
Abstract
Dendritic cells are the key antigen-presenting cells involved in the initiation of the adaptive immune response. Recombinant adeno-associated viruses (rAAVs) can transduce dendritic cells and have gained attention as potential vaccines capable of stimulating T cell immunity. Here we show that rAAV2 pseudotyped with type 6 capsid (rAAV2/6) exhibits significantly higher tropism for human monocyte-derived dendritic cells (MoDCs) than other serotypes and variants. Transduction was abolished by a single lysine-to-alanine mutation within the AAV6 capsid previously shown to inhibit binding to heparin. However, unlike rAAV2, soluble heparin did not inhibit rAAV2/6 transduction of MoDCs. Further enhancement of MoDC transduction was observed after mutation of Tyr-731 in the capsid of AAV6 consistent with a report that tyrosine residues are phosphorylated, leading to ubiquitination of capsids during uptake. Pseudotyped rAAV2/6 vectors containing a Y731F mutation minimally altered the immunophenotype of MoDCs, which retained their immunostimulatory ability and were able to stimulate an antigen-specific CD8+ T cell clone. These findings should assist in the development of rAAV2/6 as a vaccine vector.
Introduction
Adeno-associated virus (AAV), a nonpathogenic member of the family Parvoviridae, can deliver genes to DCs, resulting in the presentation of vector-encoded antigens to T cells. Studies have demonstrated that recombinant AAV (rAAV) can transduce MoDCs, which can stimulate antigen-specific T cells in vitro and in vivo (Xin et al., 2006; Veron et al., 2007). Stimulation of antibody and T cell responses after delivery of antigen by rAAV vectors has also been demonstrated in mice (Aldrich et al., 2006; Xin et al., 2006) and rhesus macaques (Johnson et al., 2005). Importantly, vectors based on AAV lack sequences encoding viral proteins, significantly reducing the risk of an immunodominant cell-mediated immune response directed against vector-derived epitopes (Smith et al., 2005). In addition, the genome of rAAV replicates extrachromosomally in an episomal form, reducing the risk of insertional mutagenesis compared with lentiviral vectors (Smith, 2008). The rAAV genome is conventionally packaged as a single-stranded DNA molecule with an optimal coding capacity of 4.1–4.9 kb, although up to 5.2 kb can be packaged with reduced efficiency (Dong et al., 1996).
At present 12 distinct AAV serotypes (AAV1–AAV12) and more than 100 variants have been isolated from human or nonhuman primate sources (Wu et al., 2006b; Schmidt et al., 2008). This variability has led to the development of pseudotyped rAAV vectors in which transgenes flanked by AAV2 inverted terminal repeats (ITRs), either as single-stranded or self-complementary molecules, are packaged into a diverse range of capsids. Although rAAV2 was the first serotype to be exploited as a gene vector and remains the most commonly used, the availability of diverse AAV serotypes exhibiting different cell tropism has allowed for a more rational approach to rAAV serotype selection for the delivery of genes to specific cell types and evasion of preexisiting neutralizing antibodies against the common human AAV2 (Wu et al., 2006b).
Successful transduction of human MoDCs with AAV was first demonstrated using rAAV2, establishing the potential for the use of rAAV in ex vivo immunotherapy (Ponnazhagan et al., 2001). More recent studies have confirmed the ability of AAV2 to stimulate antigen-specific CD8+ T cells after ex vivo transduction of human DCs and revealed that rAAV2/1 (i.e., rAAV2 pseudotyped with type 1 capsid) is more efficient than rAAV2 in transducing MoDCs (You et al., 2006; Veron et al., 2007). Although these studies confirm the ability of DCs to present AAV-encoded antigen to T cells and thus the potential utility of rAAV as a vaccine vector, the successful deployment of rAAV-based vaccines may be limited by the tropism of these serotypes for other tissue/cell types. A more systematic screen of AAV1–AAV5, AAV7, and AAV8 revealed an inherent tropism of rAAV5/5 for both human and murine DCs, leading to robust immune responses against human immunodeficiency virus in mice (Xin et al., 2006), whereas another study identified rAAV2/6 as having the highest tropism for murine bone marrow-derived DCs (Aldrich et al., 2006). Further refinement of the serotype specificity of rAAV toward human dendritic cell subtypes will likely have a significant impact in the choice of vectors used as vaccines.
In the present study we compared a wide range of pseudotyped rAAV vectors for their ability to transduce human MoDCs at various stages of differentiation. We report that a single exposure of monocytes or immature MoDCs to rAAV2/6 results in significantly greater transduction of DCs than with any other serotype. Further improvement in transduction efficiency was observed after mutation of tyrosine residues within the viral protein 1 (VP1) capsid. Transduction with rAAV2/6 caused minimal alteration of the immunophenotype of MoDCs, which retained their immunostimulatory ability. Furthermore, antigen encoded by rAAV2/6 was correctly processed and presented to T cells by transduced MoDCs. Altogether, these results provide a rational basis for the use of AAV2/6-based vectors in the development of human vaccines and immunotherapy applications.
