
Abstract
Indoleamine 2,3-dioxygenase (IDO) has been known as an emerging therapeutic target in autoimmunity-related arthritis. The treatment responses of adenoviral vectors encoding IDO (AdIDO) gene therapy in rat collagen-induced arthritis (CIA) were examined in this study. The therapeutic effects on ankle circumference, articular index, and radiographic and histological scores were evaluated in AdIDO-injected ankle joints. We further determined CD4+ T-cell numbers and their apoptotic status, CD68+ macrophage numbers, kynurenine (a downstream tryptophan metabolite) concentrations, interleukin-17 (IL-17) levels, and retinoic acid-related orphan receptor γt (RORγt) expression in synovial tissues of CIA rats receiving AdIDO treatment. Reduction of ankle circumference, articular index, and radiographic and histological scores were noted in AdIDO-treated ankles, as compared with those receiving injection of control vectors. Furthermore, IDO gene transfer led to decreased infiltrating CD4+ T cells with enhanced apoptosis, reduced CD68+ macrophage numbers, increased kynurenine levels, lower IL-17 concentrations, and decreased RORγt expression within the ankle joints. In addition, such a therapy diminished type II collagen-specific IL-17 production and RORγt expression in CD4+ T cells from draining lymph nodes of CIA rats. Our results demonstrate for the first time that intra-articular delivery of IDO gene ameliorated ankle arthritis of CIA rats by induction of CD4+ T-cell apoptosis and reduction of synovial IL-17 production through the supplement of kynurenine. Taken together, these findings implicate the novel strategy of using IDO gene as a therapeutic approach in treating patients with rheumatoid arthritis.
Introduction
Previous reports have put emphasis on tryptophan metabolism in human diseases such as RA and certain skin disorders, and improvement of arthritis has long been noted after the low-tryptophan diet therapy (Spiera and Lefkovits, 1967; Auckland, 1969; Houpt et al., 1973). Indoleamine 2,3-dioxygenase (IDO), a monomeric heme-containing enzyme, catalyzes the opening of the pyrrole ring of L-tryptophan and yields the product kynurenine (Higuchi and Hayaishi, 2008). Earlier studies of synovial fluid from a variety of arthritis patients including RA and osteoarthritis revealed the highest IDO activity with lowest tryptophan concentrations in RA samples, as well as an inverse correlation between these two substances (Bertazzo et al., 1999). Although a high IDO activity has been demonstrated in RA patients, the concentrations of kynurenine did not differ from that of the control group (Schroecksnadel et al., 2003). A recent report demonstrated the therapeutic effect of tryptophan immunoadsorption columns, possibly through the mechanism that raises kynurenine concentrations due to degradation of the in situ tryptophan adsorbed inside the columns by high plasma IDO activity in RA patients (Thomaidis et al., 2009). Therefore, up-regulation of kynurenine levels within rheumatoid joints by further increasing the intra-articular IDO activity could be a novel strategy in RA therapy.
The activation of CD4+ T cells has been shown to be inhibited by IDO through induction into apoptosis status (Mulley and Nikolic-Paterson, 2008). In addition to the effect of cytotoxic T-lymphocyte–associated antigen 4-IgG1 (CTLA4Ig) on the induction of T-cell anergy within rheumatoid joints, such a blockade can also up-regulate IDO activity in dendritic cells and further lead to down-regulation of T-cell proliferation in RA patients (Grohmann et al., 2002; Cron et al., 2005). The regulatory role of IDO in the CIA mice has been clearly demonstrated (Szanto et al., 2007; Bianco et al. 2009; Criado et al., 2009). Increased IDO activity in mice failing to develop arthritis and exacerbation of arthritis through inhibition of its activity indicated the feasibility of IDO supplement in suppressing arthritis (Szanto et al., 2007). IDO-transfected bone marrow-derived dendritic cells were capable of reversing established arthritis in CIA mice (Bianco et al., 2007). Furthermore, a recent study using the IDO-deficient mice demonstrated that kynurenine deficiency could initially induce the production of interferon-γ (IFN-γ) and IL-17, and later enhance the infiltration of Th1 and Th17 into inflamed joints (Criado et al., 2009).
Gene therapy, a method bypassing the inherited obstacle associated with the transfer of therapeutic compounds, has opened an important avenue greatly improving on the limitation of protein delivery treatment. By using intra-articular gene transfer into local inflamed joints, clinical improvements of arthritis were observed (Adriaansen et al., 2006). The RA gene therapy has progressed to the completion of phase II trials and provided a glimpse of clinical efficacy (Wehling et al., 2009). In the present study with a novel therapeutic strategy, i.e., supplement of tryptophan metabolites such as kynurenine through intra-articular adenoviral vector-mediated IDO gene therapy, we evaluated the therapeutic effect on CIA rats and further examined their responsible pathogenetic mechanisms including apoptosis of CD4+ T cells and production of synovial IL-17.
