
Abstract
The regenerative potential of bone marrow-derived endothelial progenitor cells (EPCs) has been adapted for the treatment of myocardial and limb ischemia via ex vivo expansion. We sought to enhance EPC function by the efficient genetic modification of EPCs in a rat model of myocardial infarction. Peripheral blood EPCs were expanded and transduced, using adeno-associated virus (AAV). AAV-mediated EPC transduction efficacy was 23 ± 1.2%, which was improved by 4.0- to 7.2-fold after pretreatment with the tyrosine kinase inhibitor genistein. Adult rats (n = 7 in each group) underwent myocardial infarction by left anterior descending coronary artery occlusion, and received autologous EPCs transduced by AAV-IGF-1 or AAV-lacZ into the periinfarct area. Echocardiography demonstrated that cardiac function in the IGF-1-EPC group was significantly improved compared with the lacZ-EPC control group 12 weeks after myocardial infarction. In addition, IGF-1-expressing EPCs led to reduced cardiac apoptosis, increased cardiomyocyte proliferation, and increased numbers of capillaries in the periinfarct area. AAV expression was limited to the targeted heart region only. Pretreatment with genistein markedly improved AAV transduction of EPCs. IGF-1-expressing EPCs exhibit favorable cell-protective effects with tissue-limited expression in rat heart postinfarction.
Introduction
Adeno-associated viral (AAV) vectors, which are derived from a nonpathogenic human parvovirus, have been widely applied as vectors for systemic and local gene delivery (Manno et al., 2006; Aiuti et al., 2007; Van Vliet et al., 2008). The physical stability of AAV makes it advantageous for in vivo use, and transgene expression can persist long term in a wide range of tissues including heart muscle (Aikawa et al., 2002; Inagaki et al., 2006; Liu et al., 2006; Van Vliet et al., 2008). In addition, we have demonstrated that AAV vectors are capable of achieving safe and efficient transduction of bone marrow cells without abolishing cell function when used in in vitro and in vivo models (Iwakura et al., 2005; Chng et al., 2007).
Insulin-like growth factor (IGF)-1 is a 7.6-kDa polypeptide growth factor expressed by many cells and tissues during embryonic and postnatal development (Liu and LeRoith, 1999; Khan et al., 2002; Delafontaine et al., 2004). IGF-1 influences various biological processes through binding to a membrane–anchored receptor (IGF-1R), although at higher concentrations, IGF-1 can also activate the insulin receptor (Khan et al., 2002). The in vivo action of IGF-1 is modulated by a family of circulating IGF-binding proteins (Delafontaine et al., 2004). IGF-1 is secreted from the liver in response to growth hormone (GH) and promotes cellular proliferation in bone, muscle, fat, and other tissues (Liu and LeRoith, 1999; Khan et al., 2002; Delafontaine et al., 2004). Human and murine IGF-1 deficiency causes severe intrauterine growth retardation, perinatal lethality, postnatal growth retardation, and delayed development in a variety of organs (Liu and LeRoith, 1999), indicating that IGF-1 is a key regulator of cell development. Overexpression of IGF-1 attenuates myocyte necrosis and apoptosis after infarction in a transgenic mouse model (Li et al., 1997) and rescues cardiac myocytes from apoptosis in dilated cardiomyopathy (Welch et al., 2002). It has also been reported that overexpression of IGF-1 reduces cardiomyocyte senescence in a transgenic mouse model (Torella et al., 2004) and mice overexpressing IGF-1 exhibit less cardiomyocyte atrophy after ischemia (Schulze et al., 2005). Taken together, IGF-1 is a pleiotropic growth factor that induces myocyte proliferation and regeneration in heart development and injury. We have reported that local IGF-1 overexpression markedly improved cardiac function after myocardial infarction (MI) through direct AAV injection to heart muscle (Dobrucki et al., 2010).
The aims of this study were to further develop cardiac therapies, using genetically modified peripheral blood-derived EPCs. We sought to investigate whether transplantation of EPCs expressing IGF-1 after ex vivo AAV gene transfer improved cardiac function in rats post-MI.
