
Abstract
Lentiviral vectors (LV) can induce type I interferon (IFN I) production from murine plasmacytoid dendritic cells (pDC), but not myeloid (my)DC. Here, we investigated whether this mechanism is conserved in human DC. MyDC and pDC were isolated from peripheral blood and transduced with increasing vector concentrations. Compared with in vitro differentiated monocyte-derived DC, the transduction efficiency of peripheral blood DC was low (ranging from <1% to 45%), with pDC showing the lowest susceptibility to LV transduction. Phenotype and function of myDC were not directly modified by LV transduction; by contrast, pDC produced significant levels of IFN-α and tumor necrosis factor-α. pDC activation was dependent on functional vector particles and was mediated by Toll-like receptor 7/9 triggering. Coculture of myDC with pDC in the presence of LV resulted in myDC activation, with CD86 up-regulation and interleukin-6 secretion. These findings demonstrate that the induction of transgene-specific immunity is triggered by an innate immune response with pDC activation and consequent myDC maturation, a response that closely resembles the one induced by functional viruses. This information is important to design strategies aimed at using LV in humans for gene therapy, where adverse immune responses must be avoided, or for cancer immunotherapy, where inducing immunity is the goal.
Introduction
HIV-1-derived lentiviral vectors (LV) (Naldini et al., 1996) represent an attractive strategy for therapeutic gene transfer (Vigna and Naldini, 2000; Follenzi et al., 2002, 2004; Kang et al., 2005), as well as antitumor vaccination (Lizee et al., 2004; Dullaers et al., 2006; Lopes et al., 2006; Rowe et al., 2006; Breckpot et al., 2008; Arce et al., 2009). Crucial to the success of LV for either gene therapy or vaccination is their impact on the immune system. However, the effects of LV on the human innate and adaptive immune system have not been extensively investigated. In studies undertaken with other gene transfer systems, including adenoviral vectors and naked DNA, the delivery vehicle was shown to act as adjuvant for induction of APC maturation and subsequent antigen-specific T-cell priming (Meng and Butterfield, 2005), whereas conflicting results regarding the effect of LV on human APC have been reported (Gruber et al., 2000; Schroers et al., 2000; Dyall et al., 2001; Rouas et al., 2002; Chen et al., 2004; Lizee et al., 2004; Tan et al., 2005; Veron et al., 2006; Breckpot et al., 2007).
LV-mediated gene transfer can provide long-term expression of the transgene (Tg) in immunodeficient animals and humans. However, systemic administration of LV encoding the strong neo-antigen green fluorescent protein (GFP) to immunocompetent mice resulted in the induction of an adaptive immune response characterized by infiltration of GFP-specific CD8+ cytotoxic T cells in liver and spleen, and production of Tg-specific antibodies, with clearance of the GFP-expressing cells (Follenzi et al., 2004). Systemic administration of LV to mice triggers a rapid and transient IFN I response, which is dependent on functional vector particles; similar to what happens during viral infections, pDC were proposed to be involved in this response (Brown et al., 2007; Pichlmair et al., 2007). On the other hand, murine studies have shown that cutaneous LV delivery promotes direct transfection of skin-derived myDC, resulting in prolonged antigen presentation and potent cytotoxic T-cell immunity (He et al., 2006); moreover, very recently murine myDC were shown to respond to LV transduction in vitro via TLR3 and TLR7 (Breckpot et al., 2010). Therefore, different DC subsets have been involved in triggering innate and adaptive immune responses after LV administration, leading to the clearance of Tg-expressing cells and failure of gene transfer strategies.
The present work aimed to better characterize the direct effect of LV on the two major subsets of human DC. The response of in vitro differentiated monocyte-derived DC (moDC) and of ex vivo isolated myDC and pDC to LV transduction was investigated. Results show that moDC and myDC are not directly activated by LV, whereas pDC release significant amounts of IFN I and tumor necrosis factor (TNF)-α upon LV exposure and, in turn, activate myDC.
Materials and Methods
Cell preparation
Peripheral blood mononuclear cells (PBMC) from healthy donors were isolated by centrifugation over Ficoll-Hypaque gradients (Nycomed, Amersham). Approval for these studies was obtained from the San Raffaele Scientific Institute Institutional Review Board, and informed consent was provided for the use of blood samples according to the Declaration of Helsinki.
Monocyte-derived DC
CD14+ monocytes were isolated as the adherent fraction of PBMC after incubation for 1 hr in RPMI 1640 (BioWhittaker, Inc., Walkersville, MD) supplemented with 10% fetal calf serum, 100 U/ml penicillin-streptomycin (Bristol-Myers Squibb, New York, NY), and 50 μM 2-mercaptoethanol (DC medium) at 37°C. After extensive washing, adherent monocytes were differentiated into moDC by culture in 10 ng/ml recombinant human IL-4 (R&D Systems, Minneapolis, MN) and 100 ng/ml recombinant granulocyte macrophage colony-stimulating factor (GM-CSF) (Schering-Plough, Kenilworth, NJ) in DC medium. After 5 days, moDC were left unstimulated or were activated by 1 μg/ml lipopolysaccharide (LPS) (Sigma, St. Louis, MO). In some experiments, moDC were transduced at the indicated time point, concentration, and type of VSV-G.LV. Endotoxin levels in all vector preparations were evaluated and found to be below the minimal detection level (Pyrogent, BioWhittaker, Inc.). Purity and maturation state of moDC were checked by flow cytometric analysis to determine the expression of CD1a, CD14, CD83, HLA-DR, CD80, CD86, CD11c, and CD123 using directly coupled monoclonal antibodies (mAbs) (BD Biosciences, Franklin Lakes, NJ). Cell supernatant was harvested at the indicated time points, and IL-6, IL-10, IL-12, and TNF-α levels were determined by capture enzyme-linked immunosorbent assay (ELISA) according to the manufacturer's instructions (BD Biosciences). Detection limits were 5–15 pg/ml. The concentration of IFN-α was determined by an ELISA kit from Bender Medical Systems (Vienna, Austria; BMS216MST), according to the manufacturer's instructions. For mixed lymphocyte reaction response, allogeneic responder naive CD4+CD45RO− T cells (purified by MACS magnetic microbead separation) were stimulated by irradiated DC for 72 hr. [3H]Thymidine was added during the last 18 hr.