Materials and Methods
Construction of recombinant adenoviral vectors
IDO gene and its corresponding short-hairpin RNA gene (shIDO) driven by human RNA polymerase III U6 (HsU6) promoter were constructed into pShuttle-CMV and pShuttle vectors, respectively (Wu et al., 2005; Lu et al., 2006; Yen et al., 2009). Recombinant adenoviral vectors were further generated by cotransfection of PmeI-cut pShuttle-CMV-IDO or pShuttle-HsU6-shIDO linearized vectors with the adenoviral backbone vector pAdeasy-1 (E1/E3-deleted adenovirus type 5; Stratagene, La Jolla, CA) into BJ5183 cells (E. coli bacteria). The colonies of recombinant adenoviral vectors were further tested by restriction digest. Adenoviral plasmids carrying IDO (AdIDO) or shIDO (AdshIDO) were digested with PacI and further transfected into the 293 packaging cells (human embryonic kidney cells). The resulting high-titer adenoviruses were harvested 7 days later, and then further purified by CsCl density centrifugation and desalted by gel filtration through a Sephadex G-25 column (PD-10 column; GE Healthcare, Piscataway, NJ) (Hsieh et al., 2009). The adenoviral titers [plaque-forming units (PFU) per milliliter] were measured by standard plaque-forming assay using 293 cells. The AdLacZ, an adenoviral vector encoding β-galactosidase, was the control vector for the following experiments.
Cell viability, in situ apoptosis, and immunofluorescent staining
RA synovial fibroblasts (RASFs; 5 × 103/well) (synovial tissues were obtained during the orthopedic surgery; the Institutional Review Board of National Cheng Kung University Hospital approved this study) were seeded into 96-well dishes and then infected with AdLacZ, AdIDO plus AdLacZ, or AdIDO plus AdshIDO at a total multiplicity of infection (MOI) of 100 or left uninfected for 72 hr. In addition, human RASFs were prepared and their proliferations were assessed as previously described (Wang et al., 2008). The supernatants of each group were collected and incubated with Jurkat T cells (human leukemia T cells; ATCC, Manassas, VA) for another 72 hr. Their proliferation was analyzed by using the MTT assay with measurement of absorbance at 570 nm. Cell viability was represented as the percentages normalized with the average MTT values of the mock group.
Cryostat sections (5 μM) were prepared from ankle joints receiving different treatments on day 16 for detection of in situ apoptotic CD4+ T cells in synovial tissues. The DeadEnd fluorometric terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) assay kit (Promega, Madison, WI) was used for this experiment according to the standard protocol. Each cryostat section was further incubated with mouse anti-rat CD4 antibodies (BD Biosciences, San Diego, CA) for 1 hr and then with rhodamine-conjugated goat anti-mouse IgG (H + L) for 30 min (KPL, Baltimore, MD). Cells with double stainings of TUNEL and CD4 were identified and counted in three high-power (400 ×) fields to determine the average percentages of CD4+ apoptotic cells in each section under fluorescence microscopy.
Measurement of kynurenine
L-Kynurenine concentrations in joint extracts or cell supernatants were quantified according to previous reports with modifications (Matin et al., 2006; Criado et al., 2009). In brief, after the synovial tissue extracts or cell supernatants were heated at 60°C for 15 min, 100 μl of each was added with an equal volume of IDO assay buffer (50 mM potassium phosphate, 20 mM ascorbic acid, 10 μM methylene blue, 100 μg/ml catalase). After 40 μl of 30% (w/v) trichloroacetic acid was added, these assay mixtures were centrifuged at 3,000 g for 15 min. Then 125 μl of the supernatants were added to an equal volume of 2% (w/v) Erhlich's reagent (4-dimethylaminobenzaldehyde in acetic acid), and the absorbance was measured at 480 nm to quantify the concentrations of L-kynurenine. In addition, a standard curve of L-kynurenine was processed in parallel with the above experimental procedures.
Induction of CIA and intra-articular AdIDO gene therapy
CIA was induced in Sprague-Dawley rats by two dorsal intradermal injections with bovine type II collagen (CII) on days 0 and 7 (Wang et al., 2005, 2008; Shiau et al., 2007; Chen et al., 2009). Each ankle of the rats was then injected intra-articularly with 5 × 107 PFU of AdIDO or AdLacZ on day 7. In addition, the phosphate-buffered saline (PBS)-injected ankle joints served as a control group. Articular index scores and ankle circumferences were monitored up to day 16, and radiographic and histological changes of the ankle joints were assessed on day 16. All animal experiments were done following the guidelines approved by the Laboratory Animal Care and Use Committee of the National Cheng Kung University.
Immunohistochemical staining, immunoblot analysis, and enzyme-linked immunosorbent assay (ELISA)
Sections of paraffin-embedded ankle joints were deparaffinized in xylene, dehydrated in serial alcohol concentrations, treated with protease K to unmask the epitopes, and then washed with 3% PBS-buffered H2O2. They were then immunostained with mouse anti-rat CD4 antibodies (BD Biosciences) or mouse anti-rat CD68 macrophage/monocyte antibodies (ED1; Chemicon, Temecula, CA), followed by peroxidase-labeled secondary antibodies, and aminoethyl carbazole was used as the substrate chromogen. The CD4+ and CD68+ cells were counted in three high-power (400 × ) fields to determine the average percentages of CD4+ T cells and CD68+ macrophages in each section.
