
Abstract
Advances in delivery techniques and in expression construct design have renewed interest in nonviral gene transfer. Here, we test plasmid or bacterial backbone free minicircle vectors for factor IX (FIX) expression by hydrodynamic liver-directed delivery. Both constructs are driven by a hepatic control region, the human α1-antitrypsin promoter, which results in long-term expression in FIX knockout mice. However, levels of expression were higher and expression loss over time was reduced when using minicircles. Even at the highest expression levels (>700% of normal) FIX was fully functional. Transgene loss was the main determinant for expression loss over time for both vector types. A significant effect of gene silencing was observed only for the plasmid, not for the minicircle vector. To determine the influence of promoter methylation, we performed bisulfite-mediated conversion and sequencing of vector DNA on days 14 and 100 after gene transfer. We determined a higher frequency of methyl-protected cytosines in CpGs and a lower degree of demethylation at bacterial Dcm methylation sequences near transcription factor-binding sites in the α1-antitrypsin promoter in plasmid compared with minicircle mice on day 100. Therefore, the methylation status might reflect differences in the levels and durability of expression. Judging from the high levels of functional FIX obtained, small fractions of liver or single liver segments should be sufficient to reach therapeutic efficacy in translating hydrodynamic delivery to humans. However, transgene loss remains to be addressed to further guarantee sustained expression over time.
Introduction
In the present study, we investigated nonviral gene transfer for coagulation factor IX (FIX) to the liver. FIX is the deficient coagulation protease in the X-chromosomal inherited bleeding disorder hemophilia B. Gene delivery to the liver of affected individuals has already proven to be clinically effective using an adeno-associated viral vector (Manno et al., 2006). However, long-term transgene expression has not been obtained in humans to date. Although high levels of expression were obtained in mice by nonviral gene transfer (Miao et al., 2000; Chen et al., 2003), expression levels significantly decline over time. Furthermore, by targeting liver segments in humans it is still unknown whether a small fraction of liver cells can produce sustained levels of functional protein (Fabre et al., 2008) by catheter-based transfer approaches.
We tested a plasmid bacterial backbone free minicircle (MC) vector using a highly efficient liver-specific expression cassette and compared the results with regular plasmid vector (pV) constructs. DNA copy measurements revealed vector genome loss as the main determinant for the decline in expression. At later time points, we detected differences in the methylation status of the vector promoter region between pV- and MC-treated mice. Therefore, these changes could represent an additional component responsible for vector silencing.
Materials and Methods
Nonviral vector constructs
Two different expression vectors for FIX were employed for the experiments (see Fig. 1A and B). The expression cassette was introduced either into a plasmid vector containing the pSL1180 backbone or into an MC producer plasmid kindly provided by M. Kay (Stanford University School of Medicine, Stanford, CA) (Chen et al., 2005). This system allows the elimination of bacterial plasmid backbone sequence for circular DNA vectors. MCs were then purified by passage through a regular one-step affinity column.

(
Mouse models
All animal procedures were approved by the local animal care, protection, and use authorities. C57BL/6 mice were purchased from Harlan (Indianapolis, IN). FIX-deficient mice on a C57BL/6 background were kindly provided by K. High (Children's Hospital of Philadelphia, Philadelphia, PA) (Schuettrumpf et al., 2005). The nonviral gene transfer vector was administered by hydrodynamic injection technique as previously described (Schüttrumpf et al., 2008). Blood was collected either by retro-orbital bleeding, or by bleeding following tail cut.
FIX assays
FIX concentrations were determined in an enzyme-linked immunosorbent assay in which a monoclonal antibody to human FIX, clone HIX-1 (Sigma-Aldrich, St. Louis, MO) was used as a capture antibody. Peroxidase-conjugated polyclonal goat anti-human FIX (Affinity Biologicals, Hamilton, ON, Canada) was used as the detecting antibody. FIX functional activity was determined by a modified one-stage factor assay; 25 μl of human FIX-deficient plasma was incubated with 25 μl of automated activated partial thromboplastin time (aPTT) reagent (Dade Behring, Marburg, Germany). A total of 25 μl from each test sample (when required, samples were diluted in imidazole buffer for 3 min at 37°C) was added to 25 μl of CaCl2 (25 mM); time to clot formation was measured with a fibrometer. Antibodies to FIX were measured by ELISA (specific to murine immunoglobulin subclasses) as described (Schuettrumpf et al., 2005) with minor modifications.