Peripheral blood DC purification and culture
Peripheral blood myDC and pDC were magnetically sorted with the CD1c (BDCA-1)+ dendritic cell isolation kit (Miltenyi Biotec, Bergisch Gladbach, Germany) and the negative pDC enrichment kit (StemCell Technologies, Vancouver, BC, Canada), respectively, following the manufacturers' instructions, to a purity of >95% in both cases. Purity and maturation state of blood DC were checked by flow cytometric analysis using directly coupled mAbs against CD1c, CD83, HLA-DR, CD80, CD86, CD11c, CD4, CD45RA, and CD123 from BD Biosciences. CD303 was from Miltenyi Biotec. Blood myDC and pDC (105 cells/well) were cultured in 96-well plates in 100 μl of the above-mentioned DC medium, containing 10 ng/ml recombinant human GM-CSF (Schering-Plough) or 20 ng/ml recombinant human IL-3 (R&D Systems), respectively. In some experiments, myDC were stimulated with 1 μg/ml LPS (Sigma-Aldrich) and pDC were stimulated with 10 μg/ml CpG-A oligonucleotide 2216 (Primm Srl, Milan, Italy). MyDC and/or pDC were transduced at the indicated concentration with the indicated type of LV encoding GFP in the presence or absence of an oligonucleotide inhibiting TLR7/9 activation (IRS954) at 10 μg/ml (Barrat et al., 2005). For coculture experiments, pDC and myDC were cultured (105 cells/well; 1:1 ratio; 20 ng/ml IL-3) in 96-well plates in the presence or absence of VSV-G.LV.
Vector production
Third-generation pCCLsin.PPT.hPGK.GFP.pre LV were produced by Ca3PO4 cotransfection of four plasmids into 293T cells. Supernatants were collected, passed through a 0.22-μm filter, and purified by ultracentrifugation as described (De Palma and Naldini, 2002). Titer was estimated on HeLa cells, and vector particles were measured by HIV-1 Gag p24 antigen immunocapture (NEN Life Science Products, Boston, MA). Vector infectivity was calculated as the ratio between titer and particle. For concentrated vesicular stomatitis virus (VSV) pseudotyped vector (VSV-G.LV), titer ranged from 6 × 109 to 8 × 109 transducing units (TU)/ml, and the concentration of physical particles ranged from 150 to 170 μg/ml HIV Gag p24 equivalents. For concentrated GP64/Ampho pseudotyped vector (GP64.LV or Ampho.LV), the titer was 109 TU/ml, and the particles were 100 μg/ml p24. Bald.LV was produced as described, but without envelope-expressing plasmid. VSV-G.LV-empty was produced as described, but without transfer vector. VSV-G.LV was used at concentrations ranging from 106 to 108 TU/ml, corresponding to a multiplicity of infection (MOI) of 2.5 to 250. The concentration of the other LV was matched by p24. In some experiments, VSV-G.LV was heat-inactivated (VSV-G.LV-HI) for 40 min at 65°C.
Statistical analysis
All statistical analyses for significant differences were performed with the nonparametric Mann-Whitney test. P values less than 0.05 were considered significant. Plots represent mean ± SEM.
Results
LV do not modify phenotype and function of moDC
We investigated the ability of LV encoding for GFP under the control of the ubiquitous phosphoglycerate kinase promoter and pseudotyped with the envelope glycoprotein from vesicular stomatitis virus (VSV-G.LV) to transduce moDC. Human moDC were differentiated by culturing adherent monocytes, isolated from peripheral blood of healthy donors, with IL-4 and GM-CSF. The cells were transduced at different days in culture (day 0 to day 3) during moDC differentiation and analyzed by flow cytometry on day 7. VSV-G.LV efficiently transduced DC precursors from day 0 to day 3 of culture, without significant differences (20–50% at MOI of 2.5, 60–80% at MOI of 25) (data not shown and Fig. 1a, respectively). Thus, in subsequent experiments, moDC were transduced on day 2. DC differentiated from transduced precursors maintained their immature phenotype when cultured in medium with cytokines alone (Fig. 1b), and when activated with LPS matured to produce pro-inflammatory cytokines (data not shown) and to up-regulate HLA-DR, CD83, and CD86, as untransduced mDC (Fig. 1b).
