dysfunction is always followed by the activation of caspase-9, which in turn activates caspase-3 and cleaves PARP. This was confirmed by the Western blot assay (Fig. 5D). Actin was used as control. The apoptosis was significantly blocked by z-VAD-fmk, a pan-caspase inhibitor, suggesting that the MUD55-IL-24-induced apoptosis was caspase dependent (Fig. 5B).
Antiangiogenesis and antimigration effect of MUD55-IL-24
As it has been reported that secreted IL-24 is a potent inhibitor of angiogenesis, we further examined the antiangiogenesis activity of MUD55-IL-24 by the tube formation assay. The ability of HUVECs to form tube-like structures was more inhibited by CM from SG-7901 cells, which were infected by MUD55-IL-24 rather than by Ad-IL-24. The CM from MUD55- or PBS-treated SG-7901 cells did not inhibit the tube formation obviously (Fig. 6A).
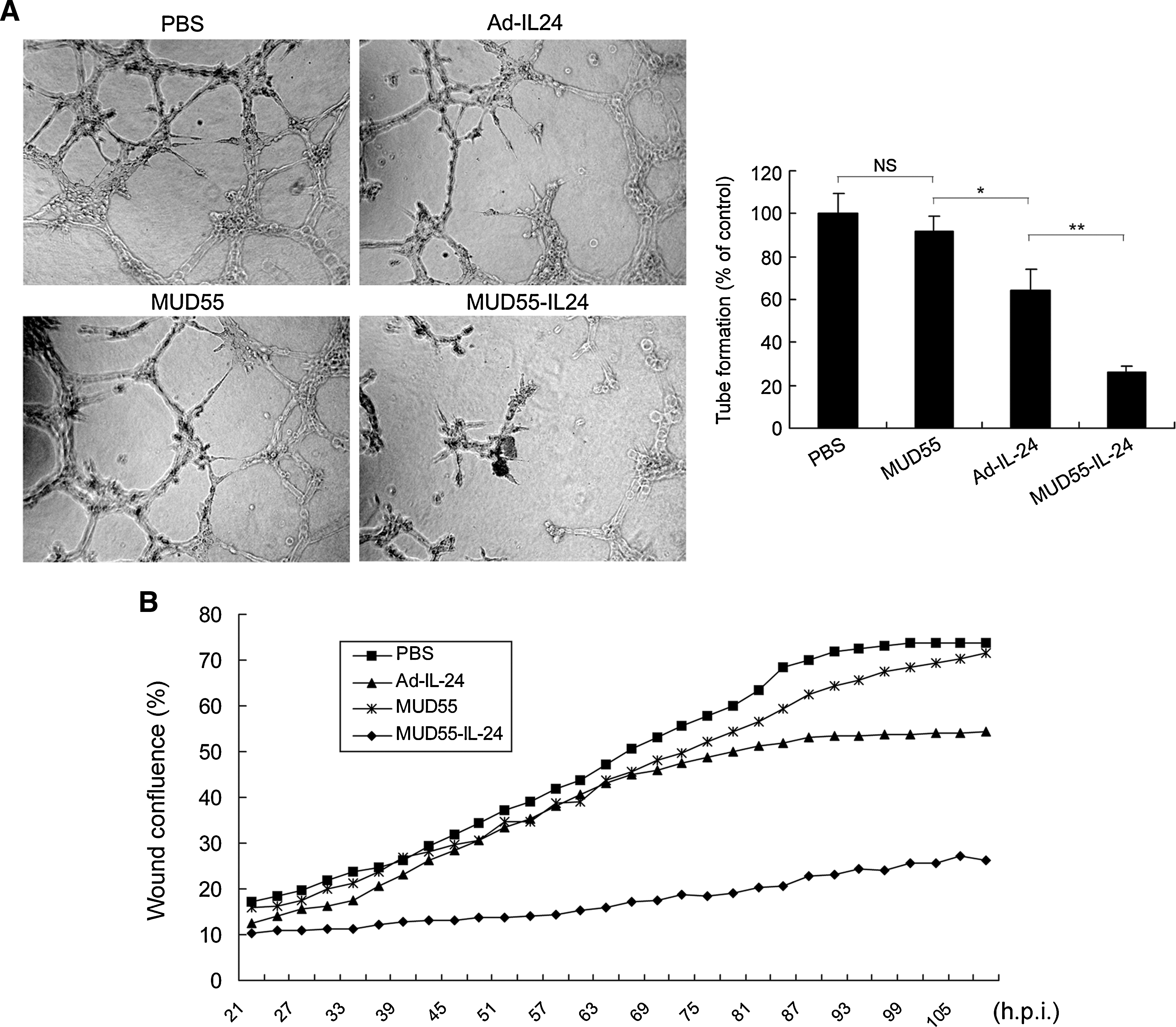
Antiangiogenesis and antimigration activities of MUD55-IL-24 in vitro. (
We also evaluated the antimigration ability of different viruses by the wound healing assay. The SG-7901 cells treated by the CM from MUD55-IL-24-infected cells migrated much slower than the cells treated by the CM from Ad-IL-24- or MUD55-infected cells (Fig. 6B).