Materials and Methods
Materials
Genistein, hydroxyurea (HU), N-acetyl-
AAV vector production
Standard recombinant DNA approaches were used to incorporate the human IGF-1 cDNA (NCBI database, accession number X00173) open reading frame under the control of the cytomegalovirus (CMV) promoter as previously described (Kusano et al., 2007). Briefly, human IGF-1 cDNA was amplified by polymerase chain reaction (PCR) with the following specific primers: 5′-CCGAATTCTTCAGAAGCAATGGGA-3′ and 5′-CGGGATCCGTCTTCCTACATCCTG-3′, from a human gene clone (clone ID 984882, cat. no. 97002RG; Invitrogen Life Technologies, Carlsbad, CA) (Dobrucki et al., 2010). AAV vectors of serotype 2 were prepared at a titer of 1 × 1012 vector genomes/ml as shown previously (Kusano et al., 2007).
AAV transduction and detection
Mononuclear cells were isolated from rat peripheral blood by density-gradient centrifugation, and then cultured in endothelial cell (EC) basal medium-2 (Clonetics EBM-2; Lonza Walkersville, Walkersville, MD) for 7 days. The majority of these ex vivo-expanded cells were of endothelial lineage and, as such, they constituted the ex vivo-expanded EPC-enriched fraction (Kawamoto et al., 2001). EPCs were transduced with AAV-CMV-lacZ (titer, 3 × 103 genomes per cell) or AAV-CMV-IGF-1, fixed in by 4% paraformaldehyde (PFA) 1 week later, and stained for β-galactosidase (β-Gal) activity (Aikawa et al., 2002). Quantification of β-Gal activity in these cells was performed with a mammalian β-galactosidase assay kit (Pierce/Thermo Fisher Scientific, Waltham, MA).
To detect the AAV genome, total DNA was extracted from tissues, using a Gentra Puregene DNA isolation kit (Qiagen, Valencia, CA). A fragment of the AAV genome (286 bp) was amplified, using a forward primer within the inverted terminal repeat (ITR) region and a reverse primer within the CMV promoter as described previously (Iwakura et al., 2005).
RT-PCR for mRNA expression
Total RNA was extracted from heart tissue with RNA STAT (Tel-Test, Friendswood, TX) according to the manufacturer's instructions. First-strand cDNA was generated with a TaqMan MultiScribe reverse transcription kit (Applied Biosystems, Foster City, CA) primed with a mix of oligo(dT) and random hexamers. Gene expression was determined by TaqMan real-time (RT) quantitative PCR on a 7300 sequence detection system (Applied Biosystems), using TaqMan PCR master mix (Applied Biosystems). TaqMan primer–probe sets to detect IGF-1 expression (Biosearch Technologies, Novato, CA) were designed with Primer Express software (Applied Biosystems) with the following sequences: forward, 5′-CCATGTCCTCCTCGCATCTC; reverse, 5′-CGTGGCAGAGCTGGTGAAG; probe, 5′-ACCTGGCGCTGTGCCTGCTCA. PCR conditions were as follows: hold for 2 min at 50°C, 10 min at 95°C, followed by a two-step PCR for 40 cycles of 95°C for 15 sec and 60°C for 60 sec with fluorescence monitoring at the end of each elongation step. Relative mRNA expression of target genes was calculated by the comparative C T method (Kusano et al., 2007). All target sequences were normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) in multiplexed reactions performed in duplicate. Differences in C T values were calculated for each target mRNA by subtracting the mean value of GAPDH (Kusano et al., 2007).
Quantification of cell number, proliferation, and apoptosis
The MTT-formazan assay (Vistica et al., 1991) was used to assess the metabolic activity of cultured cells according to the instructions of the manufacturer (Sigma-Aldrich). The optical density of the solution was measured at 545 nm with a microwell plate reader. Rat EPCs transduced with AAV were cultured with neonatal rat cardiomyocytes (1:4 ratio) for 7 days (Yoon et al., 2005). For detection of apoptosis, the cells were first probed with an anti-α-actinin antibody (cat. no. A7811; Sigma-Aldrich). Nuclear staining for DNA fragmentation was performed by the terminal deoxynucleotidyltransferase-mediated dUTP nick end labeling (TUNEL) method (cat. no. 11684795910; Roche Molecular Biochemicals, Indianapolis, IN). Bromodeoxyuridine (BrdU) was detected with sheep anti-BrdU antibody (diluted 1:50) (cat. no. M20105S; BIODESIGN International, Saco, ME) followed by streptavidin–fluorescein isothiocyanate (FITC) (diluted 1:100) (cat. no. SA-5001; Vector Laboratories, Burlingame, CA). BrdU-positive cells were counted in the periinfarct area, where the scar tissue constituted less than 20% of the visual field (magnification, × 200) (Yoon et al., 2005).