RASFs transfected with AdLacZ, AdIDO plus AdLacZ, or AdIDO plus AdshIDO at a total MOI of 100 or left uninfected for 48 hr were further subjected to the first experiment of IDO immunoblot. Ankle-joint extracts from CIA rats receiving PBS, AdLacZ, or AdIDO injection for 72 hr were also analyzed with IDO immunoblot. Cell lysates or joint extracts were examined for their IDO expression by mouse anti-rat IDO antibodies (Chemicon). The blots were then reprobed with anti-β-actin antibodies (Sigma-Aldrich, St. Louis, MO) as quantitative controls. Synovial tissues taken from the ankle joints of CIA rats were snap-frozen and homogenized for further ELISA. Concentrations of IL-17, IL-6, and IL-1β in ankle-joint homogenates were determined by using commercial ELISA kits for quantification of rat cytokines (eBioscience, San Diego, CA). Total protein concentrations were determined by using a BCA protein assay kit (Pierce, Rockford, IL).
CII-stimulation assay, cell sorting, and reverse transcription-PCR (RT-PCR) analysis
Single-cell suspensions of draining inguinal lymph nodes (LNs) from CIA rats treated with AdLacZ, PBS, or AdIDO were prepared and cultured in the presence or absence of 100 μg/ml denatured CII. After 72 hr, 100 μl of the supernatants were collected for further measurement of IL-17 by using ELISA kits. LN cells (2 × 107) were doubly stained with phycoerythrin-conjugated anti-CD4 (BD Biosciences) and fluorescein isothiocyanate-conjugated anti-CD3 antibodies (BD Biosciences), and then 5 × 106 CD3+CD4+ double-positive cells were sorted by the FACSAria cell sorter (BD Biosciences).
The expressions of retinoic acid-related orphan receptor γt (RORγt) or IDO in double-positive cells, synovial tissues, or LN cells were detected by RT-PCR. Total RNA was isolated by the TRIzol reagent (Invitrogen, Carlsbad, CA). The REVERSE-IT first-strand synthesis kit (ABgene, Epsom, UK) was used for the cDNA synthesis. The forward and reverse PCR primers of RORγt or IDO were either 5’-CATCTCTGCAAGACTCATCGACA-3’ and 5’-TTTCCACATGTTGGCTGCA-3’, or 5’-TGTGGCTAGAAATCTGCCTGT-3’ and 5’-CTGCGATTTCCACCAATAGAG-3’. The conditions of PCR for RORγt or IDO were either 50 cycles of 20 sec at 94°C, 20 sec at 51°C, and 20 sec at 72°C, or 30 cycles of 30 sec at 94°C, 30 sec at 55°C, and 60 sec at 72°C.
Statistical analysis
Data are expressed as means ± SEM. Differences in ankle circumferences and articular index among various treatment groups were compared by the repeated-measures analysis of variance using SAS software version 9.1 (GLM program; SAS Institute, Cary, NC). Differences in the remaining data obtained from two groups were compared with Student's t test. P values less than 0.05 were considered significant in this study.
Results
Reduction of Jurkat T-cell proliferation through the up-regulation of kynurenine mediated by IDO gene transfer
Biological functions of viral vectors AdIDO and AdshIDO were examined. RASFs were transfected with the mock condition, AdLacZ (MOI = 100), AdIDO (MOI = 50) plus AdLacZ (MOI = 50), or AdIDO (MOI = 50) plus AdshIDO (MOI = 50). IDO expression was clearly up-regulated in AdIDO-infected RASFs and down-regulated in AdIDO plus AdshIDO ones (Fig. 1A). Kynurenine concentration was increased in the supernatant of AdIDO-infected RASFs and decreased in AdIDO plus AdshIDO ones (Fig. 1B) (AdLacZ vs. AdIDO, 0.8610 ± 0.02214 vs. 5.021 ± 0.3814, p = 0.0004; AdshIDO vs. AdIDO, 1.193 ± 0.1594 vs. 5.021 ± 0.3814, p = 0.0008). We further examined the endogenous IDO activity in RASFs and found a dose-dependent IFN-γ-induced up-regulation of kynurenine production in their supernatants (Fig. 1C).