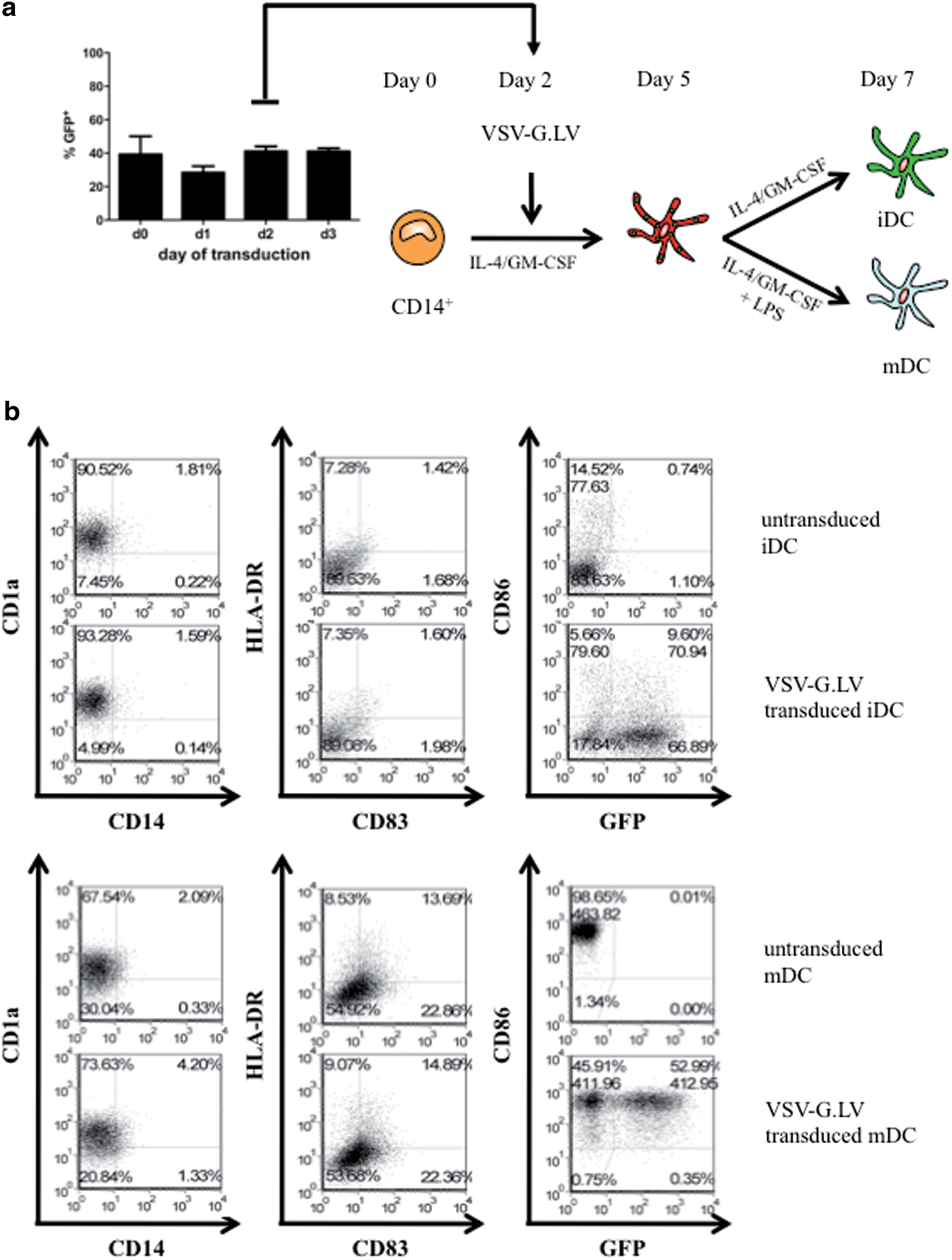
High transduction efficiency and no VSV-G.LV-mediated activation of transduced human moDC. Adherent monocytes were differentiated into moDC in the presence of GM-CSF and IL-4. (
VSV-G.LV (MOI of 25) poorly transduced DC when added during DC differentiation on day 5 of culture (1–5%; Fig. 2a), indicating that differentiated moDC are less susceptible to LV transduction compared with monocyte precursors. The low transduction efficiency was not due to activation of moDC by LV, because moDC exposed to LV on day 5 did not up-regulate CD83, HLA-DR, and CD86 (Fig. 2a), did not secrete the pro-inflammatory cytokines IL-6, IL-12, TNF-α, and IFN-α (Fig. 2b and data not shown), and did not acquire high allo-stimulatory capacity against naive CD4+ T cells (Fig. 2c), in contrast to LPS-activated moDC. Notably, moDC analyzed for the expression of CD86, CD83, and HLA-DR at early time points (5 and 10 hr) after VSV-G.LV exposure did not show any sign of activation (data not shown and Supplementary Fig. S1; Supplementary Data are available at
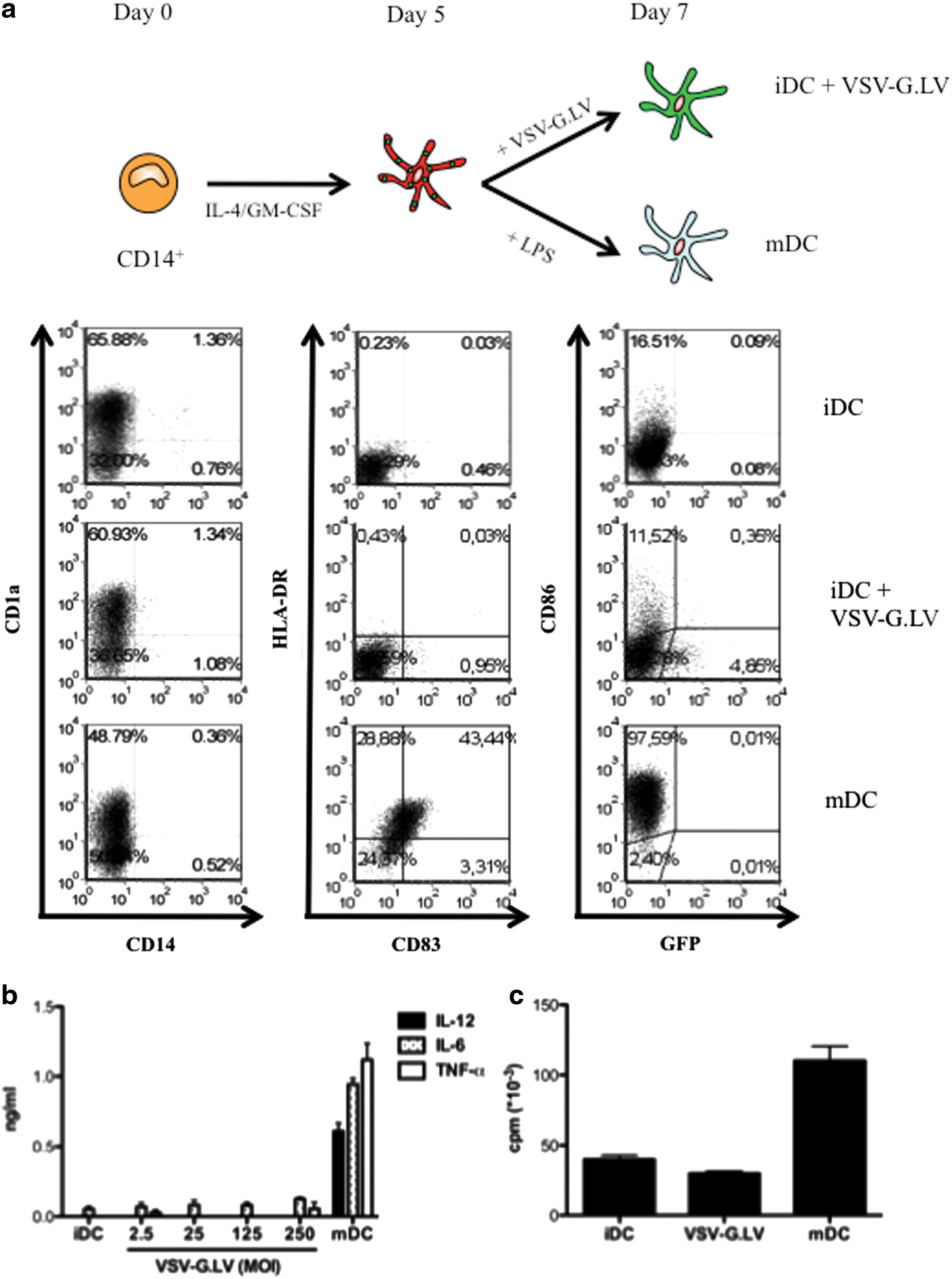
VSV-G.LV does not transduce or activate differentiated moDC. Adherent monocytes were differentiated into DC in the presence of GM-CSF and IL-4. (
These results demonstrate that monocytes display higher susceptibility to LV transduction compared with in vitro differentiated immature moDC. Moreover, LV transduction does not directly activate moDC and does not alter their ability to respond to the TLR4 ligand LPS.
LV do not activate peripheral blood myDC but promote IFN-α secretion by pDC