Animal studies
All procedures were performed in accordance with Caritas St. Elizabeth's Medical Center (Boston, MA) Institutional Animal Care and Use Committee (Approval number 248). Sprague-Dawley rats (7- to 8-week-old males; Jackson Laboratory, Bar Harbor, ME) were anesthetized with an intraperitoneal injection of ketamine (40–90 mg/kg) and xylazine (5–10 mg/kg) and respiration was controlled with an animal ventilator during thoracotomy. Peripheral blood (1 ml) was obtained from the heart of each rat and EPCs were cultured ex vivo. One week after transduction, the EPCs were used for autologous transplantation. Myocardial infarction was induced by ligating the proximal left anterior descending coronary artery with a 6-0 Prolene suture. Under direct visualization, transduced EPCs (1 × 104 cells in 20 ml) were injected immediately into five sites of the myocardium surrounding the infarcted area, using a 30-gauge needle.
Echocardiography
Transthoracic echocardiography (SONOS 5500; Philips Healthcare, Amsterdam, The Netherlands) was performed on day 0 (before thoracotomy) and 12 weeks after myocardial infarction. Left ventricular diastolic dimension (LVDd), left ventricular systolic dimension (LVDs), and fractional shortening (FS) were measured at the mid-papillary muscle level (Kusano et al., 2007).
IGF-1 ELISA
Blood samples were taken periodically from the tail vein of anesthetized rats for the measurement of plasma human IGF-1 levels, using a human IGF-1 ELISA kit (Quantikine; R&D Systems, Minneapolis, MN) (Dobrucki et al., 2010).
Angiogenesis, apoptosis and proliferation in heart tissue
For the in vivo assay, hearts were harvested 12 weeks after myocardial infarction and transplantation of EPCs. To detect β-galactosidase, freshly excised tissues were flash frozen in O.C.T. compound (Sakura Finetek USA, Torrance, CA) and sectioned. Slides were incubated with an anti-β-Gal antibody (cat. no. G8021; Sigma-Aldrich) and anti-CD31 antibody (cat. no. 555445; BD Biosciences, San Jose, CA) for the detection of AAV-lacZ-transduced EPCs. Additional staining for capillaries was performed with rat-specific endothelial cell marker isolectin B4 (Vector Laboratories) as described previously (Dobrucki et al., 2010). Capillary density was scored morphometrically by histological examination of five randomly selected fields of tissue sections of periinfarct left ventricular myocardium. Capillaries were defined as tubular structures positive for isolectin B4. For apoptosis and proliferation assays, fixed heart tissue was first probed with α-actinin antibody (cat. no. A7811; Sigma-Aldrich). Nuclear staining for DNA fragmentation was performed by the TUNEL method to detect apoptosis, and Ki-67 staining was performed with rabbit polyclonal antibody against Ki-67 (cat. no. NCL-Ki67p; Novocastra Laboratories, Newcastle, UK) for cell proliferation, followed by 4′,6-diamidino-2-phenylindole (DAPI) staining (cat. no. 10236276001; Roche) to identify the number of nuclei in the periinfarct area as previously described (Kusano et al., 2007).
Statistical analysis
The mean and standard error (SE) were determined for replicate samples. An unpaired Student t test was applied to calculate the statistical significance between the means of two groups. Analysis of variance was performed to calculate the statistical significance for multiple samples. A p value less than 0.05 was considered significant.