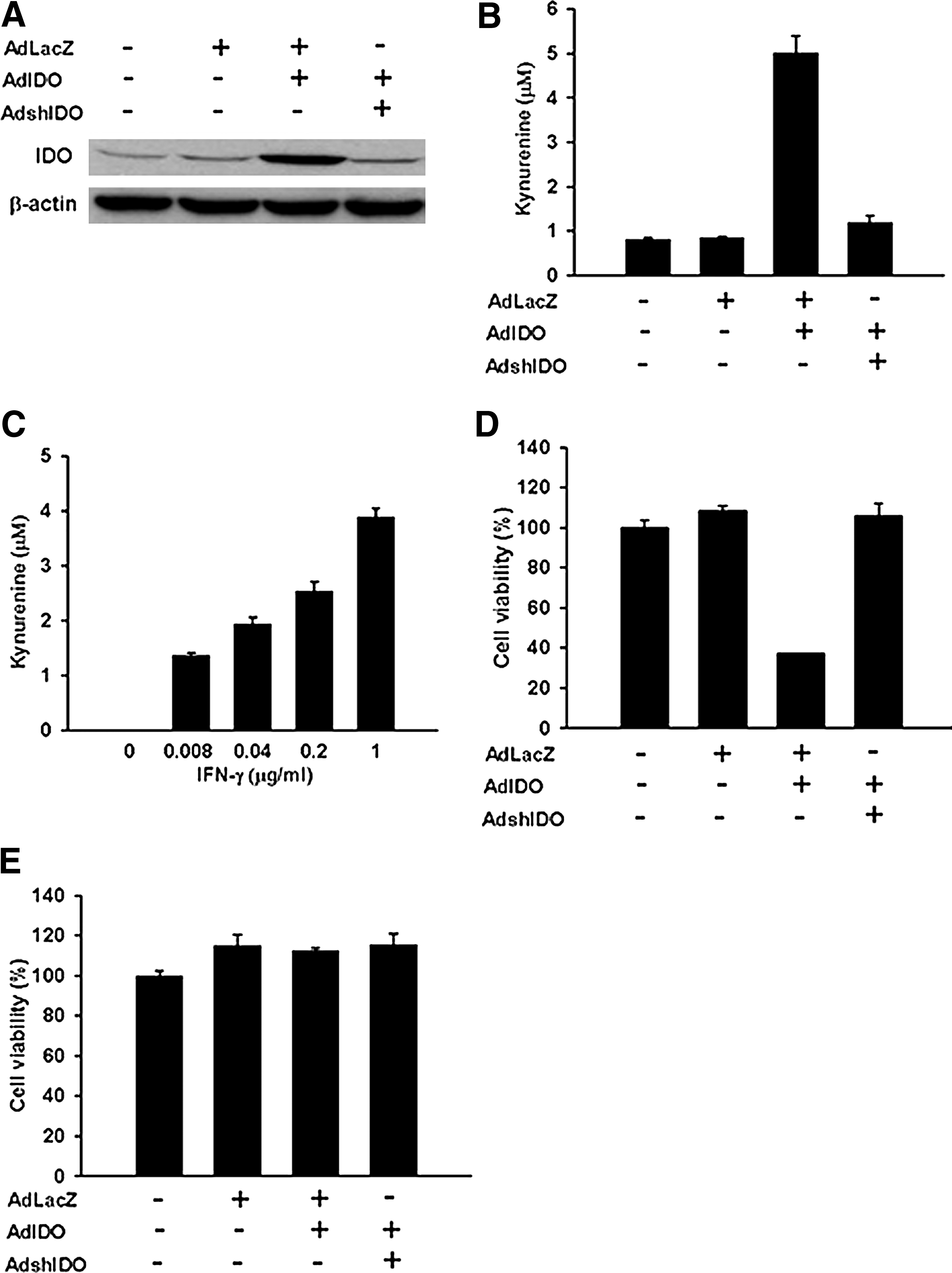
Characterization of biological functions of the viral vector AdIDO. RASFs were infected with AdLacZ (AdLacZ), AdIDO plus AdLacZ (AdIDO), or AdIDO plus AdshIDO (AdshIDO) for 48 hr.
Next, we infected RASFs with either AdIDO or AdIDO plus AdshIDO at a total MOI of 100. Supernatants collected from AdIDO-infected RASFs significantly reduced the viability of Jurkat T cells as compared with those infected with mock, AdLacZ, or AdIDO plus AdshIDO by using the MTT assay (Fig. 1D) (AdLacZ vs. AdIDO, 108.2 ± 2.45 vs. 37.17 ± 0.22, p < 0.0001; AdshIDO vs. AdIDO, 105.90 ± 6.05 vs. 37.17 ± 0.22, p = 0.0003). Cell viability of human RASFs was not affected after transfection with adenoviral vectors expressing LacZ, IDO, or IDO plus shIDO (Fig. 1E). Furthermore, the designed shIDO not only down-regulated IDO expression, but also reversed the inhibition of Jurkat T-cell proliferation mediated by IDO transfection (Fig. 1D).