We then investigated the effect of LV on freshly isolated human peripheral blood DC subsets. Bead-purified myDC and pDC were cultured in the presence of VSV-G.LV for 2 days. At MOI of 25, myDC were transduced at lower levels compared with moDC (on average, 24.4% of GFP+ cells, range 11–47%), and pDC were barely transduced (8.3% GFP+ cells, range 0.2–16.5%; p = 0.0037 between pDC and myDC; Fig. 3a). At higher concentrations, VSV-G.LV transduced both DC subsets, although transduction efficiency remained significantly higher in myDC (76%, range 66–88%, at MOI of 250) compared with pDC (33%, range 4.7–58%; p = 0.0043 for MOI of both 125 and 250; Fig. 3b). VSV-G.LV did not induce pDC activation in terms of CD80, CD83, and CD86 up-regulation at any dose tested (Fig. 3a and data not shown). Similarly, VSV-G.LV did not induce up-regulation of CD80 and CD83 on myDC, but induced only marginal up-regulation of CD86; however, the latter effect was not consistent among different donors and LV doses (Fig. 3a and data not shown). Similar to moDC, peripheral blood myDC did not secrete IFN-α, IL-6, and TNF-α in response to VSV-G.LV, indicating that the mild up-regulation of CD86 observed likely resulted from a nonspecific DC perturbation (Fig. 3c). Conversely, upon incubation with VSV-G.LV, pDC secreted significant amounts of IFN-α (on average, 19 ng/ml; p = 0.0003 between transduced pDC and myDC; p < 0.0001 between untransduced and tranduced pDC) and discrete levels of TNF-α (0.5 ng/ml; p = 0.0001 between transduced pDC and myDC; p = 0.0001 between untransduced and tranduced pDC), but not IL-6. IL-12 was not produced by any DC population under these culture conditions, but myDC produced up to 1.4 ng/ml IL-12 in response to the TLR8 ligand R848 (data not shown).
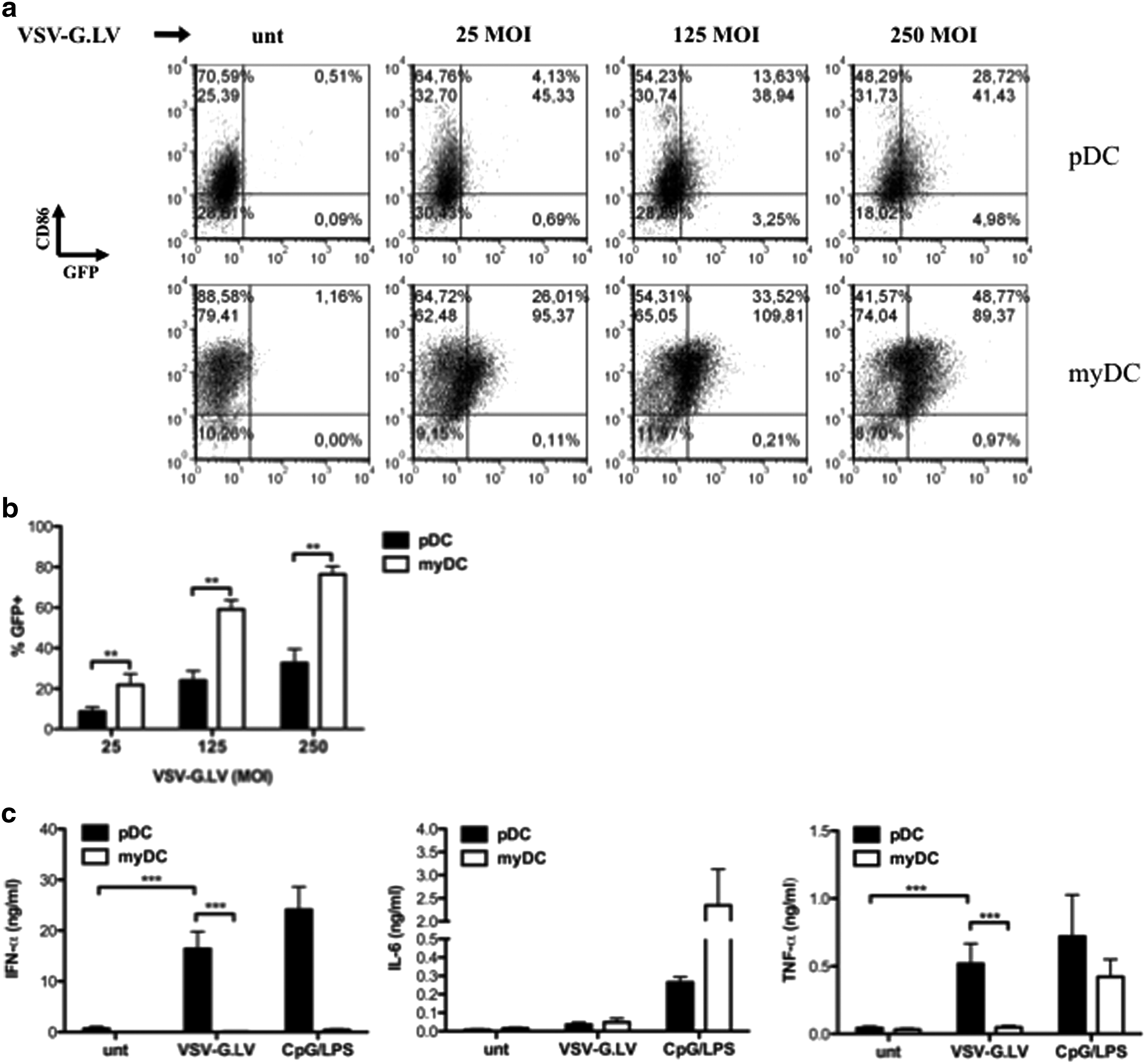
VSV-G.LV poorly transduces freshly isolated myDC and pDC and activates pDC, but not myDC, to produce IFN-α and TNF-α. pDC and myDC were purified from peripheral blood and cultured in the presence of the indicated stimuli for 48 hr. (