Vector DNA analysis
Treated mice were killed at various time points after vector treatment. Representative liver samples from all major liver lobes were used for the following DNA extraction. Whole genomic DNA was prepared with a DNeasy blood & tissue kit (Qiagen, Hilden, Germany). Determination of gene copy number was performed by quantitative real-time polymerase chain reaction (PCR), using a QuantiTect probe PCR kit (Qiagen) and a PRISM 7700 sequence detection system (Applied Biosystems, Foster City, CA). For this purpose 100 ng of DNA was applied as template in a PCR using the following primers and probe for vector detection: 5′-TTCGATCTACAAAGTTCACCATCTATAAC-3′ (forward), 5′-AAACTGGTCCCTTCCACTTCA G-3′ (reverse), and FAM-AATCTCTACCTCCTTCATGGAAGCCAGCA-BHQ-1 (probe). Genomic β-actin gene measurements with the following primers were used as internal controls: 5′-AGAGGGAAATCGTGCGTGAC-3′ (forward), 5′-CAATAGTGACCTGGCCGT-3′ (reverse), and TET-CACTGCCGCATCCTCTTCCTCCC-BHQ-1 (probe).
For methylation status analysis of the vector promoter region we performed a bisulfite-mediated conversion of unmethylated cytosines, using an EpiTect bisulfite kit (Qiagen) with 200 ng of liver DNA. The converted DNA was used as template for a nested PCR to amplify a CpG-enriched 500-bp fragment containing the human α1-antitrypsin (hAAT) promoter and the 5′ untranslated region (UTR) of the FIX expression cassette (see Fig. 3B) in both plasmid and minicircle vectors, using the primers 5′-ATTTTTTTTATTTTGGATATAGGA-3′ and 5′-TAAAAAACAAATAATAATAAAACCT-3′ for the second PCR run. The corresponding PCR fragments were subcloned into the pCR2.1-TOPO vector (Invitrogen, Carlsbad, CA) and sequenced with the M13 forward and M13 reverse primers.
Statistical analysis
The Student t test was employed for parametric data and the Fisher exact test for categorical data including the Bonferroni adjustment for multiple comparisons, using GraphPad InStat software 3.06 (GraphPad Software, La Jolla, CA).
Results and Discussion
MC gene transfer results in higher and more sustained FIX expression levels
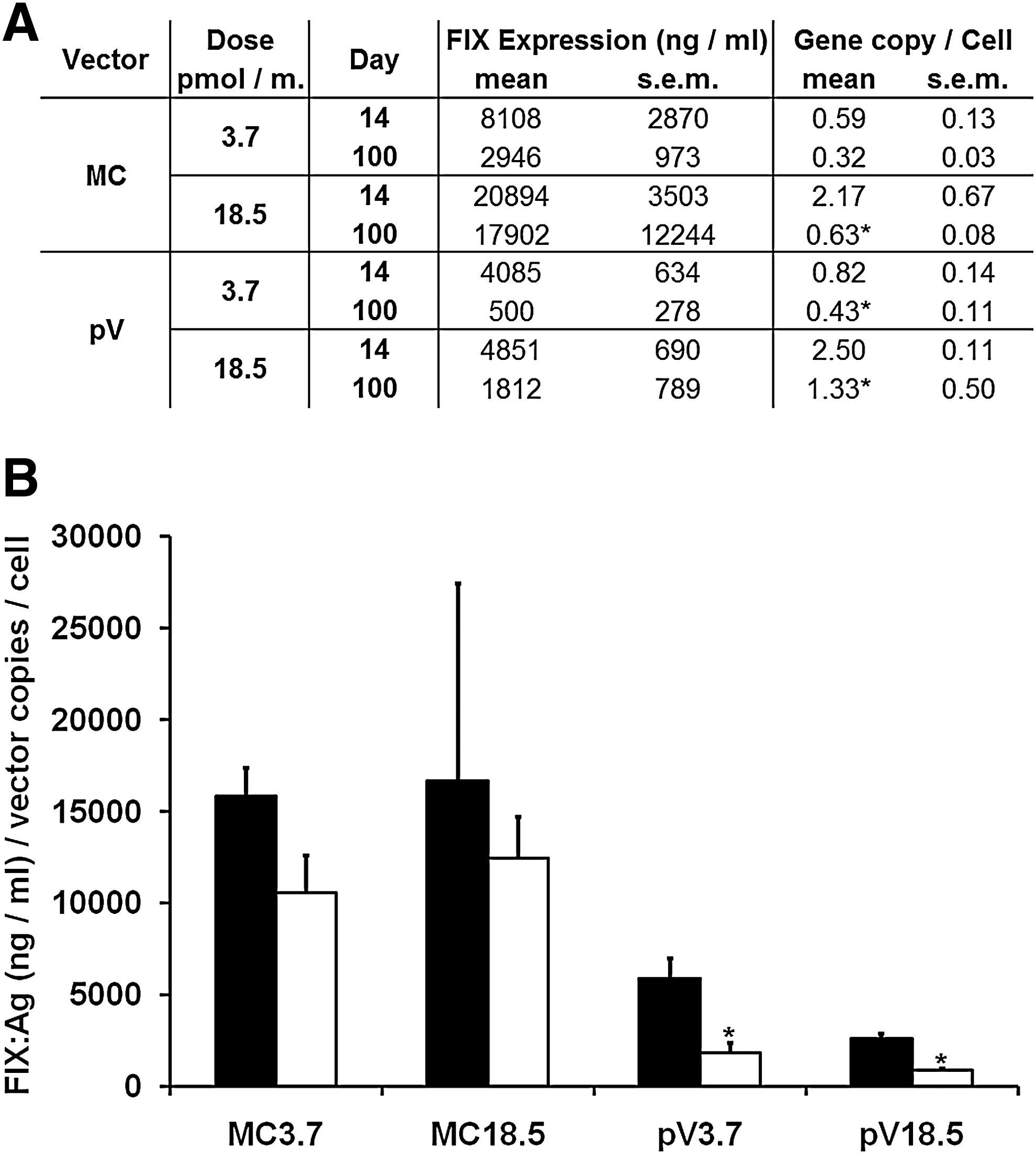
FIX knockout mice were treated with either pV or MC at a dose of 3.7 or 18.5 pmol of vector per mouse, corresponding to 17.6 and 88 μg of pV (size, 7.3 kb), or 10 and 50 μg of MC (size, 4.2 kb), respectively. Figure 1C shows the expression levels in mice treated with the lower vector doses and Fig. 1D shows the levels for the higher vector doses. Vector injection resulted in a fast increase in expression. The highest expression levels were obtained on day 14 in most of the mice. Initially, expression levels using MC