Results
AAV-mediated transduction of EPCs augmented with genistein
We first examined the efficiency of transduction of peripheral blood-derived EPCs, using an AAV vector system (Kawamoto et al., 2001, 2003; Iwakura et al., 2005). Without pretreatment, the transduction efficiency was 23 ± 1.2% (see Supplementary Fig. 1A at
IGF-1-transduced EPCs inhibit apoptosis in and augment proliferation of neonatal cardiac myocytes
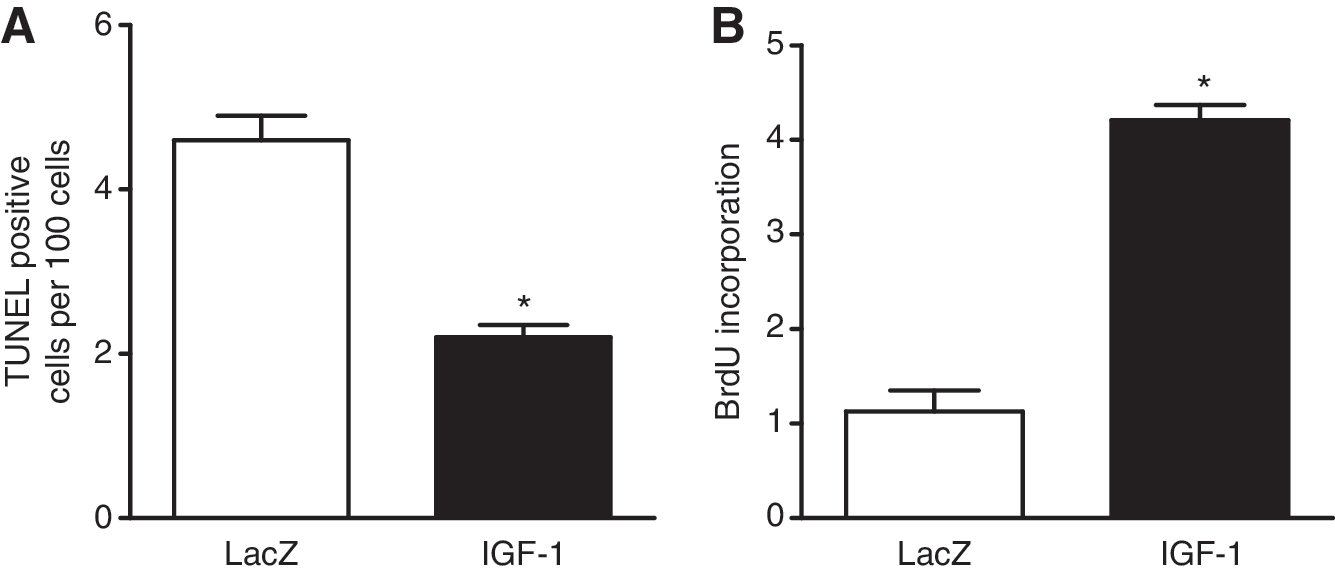
Reports demonstrate that IGF-1 has potent antiapoptotic and cell proliferation effects (Laviola et al., 2007). To determine whether IGF-1-expressing EPCs could directly exert this antiapoptotic effect, we cocultured neonatal rat cardiomyocytes with AAV-transduced EPCs (Yoon et al., 2005), and induced apoptosis by exposure to 100 mM hydrogen peroxide (H2O2) (Aikawa et al., 2000). In the presence of IGF-1-expressing EPCs, the number of H2O2 induced TUNEL-positive cardiomyocytes was significantly reduced compared with the control group (2.1 ± 0.2 vs. 4.6 ± 0.4; Fig. 1A). Next, to determine whether IGF-1-expressing EPCs could directly exert a cell proliferation effect, we measured the incorporation of BrdU, using the same coculture system (Yoon et al., 2005). When we cocultured neonatal cardiomyocytes with IGF-1-EPCs, the BrdU uptake of cardiomyocytes was significantly increased compared with the control group (4.1 ± 0.3 vs. 1.1 ± 0.1; Fig. 1B). Taken together, overexpression of IGF-1 in EPCs inhibited cardiomyocyte apoptosis induced by oxidant stress and augmented cardiomyocyte proliferation.