Based on these findings, we conclude that LV do not promote the maturation of myDC, but directly activate pDC to produce the antiviral cytokine IFN-α and the pro-inflammatory cytokine TNF-α.
The effect of LV on pDC requires viral entry
To define which LV components could directly activate pDC, the cells were cultured in the presence of modified VSV-G.LV preparations. None of the vectors tested, except for VSV-G.LV, were able to transduce pDC or to induce up-regulation of CD86 or CD80 (Fig. 4a and data not shown). Bald.LV (without envelope) did not trigger IFN-α production by pDC (Fig. 4b), suggesting that either the envelope itself or viral entry is required for pDC activation. To address this issue, we cultured pDC in the presence of an LV preparation devoid of viral genetic material, but pseudotyped with VSV-G (VSV-G.LV-empty). As expected, we could not find any GFP+ cells, but pDC were still able to produce IFN-α (on average, 7.5 ng/ml vs. 12 ng/ml induced by VSV-G.LV). The activation was specifically due to the use of the VSV-G envelope, because pDC did not respond to LV carrying genetic material but pseudotyped with different envelopes (Ampho.LV or GP64.LV; Fig. 4b and data not shown). Moreover, heat-inactivated LV (VSV-G.LV-HI), in which both genetic material and viral proteins are inactivated, was ineffective in pDC transduction and did not stimulate IFN-α production. These results suggest that the stimulatory effect of VSV-G.LV on pDC requires viral entry through the endosomal route and is enhanced in the presence of a functional VSV-G envelope. However, VSV-G.LV-empty induced lower levels of IFN-α secretion by pDC compared with VSV-G.LV in all donors tested, suggesting that pDC activation may require also recognition of the viral RNA packaged in the viral particle.
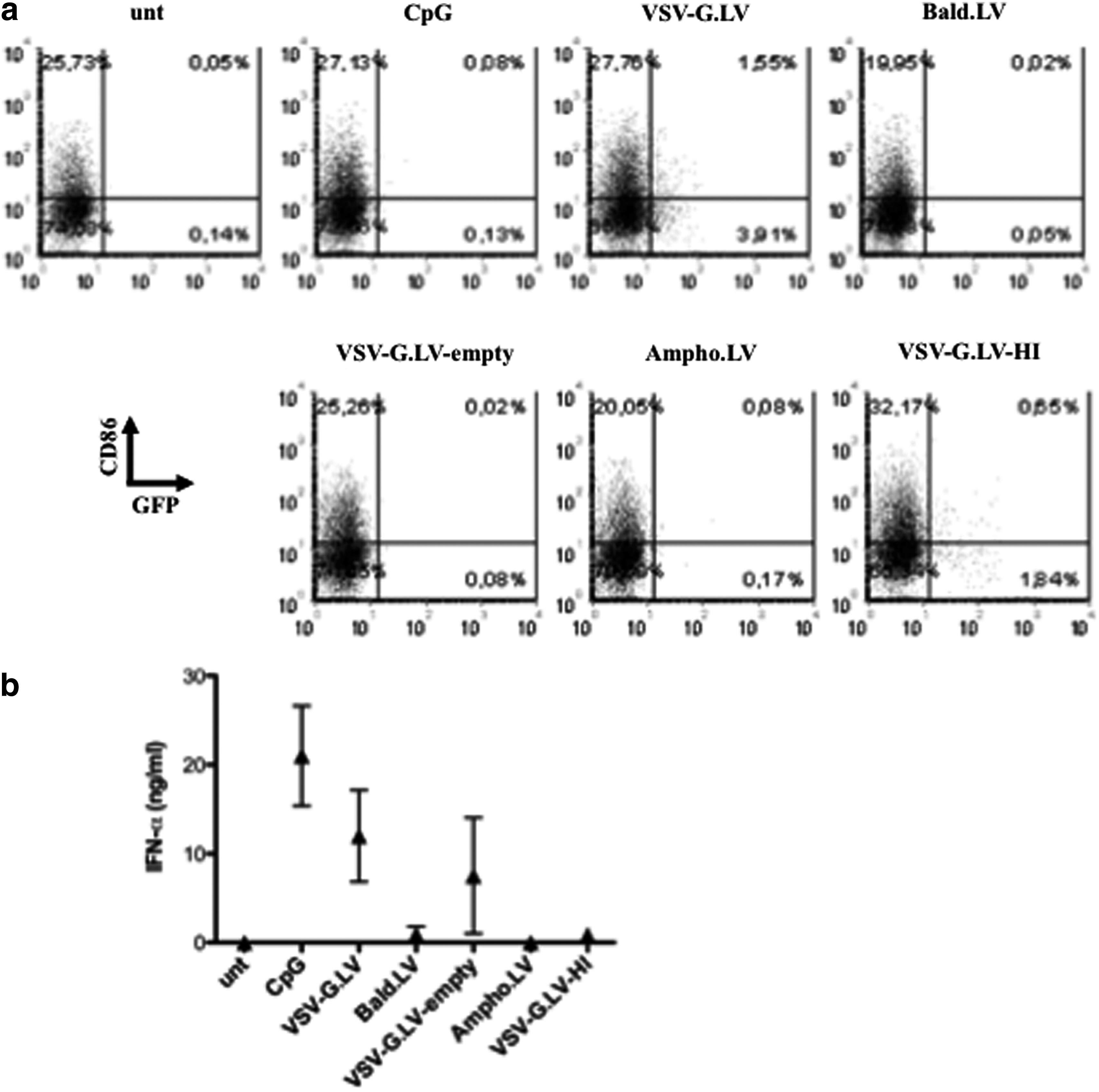