were about twice as high compared with pV. There was a continuous decline in FIX levels until the end of the experiment on day 100 for all groups of mice, resulting in 4- to 10-fold higher levels for MC-treated mice. A comparison of expression levels between day 100 and day 14 for pV-treated mice is represented as a ratio in Fig. 1E for MC- and pV-treated mice. Overall, the decline in expression was less pronounced in the MC-treated mice (0.6 compared with 0.25; p < 0.05).
This observation is in accordance with previously published data, in which expression plasmids with an HCR1/hAAT-FIX expression cassette (Miao et al., 2000; Schüttrumpf et al., 2008) or bacterial backbone free minicircle constructs (Chen et al., 2003) were able to ensure sustained FIX expression after hydrodynamic injection into mice. Chen and colleagues (2003) reported a 45-fold higher change in FIX expression levels when using MC vectors and even 560-fold higher levels in a similar experiment using α1-antitrypsin as the transgene. On day 100 expression levels between MC- or pV-treated differed 4- to 10-fold. The difference might be attributed to the relatively small degree of decline observed in the pV group and could represent differences in the bacterial backbone or in the efficiency of the expression cassette. Furthermore, the advantage of MC vectors resulting in stable transgene expression increased over time, so that differences were expected to increase with the duration of the experiments.
Supraphysiological expression of fully functional FIX after MC gene transfer
Despite the promising results in mice, the translation of similar hydrodynamic delivery techniques to humans remains a major obstacle. Several reports from large animal models (Fabre et al., 2008) have suggested the feasibility of catheter-mediated naked DNA vector transfection of isolated liver segments in humans. There are still some shortcomings. Observed transfection efficacies in these models were less than in rodents and procedures are invasive in nature, which limits their applicability to patients. To elucidate these approaches it would be crucial to delimit the targeted segment of the liver and ensure protein secretion in the therapeutic range. Here, we report FIX expression up to 600–700% of normal human FIX levels. We could show that the expressed protein was fully functional (Fig. 1F). Taking into account that in hemophilia therapy low continuous levels (above 1%) can avoid major long-term complications such as debilitating arthropathy (Lofqvist et al., 1997), 1% or less of liver tissue should be sufficient to secrete FIX in a range considered therapeutic.