Amelioration of CIA by AdIDO treatment
The therapeutic effects of AdIDO, AdLacZ, or PBS injection on ankle joints of CIA rats were evaluated. Both ankle circumference and articular index scores of the AdIDO-treated CIA rats were significantly lower than those of other control counterparts [ankle circumference (Fig. 2A): AdLacZ vs. AdIDO, p < 0.0001; PBS vs. AdIDO, p = 0.0189; normal control vs. AdIDO, p < 0.0001; and articular index (Fig. 2B): AdLacZ vs. AdIDO, p = 0.0008; PBS vs. AdIDO, p = 0.0134; normal control vs. AdIDO, p < 0.0001].

Amelioration of arthritis by overexpression of IDO gene in the ankle joints of CIA rats. Intra-articular injection of AdIDO into ankle joints on day 7 reduced the ankle circumference
We next assessed the radiographic and histological changes in their ankle joints on day 16. The representative plain radiographs revealed that AdIDO-treated rats had more intact joint structures, as compared with their control counterparts (Fig. 2C). The radiological scores based on joint space width, degree of bony destruction, and soft tissue swelling were significantly lower in the AdIDO-treated group than in other control groups (Fig. 2E) (AdLacZ vs. AdIDO, 3.83 ± 0.33 vs. 1.92 ± 0.24, p = 0.0009; PBS vs. AdIDO, 3.58 ± 0.30 vs. 1.92 ± 0.24, p = 0.0015).
To further confirm the clinical improvement of ankle arthritis, histological evaluations for synovial tissues were conducted. Histological analysis of either the AdLacZ- or PBS-treated joints revealed synovial hyperplasia, significant inflammatory cell infiltration, and obvious bone erosion; however, the joint tissues from AdIDO-injected ankles demonstrated relatively intact cartilages on the joint surface and only minimal hyperplasia of the synovial membrane (Fig. 2D). Notably, fewer infiltrating inflammatory cells were detected in the synovial tissues from the AdIDO-treated rats. Furthermore, the histological scores based on synovial hyperplasia, cartilage erosion, and leukocyte infiltration were significantly lower in the AdIDO-treated group than in other control groups (Fig. 2F) (AdLacZ vs. AdIDO, 4.92 ± 0.35 vs. 2.58 ± 0.35, p = 0.0009; PBS vs. AdIDO, 4.0 ± 0.29 vs. 2.58 ± 0.35, p = 0.011).