VSV-G.LV-mediated activation requires cell penetration by LV. pDC were purified from peripheral blood by magnetic beads and cultured in the presence of the indicated LV, all encoding GFP (MOI of 25). Cells were analyzed by flow cytometry (
Abrogation of IFN-α secretion by specific TLR7/9 blockade on pDC
It has been shown that the triggering of TLR7 and TLR9 by viral genetic material activates IFN-α production by pDC (Beignon et al., 2005; Uematsu et al., 2005). We therefore tested whether HIV-derived LV activate pDC via TLR7/9. IRS954 (a generous gift from Dr. Franck Barrat) is an oligodeoxynucleotide containing inhibitory sequences, which has been shown to inhibit TLR7/9-mediated activation in human cells (Barrat et al., 2005). As expected, IRS954 strongly reduced CpG-induced IFN-α production by pDC (30 vs. 14 ng/ml, p = 0.0457). Interestingly, IRS954 significantly inhibited LV-mediated IFN-α production by pDC (21 vs. 3 ng/ml, p = 0.005; Fig. 5a), without altering their immune phenotype (data not shown). However, incubation with IRS954 had no effect on TNF-α release upon LV or CpG exposure, indicating that the release of these two cytokines can be differentially regulated (Fig. 5b). Interestingly, IRS954 did not down-regulate IFN-α production by pDC activated with VSV-G.LV-empty (Supplementary Fig. S2), which does not contain RNA, suggesting that the other major component of VSV-G.LV responsible for pDC activation, apart from viral RNA, is not DNA contaminant, a ligand for TLR9.
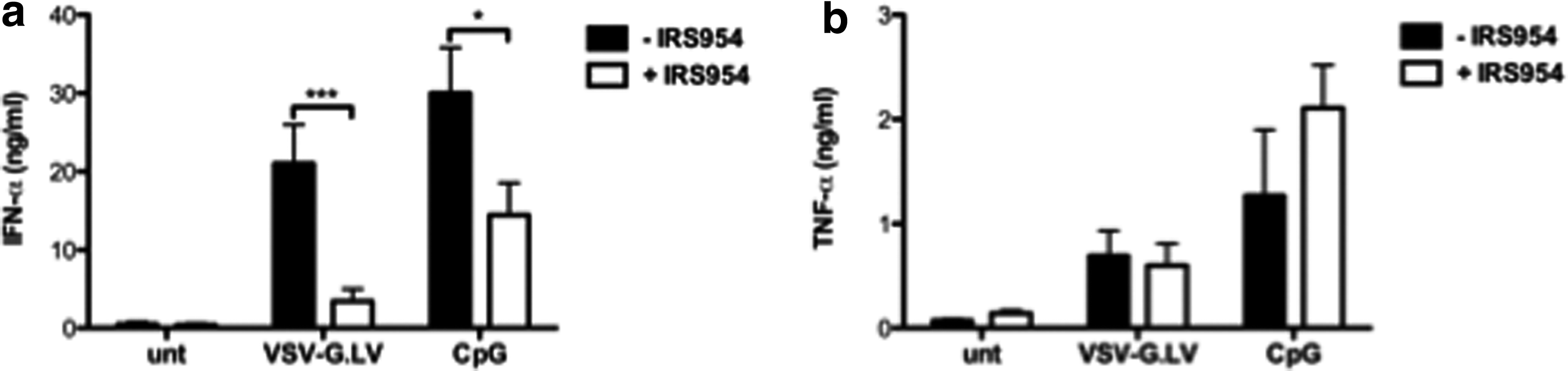
Abrogation of IFN-α secretion by specific TLR7/9 blockade on VSV-G.LV-activated pDC. pDC were purified from peripheral blood and cultured for 48 hr in the presence of the indicated agents. IFN-α (
These results demonstrate that LV induce IFN-α production by pDC through TLR7/9 engagement, and provide a possible strategy for modulating this response.