IGF-1-overexpressing EPCs improve cardiac function post-MI
For autologous EPC transplantation, we separately cultured autologous rat EPCs according to our published protocol (Kawamoto et al., 2001, 2003). After being cultured for 1 week, EPCs were transduced with AAV-lacZ control vector or AAV-IGF-1. We ligated the left anterior descending coronary artery of rats to induce myocardial infarction (MI), and transduced EPCs (1 × 104 total) were delivered by direct injection to five different portions of the periinfarct area (Kusano et al., 2007). The physiological effects of IGF-1-expressing EPCs after MI in rats were assessed by transthoracic echocardiography. At baseline before MI, left ventricular diastolic dimension (LVDd), left ventricular systolic dimension (LVDs), and fractional shortening (FS) were similar between the two groups (Fig. 2). As expected, LVDd and LVDs were substantially increased and FS was decreased 12 weeks after MI (Fig. 2). However, LVDd and LVDs were significantly lower in the IGF-1-EPC treatment group (0.89 ± 0.04 and 0.71 ± 0.03 cm, respectively) compared with the lacZ-EPC control group (0.99 ± 0.03 and 0.78 ± 0.06 cm, respectively) (Fig. 2). FS of the IGF-1-EPC group by 12 weeks (25.7 ± 0.2%) was significantly higher compared with the control group (18.2 ± 0.7%) (Fig. 2). These results indicate that IGF-1-expressing EPCs improved cardiac function and remodeling post-MI in rats.

Serial changes in echocardiographic parameters such as left ventricular diastolic dimension (LVDd), left ventricular systolic dimension (LVDs), and fractional shortening (FS) from baseline (day 0) to 12 weeks. Lines with blue triangles indicate rat heart transplanted with AAV-IGF-1-EPC, and lines with red squares indicate rat heart transplanted with control AAV-lacZ-EPCs (n = 7). *p < 0.05 versus control lacZ-EPC group.
Angiogenesis is induced by transplantation of IGF-1-overexpressing EPCs
Twelve weeks after cell transplantation, to confirm the in vivo induction of IGF-1 expression by AAV in an infarcted heart, we measured the concentration of human IGF-1 (hIGF-1) in the serum of rats by hIGF-1 ELISA (Dobrucki et al., 2010). Although there were no elevations of hIGF-1 in the lacZ-EPC control group, circulating hIGF-1 levels were significantly increased at both 1 and 3 months after transplantation of IGF-1-EPCs (Fig. 3A). These results suggest that the AAV-IGF-1 vector significantly transduced EPCs and subsequently improved expression of IGF-1 in the transplanted EPCs of the infarcted heart, and that transduced EPCs were an effective tool in delivery of gene to the targeted site.
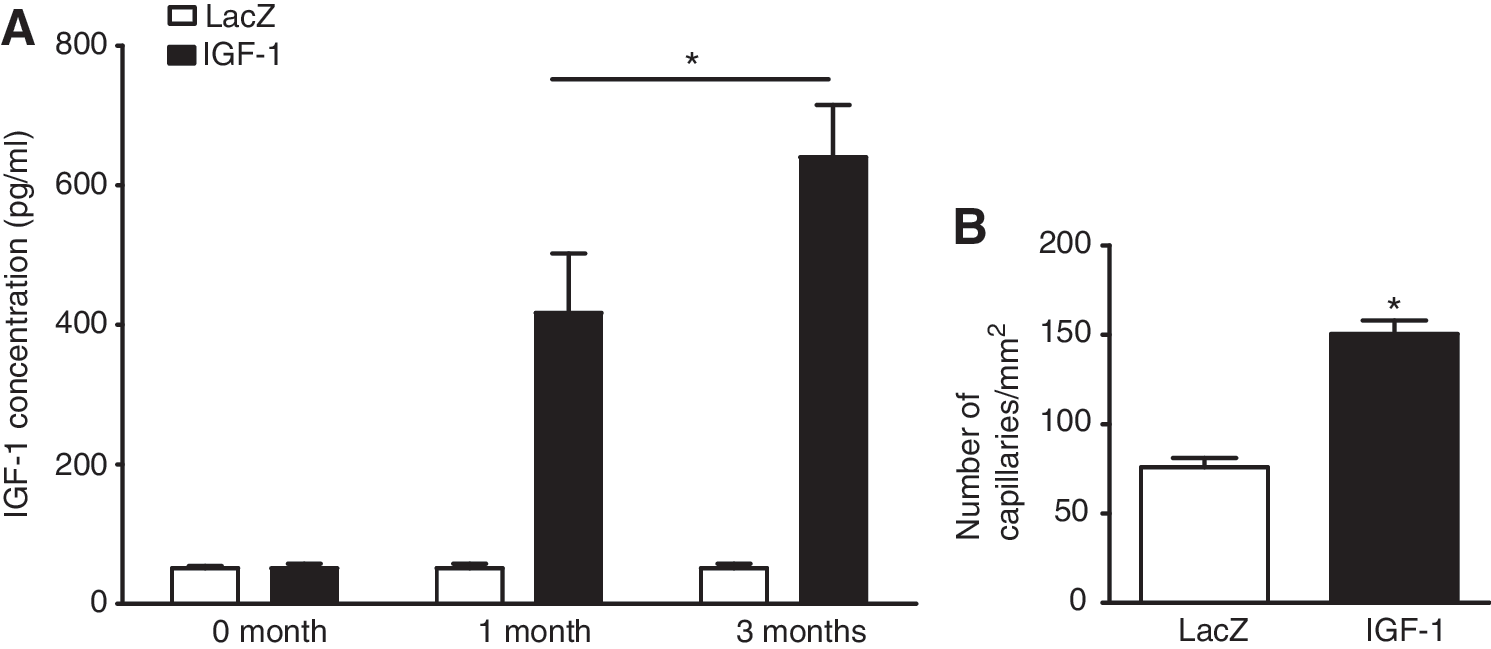