Taken together, these data indicated that AdIDO gene therapy ameliorated arthritis in CIA rats, as revealed by the results of clinical, radiographic, and histological examinations on their joints.
Induction of CD4+ T-cell apoptosis and reduction of CD68+ macrophage infiltration in ankle joints of CIA rats by AdIDO gene therapy
Immunohistochemical staining demonstrated a significant decrease in the numbers of CD4
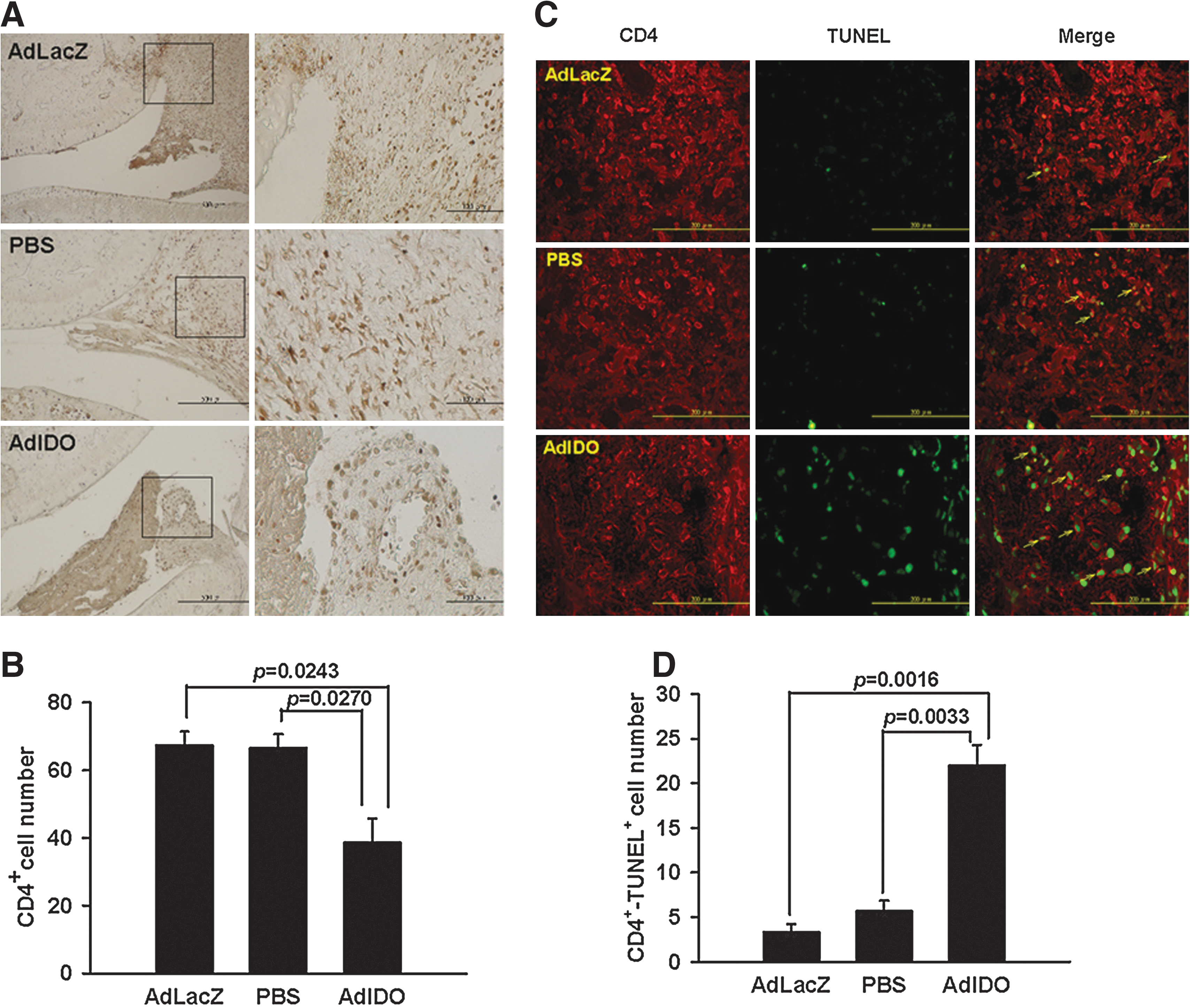
Reduction of CD4+ T-cell numbers with enhanced apoptotic status in the ankle joints of CIA rats.

Reduction in the infiltrating CD68+ macrophages in the ankle joints of CIA rats by IDO gene therapy.
Effects of AdIDO gene therapy on IL-17 concentrations, kynurenine levels, RORγt expression, and CII-specific IL-17 production in CIA rats
IDO levels in the draining LNs and ankle joints of CIA rats were analyzed, and significant expression was detected in the draining LN cells from day 8 onward by using the RT-PCR examination (Fig. 5A). However, IDO expression was lacking in ankle-joint extracts at a similar time point compared with expression in extracts of AdIDO-infected 293 cells as the positive control (data not shown). Nevertheless, our results were in agreement with recent studies demonstrating deficient IDO expression in arthritic joints and, moreover, reduction of arthritis in CIA mice by administration of kynurenine (Criado et al., 2009). Figure 5B shows significant IDO expression in ankle joints of CIA rats 72 hr after receiving AdIDO injection, as indicated by immunoblot analysis (Ijima et al., 2001). Figure 5C revealed higher synovial kynurenine levels in AdIDO-injected joints on day 16 as compared with other control counterparts (AdLacZ vs. AdIDO, 16.61 ± 3.97 vs. 46.10 ± 9.62, p = 0.0177; PBS vs. AdIDO, 15.16 ± 1.77 vs. 46.10 ± 9.62, p = 0.0101).
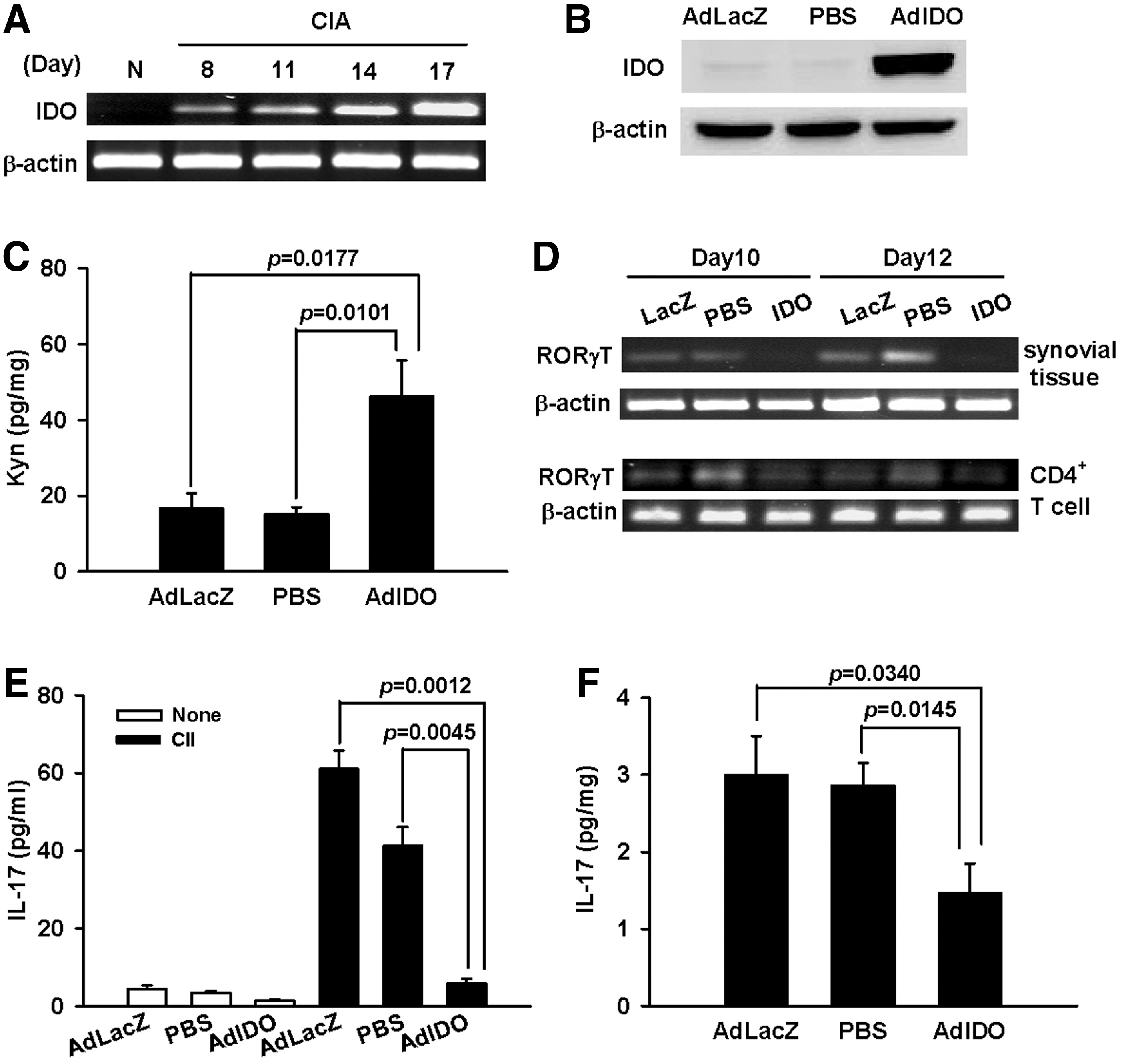
Effects of AdIDO gene therapy on kynurenine levels, RORγT expression, CII-specific IL-17 production, and IL-17 concentrations in CIA rats. CIA rats were either injected intra-articularly with 5 × 107 PFU of AdIDO, AdLacZ, or PBS on day 7 or left untreated during progression of arthritis. ankle-joint homogenates and inguinal LN cells were prepared and further subjected to ELISA, RT-PCR, or immunoblot analysis.