VSV-G.LV-activated pDC promote the bystander maturation of myDC
The demonstration of a direct activation of pDC, but not myDC, by VSV-G.LV supports the hypothesis that activation of the human innate immune response occurs following systemic LV administration (Brown et al., 2007). However, this does not necessarily mean that administration of LV will result in an immune response against the Tg-encoded product. Induction of antigen-specific adaptive immune responses would require Tg presentation by mature myDC. Fonteneau et al. (2004) have demonstrated that coculture of HIV-activated pDC with immature myDC results in myDC maturation, without a direct activation of the latter by HIV. To investigate whether VSV-G.LV-activated pDC may influence myDC in a similar way, we cocultured pDC and myDC in the presence of VSV-G.LV. MyDC cocultured with autologous pDC up-regulated CD86 only in the presence of VSV-G.LV (on average, 41% of CD86 up-regulation, calculated as percentage of CD86high within the CD11c+ cells; Fig. 6a). Furthermore, myDC did not secrete significant levels of IL-6 when cultured with either VSV-G.LV or pDC, whereas up to 700 pg/ml IL-6 was produced in cultures of myDC with pDC and VSV-G.LV (on average, 105 vs. 215 pg/ml between the cocultures in the absence or presence of VSV-G.LV, p = 0.0151; p = 0.0112 between myDC + VSV-G.LV in the absence or presence of pDC; Fig. 6b). Moreover, myDC cultured with supernatants obtained from VSV-G.LV-exposed pDC produced IL-6 (n = 4; data not shown), suggesting that one or more soluble factors produced by LV-activated pDC are responsible for the bystander activation of myDC. Notably, addition of neutralizing anti-TNF-α antibodies in the pDC–myDC cocultures during LV exposure reduced the release of IL-6 by myDC up to 60% (n = 2), whereas addition of IRS954, which blocks IFN-α release had no effect (n = 7; data not shown).

VSV-G.LV-activated pDC promote the bystander maturation of myDC. pDC and myDC were purified from peripheral blood of the same buffy coat and cocultured at a 1:1 ratio for 48 hr in the presence of VSV-G.LV at MOI of 25. Cells were analyzed by flow cytometry (
Discussion
In the present study, we characterized the in vitro response of different human DC subsets to LV. We evaluated transduction efficiency and activation of DC precursors, moDC, and peripheral blood myDC and pDC by LV. Our results show that monocytes display higher susceptibility to LV transduction compared with both in vitro differentiated moDC and peripheral blood DC. Upon LV exposure, moDC and freshly isolated myDC maintain their immature features, whereas pDC become activated and secrete IFN-α and TNF-α. LV-mediated pDC activation requires endosomal TLR7/9 triggering; moreover, LV-activated pDC promote the bystander maturation of myDC via soluble factors.
Determining the effect of LV on the human immune system is important for developing in vivo therapies with these vectors in humans. Gene replacement therapy for hemophilia B was limited by the induction of an immune response against components of the transfer vector, AAV (adeno-associated virus), ultimately resulting in elimination of the genetically modified cells (Manno et al., 2006; Nayak and Herzog, 2009). In contrast, attempts are being made to utilize the immune responses induced by viral and nonviral gene transfer systems to develop an effective method of vaccination for infectious diseases and cancer. Different factors can influence the triggering of an immune response, including the type of vector, the route of administration, and the dose of vector (Herzog et al., 1999; Mount et al., 2002; Brown et al., 2004). The innate immune system also plays a major role in determining the outcome of gene transfer (Brown and Lillicrap, 2002). This is highlighted by the unfortunate death of one individual who was administered an adenovirus for the treatment of ornithine transcarbamylase deficiency, in which the vector appears to have triggered a lethal inflammatory cascade. The type of innate response that occurs after vector delivery can also have an impact on the type of adaptive immune response that ensues.
LV are promising tools for gene therapy as well as genetic vaccination. However, little is known about how these vectors will impact the innate immune system in humans. Our group showed that, in mice, LV trigger a rapid and transient IFN I response similar to the one evoked by RNA viruses (Brown et al., 2007). Here, we demonstrate that HIV-derived LV trigger a similar IFN I response in human pDC, but not other human DC subsets (Liu, 2005). Upon LV exposure, pDC secrete high amounts of the antiviral cytokine IFN-α and significant levels of TNF-α; this activation is mediated by LV entry through the endosomal route, which allows TLR triggering by LV-derived nucleic acids. Indeed, LV not internalized by pDC, such as Bald.LV, cannot trigger their activation.
It is widely accepted that classical myDC promote the induction of antigen-specific adaptive T cell-mediated immunity through presentation of antigen in the context of pro-inflammatory membrane-bound and secreted molecules (Banchereau et al., 2000). In vitro, myDC are more susceptible than pDC to HIV infection and are not activated by the virus itself (Fonteneau et al., 2004; Smed-Sorensen et al., 2005). By contrast, HIV can activate pDC via TLR7-dependent recognition of its single-strand RNA genome; this activation results in the production of high levels of IFN-α that mediate activation of autologous bystander myDC (Fonteneau et al., 2004; Beignon et al., 2005). Similarly, myDC are not activated by LV transduction, but are more effectively transduced compared with pDC (Fig. 3). Moreover, pDC recognize LV via TLR7/9 interactions and are activated to produce IFN-α and TNF-α, finally inducing myDC maturation via soluble factors. These findings have several implications for the in vivo injection of LV in clinical settings of gene therapy or genetic vaccination. pDC initially mediate direct and bystander innate antiviral responses through the secretion of IFN I, but upon activation they can differentiate into mature DC, which directly regulate T-cell functions (Liu, 2005). pDC can also induce local activation of other APC, including myDC, which take up and process the Tg to be presented to T cells in lymphoid organs. Therefore, pDC could directly present the Tg to T cells or, more likely, promote the maturation of local myDC, which in turn would present the Tg to T cells in a pro-inflammatory context, inducing Tg-specific immunity. Our findings support the in vivo data from our earlier work, which suggested a role for murine pDC in the initiation of Tg-specific immunity after LV injection (Brown et al., 2007), but are in contrast with a very recent report showing that murine bone marrow-derived myDC recognize LV through TLR3/7 (Breckpot et al., 2010). This discrepancy can be ascribed to species-specific differences in the expression and function of TLR, or in intracellular routing of LV and their receptors. Considering the major differences between human and mouse DC, our work is of pivotal importance to understand the immunogenic potential of LV in humans.