Discussion
Because IL-24 selectively induces cell death in tumor but not in normal cells, using a replication-deficient adenovirus as vector, it has shown great therapeutic potential (Saeki et al., 2000; Fisher et al., 2003; Mahasreshti et al., 2006; Wang et al., 2006; Wei et al., 2008). In the present study, we further improved the antitumor activity of IL-24 by utilizing a replication-competent oncolytic adenovirus MUD55 as the vector and generated a gene-virotherapy agent MUD55-IL-24. It showed stomach cancer-specific adenoviral replication, IL-24 expression, cytopathic effect, and apoptosis induction not only in vitro, but also in primary culture systems. Injecting MUD55-IL-24 into gastric carcinoma xenografts in nude mice inhibited about 90% tumor growth without obvious liver toxicity. Based on these observations, we are optimistic that MUD55-IL-24 may be a safe and potent candidate for the future treatment of gastric carcinoma.
Cancer can be caused by multiple genomic modifications in transformed cells. As different varieties of cancer have different inducements, targeting therapies, such as gene therapy and virotherapy, are developing quickly nowadays. They can act specifically on a certain tumor and minimize harmful effects to normal tissues. Our approach is to combine the virtues of gene therapy and virotherapy together and employ a novel strategy named “targeting gene-virotherapy” to treat cancer (Zhao et al., 2005; Liu and Gu, 2006). Using an oncolytic adenovirus as vector, the expression of therapeutic gene can be significantly enhanced accompanying with the viral replication in tumor cells. It has been proved in this study that the gene-virotherapy agent MUD55-IL-24 expressed IL-24 much more efficiently than the traditional gene therapy agent Ad-IL-24 in gastric tumor cells. It showed better antitumor effect than both Ad-IL-24 and MUD55 in vitro and in vivo. Likewise, Devanand et al. have also observed potent oncolytic effect of the replicable adenovirus Ad.PEG-E1A-mda-7 on breast carcinoma (Sarkar et al., 2005), prostate cancer (Sarkar et al., 2007b), and melanomas (Sarkar et al., 2008), and Zhao et al. (2005) reported efficient antitumor effect of ZD55-IL-24 on colorectal cancer. Compared with those adenoviruses, the safety of MUD55-IL-24 was greatly improved because the double-regulated viral vector MUD55 was confirmed to be safer than the monoregulated adenoviruses and showed limited liver toxicity when administrated by intravenous injection in nude mice (Wei et al., 2009).
Multiple laboratories previously reported the apoptosis induction (Sarkar et al., 2002; Sauane et al., 2003, 2004; Wang et al., 2007), antimigration (Dent et al., 2005), and antiangiogenesis (Lebedeva et al., 2007; Sarkar et al., 2007a) activities of IL-24 in a variety of tumor types, but the underlying mechanism was unclear. In this study, we observed that MUD55-IL-24 also possessed these relevant properties and seemed to be stronger than Ad-IL-24 and MUD55 in gastric carcinoma cells. MUD55-IL-24 induced apoptosis in stomach cancer cells probably through promoting ROS generation and mitochondrial dysfunction. Antioxidants such as NAC inhibited the killing effect of MUD55-IL-24. This was consistent with the results that Lebedeva et al. (2003) obtained in Ad.mda-7-infected prostate cancer cells. It has also been reported by Su et al. that Ad.mda-7 altered the ratio of pro- (such as Bcl-2, Bcl-xL, and McL-1) and antiapoptotic (such as Bax, Bad, and Bak) proteins, causing the decline in ΔΨm and tipping the balance from survival to death in breast cancer cells. However, we failed to observe these ratio changes in MUD55-IL-24-infected SG-7901 cells (data not shown). ROS generation may contribute a lot in the mitochondrial dysfunction process because NAC rescued partial cells from decreasing in ΔΨm. Moreover, we observed evident cleavage of caspase-9, caspase-3, and PARP, and the apoptosis was greatly inhibited by pan-caspase inhibitor z-VAD-fmk, indicating that the cell death induced by MUD55-IL-24 in SG-7901 cells is a caspase-dependent apoptosis process.
Footnotes
Acknowledgments
The authors thank Lanying Sun for help with cell culture. This work was supported by the National Nature Science Foundation of China (no. 30623003), the Science and Technology Commission of Shanghai Municipality (no. 06DZ22032), the National Basic Research Program of China (973 Program; no. 2004CB51804), the Hi-Tech Research Development Program of China (863 Program; no. 2007AA021006), the Key Project of the Chinese Academy of Science (no. KSCX2-YW-R-09, R-04), the National Basic Research Committee of Science and Technology (no. 06ZR14072 and 074119508), and the Zhejiang Sci-Tech University (no. 0616033).
Author Disclosure Statement
No competing financial interests exist.