Reduced expression of RORγt was found in AdIDO-treated ankle extracts and in CD4+ T cells from the draining LNs after different treatments were given for 3 or 5 days (Fig. 5D). Furthermore, lower IL-17 concentrations were found in supernatants of draining LNs stimulated with CII from CIA rats 3 days after receiving AdIDO treatments (Fig. 5E) (AdLacZ vs. AdIDO, 61.02 ± 4.71 vs. 5.85 ± 1.21, p = 0.0012; PBS vs. AdIDO, 41.21 ± 4.87 vs. 5.85 ± 1.21, p = 0.0045). IL-17 levels in the synovial extracts were reduced in AdIDO-injected joints as compared with PBS- or AdLacZ-treated ankles (Fig. 5F) (AdLacZ vs. AdIDO, 3.0 ± 0.50 pg/mg vs. 1.48 ± 0.37 pg/mg, p = 0.0340; PBS vs. AdIDO, 2.87 ± 0.29 pg/mg vs. 1.48 ± 0.37 pg/mg, p = 0.0145).
In addition, IL-6 and IL-1β of the joint extracts were decreased in AdIDO-injected ankles as compared with other control groups (for IL-6: AdLacZ vs. AdIDO, 239.3 ± 27.29 pg/mg vs. 129.2 ± 26.76 pg/mg, p = 0.0164; PBS vs. AdIDO, 204.5 ± 20.27 pg/mg vs. 129.2 ± 26.27 pg/mg, p = 0.0488; for IL-1β: AdLacZ vs. AdIDO, 321.3 ±37.72 pg/mg vs. 164.4 ± 15.05 pg/mg, p = 0.0048; PBS vs. AdIDO, 253.9 ± 13.57 pg/mg vs. 164.4 ± 15.05 pg/mg, p = 0.0022).
Collectively, these data indicated that induction of CD4+ T-cell apoptosis and reduction of synovial IL-17 production might play critical roles in reduction of arthritis by manipulation of tryptophan catabolism through intra-articular AdIDO gene therapy.
Discussion
Our studies clearly demonstrated the amelioration of ankle arthritis in CIA rats by intra-articular adenoviral vector-mediated IDO gene delivery. Furthermore, following the AdIDO gene therapy, there were increased kynurenine concentrations, reduction in CD4+ T-cell numbers with enhanced apoptosis, suppression of CD68+ macrophage infiltration, lower IL-17 production in ankle joints, decreased CII-specific IL-17 response in draining LNs, and reduced RORγt expression in either synovial tissues or CD4+ T cells from draining LNs. These novel identified mechanisms by which IDO gene therapy reduced arthritis in CIA rats appeared to be the induction of apoptosis in synovial RORγt-expressing CD4+ T cells that secrete IL-17 via increased kynurenine concentrations through the up-regulation of the tryptophan-degradation pathway.