Reports have further indicated that IGF-1 has a strong effect on angiogenesis (Su et al., 2003; Lopez-Lopez et al., 2004; Rabinovsky and Draghia-Akli, 2004). Su and colleagues demonstrated that exposure to IGF-1 peptides potently induced endothelial cell migration in a Matrigel assay and capillary formation in an in vitro culture assay (Su et al., 2003), which are known to be important components of the proangiogenic response. Therefore, we examined angiogenesis between the lacZ-EPC group and the IGF-EPC group 12 weeks after MI. Immunohistochemical analysis revealed that capillary density of the heart in the IGF-1-EPC group was significantly higher than in the lacZ control group (151 ± 9 vs. 75 ± 5, respectively; Fig. 3B).
IGF-1-EPCs suppress cardiac apoptosis and induce cardiac proliferation post-MI
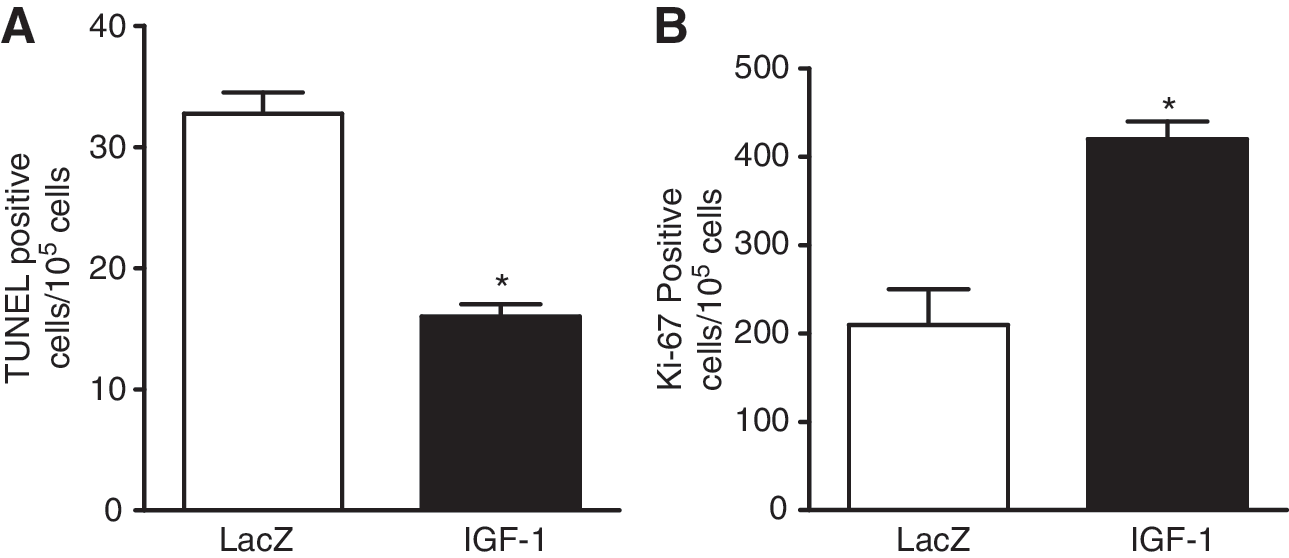
It has been reported that overexpression of IGF-1 attenuates the activation of myocyte necrosis and apoptosis after infarction in the transgenic mouse model (Li et al., 1997) and rescues cardiac myocytes from apoptosis in dilated cardiomyopathy (Welch et al., 2002). Accordingly, we compared cardiomyocyte apoptosis, by TUNEL assay analysis, between lacZ-EPC- and IGF-1-EPC-transplanted heart muscle post-MI. Twelve weeks after infarction, we harvested the tissue and performed α-actinin, TUNEL, and DAPI staining for cardiomyocytes, DNA fragments, and nuclei, respectively. The number of TUNEL-positive cells was significantly decreased in the IGF-1-EPC-treated group compared with the control lacZ-EPC group (16 ± 0.7 vs. 33 ± 1.2, respectively; Fig. 4A), indicating that AAV-IGF-1-EPCs may contribute to protect cardiac myocytes post-MI. It has also been reported that overexpression of IGF-1 increases the activation of cardiomyocyte proliferation after infarction in the transgenic mouse model (Li et al., 1997) and the number of proliferating myocytes in dilated cardiomyopathy (Welch et al., 2002). We reported that AAV-mediated growth hormone (GH) expression enhances cardiomyocyte proliferation post-MI in rats, as indicated by staining for the Ki-67 cell cycle marker protein (Kusano et al., 2007). In comparison with the lacZ control group, the number of Ki-67-positive myocytes was increased in the IGF-1 EPC group about 2.0-fold compared with the control group (420 ± 25 vs. 210 ± 55, respectively; Fig. 4B). These results, in combination with those of the in vitro assay (see Supplementary Fig. 1B), indicate that IGF-1-EPCs may contribute to promote cardiomyocyte proliferation post-MI.
Genetically modified EPCs are limited to the targeted tissue
To verify gene transfer and to determine the distribution of gene marking, total DNA was isolated from major organs in recipient rats. The presence of AAV sequences was analyzed by PCR as described previously (Iwakura et al., 2005). Agarose gel electrophoresis demonstrated a 286-bp band, derived from the inverted terminal repeat (ITR) sequence of the AAV genome and extending into the CMV promoter sequence (Iwakura et al., 2005), in the heart 12 weeks after transplantation (see Supplementary Fig. 2 at