Loss of expression after MC and pV transfection is caused by transgene loss and to a lesser degree by gene silencing
For clinical applicability, ensuring durable expression is critical. M. Kay and coworkers showed that silencing of nonviral expression cassettes by covalently linked bacterial DNA is mediated by histone modifications in the chromatin structure and independent of CpG methylation of plasmid bacterial backbone DNA (Chen et al., 2004, 2008; Riu et al., 2007). This explains the superior duration of expression when using MC compared with pV. However, the reasons behind the observed expression loss of both MC and pV vectors that are designed for long-term expression have not been analyzed previously. Therefore the highly optimized HCR/hAAT-FIX expression cassette (Miao et al., 2000) was used and introduced into both pV and MC vector systems. In our hands, there was a decline in expression between day 14 and day 100 after gene transfer for both vector types in FIX knockout and C57BL/6 mice (Figs. 1 and 2). A major part of this effect can be attributed to vector loss, which was similar in pV- or MC-treated groups of mice (Fig. 2A). The vector gene copies per cell on day 14 and day 100 were similar for pV- and MC-treated mice, suggesting that the smaller size of the MC did not lead to improved gene transfer or better persistence of the vector in the cells compared with pV at the doses tested.
(
We used day 14 for our analysis to ensure that transiently exuberant extracellular vector DNA had been degraded and could not interfere with DNA analysis. Although the persistence of extracellular vector DNA over time in the experiment cannot be excluded, the ratios of expression levels per vector genome for the MC-treated groups (Fig. 2B) differed only marginally between days 14 and 100 and were independent of the vector doses. Therefore, we observed a close correlation between both MC vector DNA and expression, suggesting that most of the DNA is intracellular, or has similar kinetics as internalized DNA responsible for gene expression. An additional effect caused by silencing was observed only in pV-treated mice (p < 0.05). An inhibitory antibody response against FIX was excluded by ELISA against all major immunoglobulin subclasses (data not shown).
Subtle differences in methylation pattern of MC and pV might contribute to vector silencing
In a second step, we analyzed the cytosine methylation status of a 500-bp fragment consisting of the hAAT promoter and the 5′ UTR of the FIX expression cassette containing 26 CpGs (Fig. 3). The hepatic control region (HCR) was not included because of the lack of CpG sites. In total, six mice per vector type (three with a 3.7-pmol vector dose and three with an 18.5-pmol vector dose) were included at each time point. Fifteen clones (5 per independent DNA preparation) per mouse were picked and subjected to further analysis. Bisulfite-mediated conversion results in a C-to-T conversion of all unmethylated cytosines in the sample DNA. Methylated cytosines are protected and remain cytosines in the processed DNA. Sequencing resulted in 46 properly converted gene sequences for the MC day 14 group, in 52 sequences for the MC day 100 group, and in 36 and 66 sequences for the pV day 14 and day 100 groups, respectively. In general, bisulfite conversion of nonmethylated CpGs was the predominant pattern, similar to vector before treatment (Fig. 3A). Although the methylation events are rare and would suggest no biologically relevant effect, there was a statistically significant accumulation of cytosine methylation in a 188-bp segment (CpGs 7–9) containing the HNF1 and HNF4 binding sites of the hAAT promoter (Fig. 3B) in the pV day 100 mice compared with other methylation sites (p < 0.05; Fig. 3). Ten of 66 clones (15.2%) in these mice showed a protected cytosine in this region (CpGs 7–9).