Inflamed rheumatoid joints can be initiated by infiltrating CD4+ T cells through the activation of synoviocytes to release pro-inflammatory cytokines such as IL-1β, IL-6, and TNF-α (Klareskog et al., 2009). Despite the fact that abundant pro-inflammatory monokines are present within rheumatoid synovial tissues, T helper-related cytokines, IFN-γ (Th1) and IL-4 (Th2), are barely detected in inflamed joints from RA patients (Choy and Panayi, 2001). Several recent studies have demonstrated elevated IL-17 levels in synovial CD4+ T cells and synovial fluid samples from rheumatoid joints (Chabaud et al., 1999; Miossec, 2007; Shahrara et al., 2008; Annunziato et al., 2009). Through the induction of cytokines, chemokines, and mediators from synoviocytes and infiltrating mononuclear cells, IL-17 is capable of promoting bone erosion in inflamed joints (Chabaud et al., 1999; Miossec, 2007). Targeting the IL-17 signaling pathway through the induction of apoptosis in synovial Th17 cells could be a potential therapeutic strategy in RA patients (Ivanov and Linden, 2009). Indeed, a very recent report of treating rheumatoid joints with anti-IL-17 monoclonal antibody has demonstrated clinical efficacy without adverse events, which strongly supports a novel therapeutic role of IL-17 neutralization in RA therapy (Genovese et al., 2010).
Accumulated evidence reveals increased degradation of tryptophan due to up-regulated IDO activity in RA patients, and such a phenomenon could be the physiological defense against over-reactive autoimmunity (Bertazzo et al., 1999; Schroecksnadel et al., 2003). Earlier studies have demonstrated the clinical efficacy of IFN-γ, a strong IDO-inducing cytokine, in RA therapy possibly by lowering tryptophan levels and subsequently slowing down the T-cell activation (Lemmel et al., 1987). More recently, tryptophan reduction has been proposed as a novel T-cell down-regulation strategy in treating RA patients (Schroecksnadel et al., 2003). Up-regulation of IDO expression in the draining LNs but not in arthritic joints of CIA rats has been shown in this study and a previous report (Fig. 5A; Criado et al., 2009). Furthermore, our studies using exogenous supplement of IDO in ankle joints through adenoviral gene transfer revealed the amelioration of arthritis in CIA rats; these results were consistent with the findings from previous studies that IDO-deficient CIA mice had exacerbated arthritis severity compared with IDO-competent mice (Criado et al., 2009).
The reported mechanisms of IDO-mediated T-cell survival include the induction of T-cell apoptosis and conversion of naive T into T-regulatory (Treg) cells (Mulley and Nikolic-Paterson, 2008; Izcue et al., 2009). Moreover, the tryptophan catabolite kynurenine was shown to support the generation of forkhead box P3 (FOXP3)+ Treg cells and suppress RORγt-expressing Th17 cells in pathological inflammatory status (Romani et al., 2008). Reduced RORγt expression in both synovial tissues and CD4+ T cells from draining LNs after AdIDO gene therapy has been demonstrated in this study; however, we failed to identify FOXP3 expression in ankle joints and inguinal LNs of our rat CIA model (data not shown). IDO-expressing CD11c+ dendritic cells from Peyer's patches were shown to be able to suppress the CII-specific T-cell proliferation and down-regulation of Th1 cytokine production through the generation of FOXP3+ Treg cells in an orally tolerized mouse CIA model (Park et al., 2008). Our novel strategy using local transfer instead of systemic delivery of AdIDO gene would most likely cause the apoptosis of synovial Th17 cells rather than induce the generation of circulating Treg cells. CD8+ T cells have been shown to resist both in vitro and in vivo apoptosis induction by the addition of tryptophan catabolites such as kynurenine into cellular cultures and the overexpression of IDO in a rodent allograft rejection model through adenovirus-mediated IDO gene delivery (Weber et al., 2006; Liu et al., 2007). Indeed, most of the TUNEL-positive cells in AdIDO-treated synovial tissues from rat ankle joints were doubly stained with CD4 (Fig. 3C).
Taken together, our findings demonstrate for the first time that in vivo intra-articular delivery of IDO gene ameliorated arthritis in CIA rats by induction of CD4+ T-cell apoptosis and reduction of synovial IL-17 production through the supplement of kynurenine, a downstream tryptophan metabolite. These results further implicate the novel strategy of using IDO gene therapy to manipulate tryptophan catabolism as a potential clinical approach for RA therapy.
Footnotes
Acknowledgments
This work was supported by the National Science Council (NSC grants 97-2314-B-006-007-MY3 and 98-2628-B-006-014-MY3), the Department of Health, Executive Yuan (grant DOH 96-TD-I-111-TM014), and the National Cheng Kung University Landmark Project (grant NCKU-97-R010), Taiwan.
Author Disclosure Statement
No competing financial interests exist.