Discussion
Although AAV2 is capable of transducing a variety of cell types (Van Vliet et al., 2008), endothelial cells (ECs) have been reported to be relatively resistant (Pajusola et al., 2002). We confirmed a relatively low transduction efficiency of EPCs despite the use of high-titer AAV vectors (see Supplementary Fig. 1A), which suggests that EPCs behave similarly to ECs. Accordingly, we sought to augment AAV transduction by using chemical reagents. In infected cells, the incoming AAV single-stranded DNA (ssDNA) must be converted into double-stranded DNA (dsDNA) to serve as a transcriptional template (Vincent-Lacaze et al., 1999; Nakai et al., 2000). Studies indicate that cellular permissiveness correlates with the phosphorylation status of FK506-binding protein-2 (FKBP2), which specifically interacts with the D sequence and inhibits AAV second-strand DNA synthesis (Qing et al., 2001; Zhong et al., 2004). Both genistein and hydroxyurea have been reported to promote dsDNA formation through inhibition of FKBP2 (Qing et al., 2001; Zhong et al., 2004). Moreover, a proteasome inhibitor, LLnL, has been shown to increase AAV transduction efficiency by protecting infected AAV vectors against degradation (Ding et al., 2003). To retain adequate EPC viability, we first used the MTT assay to determine the optimal pretreatment concentrations, which were as follows: 10 mM for hydroxyurea, 200 mM for LLnL, and 100 μM for genistein (see Supplementary Fig. 1B). At the optimal concentration, genistein yielded the greatest improvement in AAV transduction (see Supplementary Fig. 1C and D). In addition, another AAV serotype, AAV1 (Van Vliet et al., 2008), showed definite improvement after EPCs were pretreated with genistein, as transduction was lower than that of AAV serotype 2 (data not shown). Other reagents such as ethanol (Wheeler et al., 2000) and histone deacetylase inhibitors (Okada et al., 2006) may also augment AAV transduction efficacy. We have previously reported that transplantation of 1 × 105 EPCs improved cardiac function post-MI in rats (Kawamoto et al., 2001). In preliminary experiments, we observed that 1 × 104 EPCs transplanted into infarcted rat heart resulted in minimal differences between the control group and the treated group (data not shown). Accordingly, we genetically modified EPCs with AAV2 and transplantation of IGF-1-expressing EPCs (total, 1 × 104) led to significant improvement of cardiac function in the same rat infarction model (Fig. 2). In addition, we confirmed the persistence of AAV gene expression by detecting IGF-1 expression in the heart 12 weeks post-MI (see Supplementary Fig. 2).