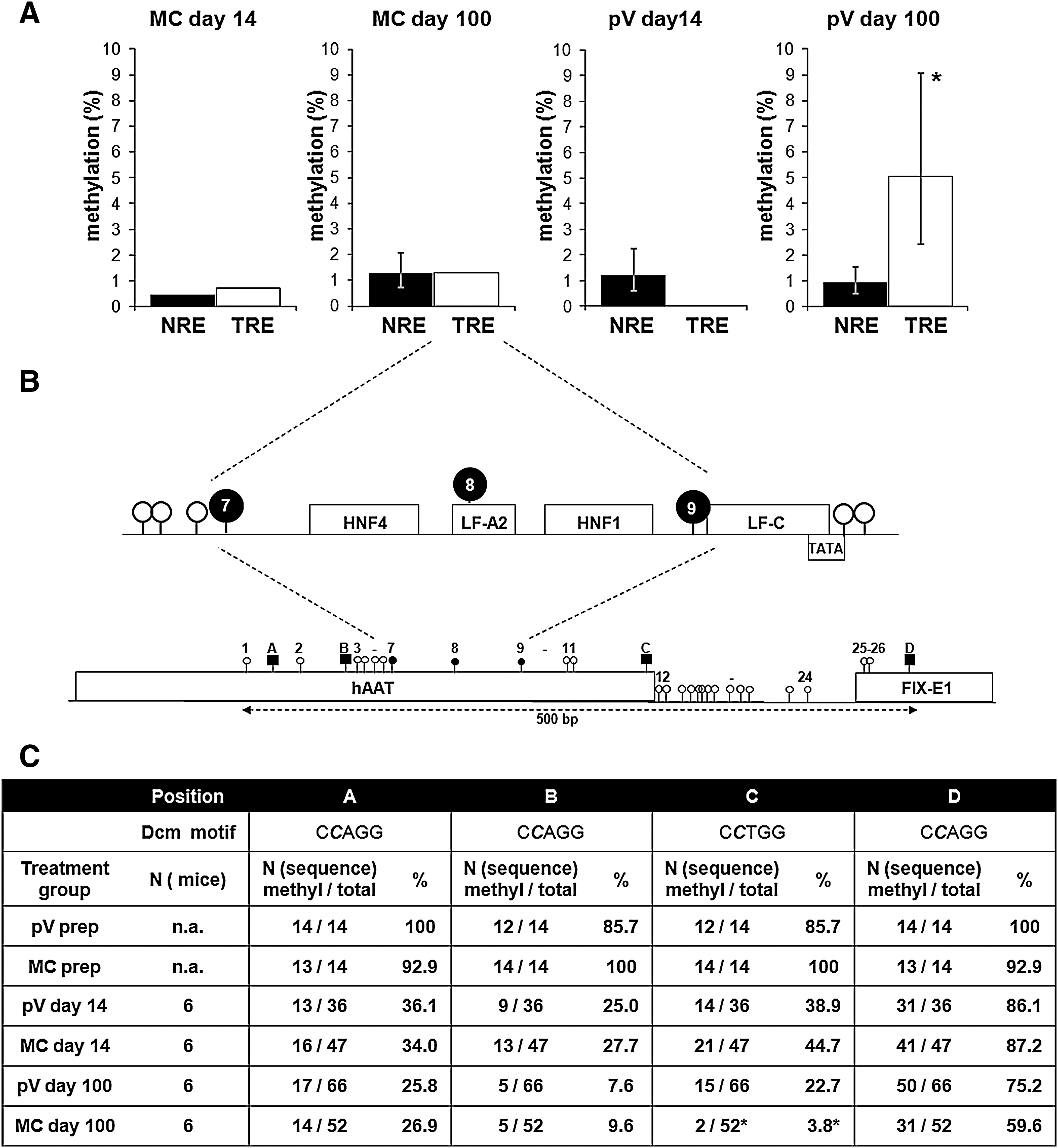
Methylation sites in the vector promoter region of pV- and MC-treated mice 14 and 100 days after vector treatment. (
As expected, we observed cytosines protected from conversion due to bacterial methylation at Dcm recognition sites (CmCTGG or CmCAGG; Fig. 3B and C). Before injection, all sites were protected; as soon as 14 days postinjection unmethylated cytosines were predominant. Demethylation over time depended on the location of the Dcm site. The three sites in the hAAT promoter showed much faster demethylation compared with the site located in exon 1 of the FIX gene (Fig. 3C). Furthermore, up to day 14, the process seemed comparable in MC- or pV-treated animals. However, on day 100, fewer remaining methylated cytosines were observed at positions C and D in the MC group (Fig. 3C). Although the functional importance of these changes is not clear at the moment, it shows that the methylation status of nonviral vector constructs is modified over time and could contribute to vector expression duration. On day 100 all sequences in the plasmid vector-treated group had at least one remaining bacterially methylated cytosine, suggesting that DNA was still of bacterial origin.
The methylation analysis detailed in this study is different in scope from the method previously described for examining CpG methylation of the bacterial backbone in plasmid vectors (Chen et al., 2008). It is believed that methylation can affect transcription factor binding and promote further methylation spreading and silencing (Turker, 2002). De novo DNA methyltransferases Dnmt3a and Dnmt3b have been proposed to be involved in promoter methylation of nondividing cells in cancer development (Hervouet et al., 2009). A similar mechanism could be involved in the silencing of transgene promoters. Nevertheless, CpG mutations as a strategy to improve long-term expression in this kind of promoter region affect only a few positions and should be considered an additional feature to ensure long-term transgene expression.
With the data obtained in our study, we clearly confirm that MC vectors ensure more durable FIX expression after hydrodynamic liver-directed gene transfer to the liver and that the high levels of functional protein obtained raise hope for future clinical developments. We identified the loss of vector transgenes over time as the main determinant for expression decline. Other processes, such as methylation of the promoter region, might play an additional role in this context. Our findings might help in the development of improved nonviral vectors for gene therapy of hemophilia B.
Footnotes
Acknowledgments
This work is part of the doctoral thesis of P.M. The research was supported by the Stiftung Hämotherapie-Forschung and by a grant from the JW Goethe University Clinics to J.S. S.R. and D.A. are students within the graduate study program GK-1172 funded by the DFG.
Author Disclosure Statement
No competing financial interests exist.