Internalization of viral genetic material activates pDC through a TLR-mediated mechanism. It is likely that LV are recognized specifically by TLR7, as already reported for HIV, because their genetic material is constituted by single-strand RNA; however, as IRS954 blocks both TLR7 and TLR9 signaling, we cannot rule out whether TLR9 is also being activated by some DNA present in VSV-G.LV preparations as a result of initiation of retrotranscription already in virions or from plasmid contaminants (Pichlmair et al., 2007). However, addition of IRS954 during pDC exposure to LV-VSV.G-empty did not block IFN-α release, suggesting that DNA contaminants (ligands for TLR9) in our LV preparations do not represent the stimulus for pDC activation, in contrast to the results in other reports (Pichlmair et al., 2007). In addition, our data suggest that VSV-G is involved in pDC activation in vitro, supporting its role in the induction of LV immunity, as already reported in vivo (Marsac et al., 2002; Kuate et al., 2006). However, it remains to be determined whether pDC activation is mediated by direct recognition of VSV-G by an unknown receptor, or by VSV-G-mediated carryover of other undetermined TLR ligands from producer cells. It must be mentioned that GP64-pseudotyped LV induced IFN I responses in vivo (Brown et al., 2007), whereas in vitro exposure of human pDC to GP64.LV did not result in IFN-α production; it is possible that species-specific differences in viral entry and/or GP64-mediated endocytosis account for this discrepancy.
In our hands, LV induced high levels of IFN-α production, but not up-regulation of costimulatory molecules on pDC (Fig. 2); these data point against a direct role of pDC in the activation of Tg-specific adaptive immune responses, suggesting that they can act indirectly, through the activation of other cell subsets, such as myDC. This split response (IFN-α production vs. enhancement of antigen presentation) has been reported on pDC for TLR9 ligands, such as CpG-A and CpG-B, and it is caused by their differential molecular structure and subcellular localization (Guiducci et al., 2006). Moreover, upon recognition of viruses via TLR7, pDC produce IFN I in significant amounts, whereas at least some synthetic TLR7 ligands induce pDC maturation in terms of up-regulation of costimulatory molecules and production of pro-inflammatory cytokines, accompanied by low-level production of IFN-α. Recently, the described differential response of pDC to TLR7 ligands has been linked to a differential nuclear translocation of IFN regulatory factor-7 (IRF-7) and nuclear factor- κB (NF-κB) (Di Domizio et al., 2009), the two transcription factors responsible for the IFN-α release, and the production of pro-inflammatory cytokines and up-regulation of costimulatory molecules, respectively (Gilliet et al., 2008). We therefore hypothesize that LV-mediated pDC activation results in TLR7-driven nuclear translocation of IRF-7, but not NF-κB, mimicking the mechanism of pDC activation by viruses.
LV represent relevant candidates for therapeutic gene transfer in various indications, including both cancer immunotherapy and immunosuppression (Dullaers and Thielemans, 2006). The ability of LV to transduce moDC without affecting their phenotype and functions has important implications for strategies aimed at ex vivo DC gene transfer, which could be effective in the induction of both antigen-specific immunity and tolerance. LV have been used for gene delivery in different cell types, including human DC differentiated in vitro either from peripheral blood CD14+ monocytes or from CD34+ hematopoietic stem cells (Gruber et al., 2000; Schroers et al., 2000; Dyall et al., 2001; Salmon et al., 2001; Rouas et al., 2002; Breckpot et al., 2004; Chen et al., 2004; Lizee et al., 2004; Tan et al., 2005; Veron et al., 2006). Contrasting results on the efficacy of LV transduction on moDC and their functional response have been obtained. The transduction efficacy reported for HIV-based vectors on moDC varies from 10% to more than 80%. Moreover, some groups have reported LV-mediated activation of moDC, whereas others, including our group, did not detect changes in moDC phenotype and functions. These discrepancies can be ascribed to different factors, including the different LV structure, dose and mode of preparation, the time of transduction, the protocol used for moDC generation, and the use of agents such as Polybrene, protamine sulfate, or fibronectin to improve the transduction. DC activation can be useful when the aim is to induce antigen-specific immunity, such as in DC cell therapy for the treatment of cancer, but is equally detrimental when it leads to clearance of Tg-corrected cells.
Our data suggest that strategies aimed at using LV in vivo for long-term gene replacement therapy may need to suppress the initial IFN I response by pDC, possibly by the use of molecules such as IRS954. In addition, several autoimmune diseases, such as systemic lupus erythematosus, could take advantage of a therapeutic strategy in which IFN I production by pDC is blocked. On the other hand, enhancing the IFN I response may be beneficial to therapies aimed at using LV for cancer immunotherapy. The data reported here represent the first demonstration that LV can activate human pDC, the professional IFN I-producing cells specifically devoted to detect and counteract viral infections, clearly showing that LV retain “viral” features that can be detected by this specialized subset, but not by other APC, such as myDC and in vitro differentiated moDC. As moDC represent the easiest tool for ex vivo DC manipulation for different purposes, these findings also prove the safety of human moDC transduction when cell activation is not required and even detrimental, paving the way for approaches aimed at inducing Tg-specific tolerance by moDC transduction.
Footnotes
Acknowledgments
We would like to thank Dr. Franck Barrat (Dynavax Technologies Corporation) for his kind gift of the IRS954 oligonucleotide. This study was supported by a grant from Italian Telethon Foundation. E.H. was supported by a Human Frontier of Science Program long-term fellowship award.
Author disclosure statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.