Transplantation of EPCs induces angiogenesis in infarcted heart after autocrine and paracrine activation of angiogenic factors such as vascular endothelial growth factor (VEGF) and IGF-1 (Kawamoto et al., 2001, 2003). In this study AAV-IGF-1-transduced EPCs induced higher capillary density in the heart compared with control AAV-lacZ-EPCs (Fig. 3B), and thus secreted IGF-1 may provide an additional improvement beyond EPCs alone. In our in vitro study, using a coculture system, IGF-1-expressing EPCs decreased apoptosis and increased cell proliferation in neonatal rat cardiomyocytes (Fig. 1A and B). These results also support the salutary effect of IGF-1. In addition, transplantation of AAV-IGF-1-EPCs reduced cardiomyocyte apoptosis (Fig. 4A) and promoted cardiomyocyte proliferation (Fig. 4B) compared with control lacZ-EPCs post-MI, presumably via a paracrine mechanism. This approach of using a combination of cell and gene therapy can achieve better results than individual therapy because the two modes of therapy will support each other. In addition, assimilation of EPCs into the vasculature at the site of treatment could lead to long-term IGF-1 production, long after the time of delivery and beyond the effects conferred by the paracrine properties of EPCs. Moreover, because Thum and colleagues demonstrated that age-dependent impairment of EPCs is improved by growth hormone (Thum et al., 2007), it is possible that IGF-1 may act similarly on functionally impaired EPCs in elderly patients with MI.
The AAV vector system is safe and continues to offer hope for human gene therapy, but its promiscuous tropism may lead to undesirable tissue expression. Both cardiac-specific promoters and cardiac-selective serotypes are being explored to avoid this potential problem (Aikawa et al., 2002; Inagaki et al., 2006). In our experiments various rat organs were examined by PCR and no AAV genomes were detected other than in heart tissue injected with lacZ-EPCs (see Supplementary Fig. 2). Thus, it is possible to restrict AAV-mediated gene expression in vivo by this method of AAV-transduced cell transplantation, in contrast to direct in vivo injection of AAV vectors. Human clinical trials of progenitor cells continue to show promise to improve cardiac function (Losordo et al., 2007). Our study provides evidence that both stem cells and progenitor cells may act as effective tools for gene delivery by AAV, and that expression of IGF-1 may improve the efficacy of this transplantation approach.
Footnotes
Acknowledgments
This work was supported by the Uehara Memorial Foundation (R.A.) and by a Starr Foundation grant (R.A.). Additional support was provided by the Leukaemia Foundation of Australia, the National Health and Medical Research Council of Australia, an anonymous Foundation, and by Cure the Future (J.E.J.R.).
Author Disclosure Statement
None declared.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.