Materials and Methods
Generation of human monocyte-derived dendritic cells
Human blood was obtained from volunteers under approval granted by the University of Auckland (Auckland, New Zealand) Human Ethics Committee. Peripheral blood mononuclear cells were isolated by centrifugation on a Lymphoprep (Axis-Shield, Dundee, Scotland) cushion in Leucosep tubes (Greiner Bio-One, Frickenhausen, Germany). Monocytes were isolated by negative selection, using either a MACS (magnetic activated cell sorting) monocyte isolation kit II (Miltenyi Biotec, Bergisch Gladbach, Germany) or Dynabeads Untouched human monocytes kit (Invitrogen, Auckland, New Zealand). Purity was assessed by flow cytometry and was consistently >85% (data not shown). Cells were seeded at 1 × 106 cells/ml and cultured in RPMI (Invitrogen) supplemented with glutamine (GlutaMAX-I supplement; Invitrogen), penicillin, and streptomycin (Invitrogen) (sRPMI), 10% fetal bovine serum (FBS; Invitrogen), and IL-4 (50 ng/ml) and GM-CSF (100 ng/ml) (both from Peprotech, Rocky Hill, NJ). Cells were cultured for 6 days, with half the medium replaced every 2 days. Phenotype was assessed on day 6 by assessing CD14, CD1a, HLA-DR, CD83, and CD86 expression by fluorescence-activated cell sorting (FACS) (data not shown and see the section Flow Cytometry).
Production and purification of rAAV
Each batch of rAAV was produced by transient triple transfection of five 15-cm plates of 293 cells with linear 25,000-Da polyethylenimine (Polysciences, Warrington, PA) (Reed et al., 2006). The transfected plasmids encoded the vector genome (pAM/CAG-eGFP-WPRE-bGHpA [Luk et al., 2003], kindly provided by D. Young, University of Auckland), the adenoviral helper functions (pFΔ6; Harrop et al., 2004), and the AAV Rep and Cap proteins (pH21, pNLrep, pH25A, pBR18-D2/8 [all kindly provided by D. Young], pAAV2/6, pAAV2/9, pAAV2/rh8c, pAAV2/rh10, or pAAV2/rh13R [all kindly provided by J. Wilson, University of Pennsylvania, Philadelphia, PA]). To make rAAV encoding the hepatitis B virus core antigen (HBcAg), HBcAg was cloned into pAM/CAG-eGFP-WPRE-bGHpA in place of eGFP (enhanced green fluorescent protein) by standard molecular biology techniques. Cells were harvested 48 hr after transfection and the virus was purified essentially as described (Zolotukhin et al., 2002; Grieger et al., 2006), but with some modifications. Briefly, harvested cells were lysed by three freeze–thaw cycles and treated with 0.5% sodium deoxycholate (Sigma-Aldrich, St. Louis, MO) and Benzonase endonuclease (Sigma-Aldrich) at 50 U/ml before clarification by centrifugation. The clarified lysate was purified by ultracentrifugation on a discontinuous iodixanol gradient (OptiPrep; Axis-Shield). All serotypes and clones, except rAAV2/9, were then further purified by manual anion-exchange chromatography on 1-ml Bio-Scale UNOsphere Q columns (Bio-Rad, Hercules, CA). Briefly, after preequilibration of the column with buffer AI (20 mM Tris, 15 mM NaCl, pH 8.5) the vector-containing iodixanol fraction was diluted 1:10 with buffer AI, loaded on the column, and washed with 10 ml of buffer AI. The vector was eluted with a 1-ml step gradient containing 50, 100, 150, 200, 250, and 500 mM NaCl in 20 mM Tris, pH 8.5. The vector-containing fraction was determined by quantitative real-time polymerase chain reaction (PCR), aliquoted, and stored at −80°C. Alternatively, some batches of AAV2/6 were purified by manual heparin affinity chromatography on HiTrap heparin HP 1-ml columns (GE Healthcare Life Sciences, Piscataway, NJ) by an adaptation of previously published methods (Halbert et al., 2001; During et al., 2003). Briefly, after preequilibration of columns with buffer AH (100 mM NaCl, 20 mM Tris, pH 7.4) the vector-containing iodixanol fraction was diluted 1:5 in buffer AH, loaded on the column, and washed with 10 ml of buffer AH. The vector was eluted with a 3-ml step gradient containing 150, 200, 300, and 500 (× 2) mM NaCl in 20 mM Tris, pH 7.4. The 300 and 500 mM NaCl fractions were pooled, concentrated in a 4-ml 100K NMWL (nominal molecular weight limit) ultracentrifugal concentrator (Amicon Ultra-4; Millipore, Bedford, MA), and then washed with cold phosphate-buffered saline (PBS). The concentrated sample was filter sterilized (Acrodisc 13-mm syringe filters with 0.2-μm HT Tuffryn membrane; Pall, Port Washington, NY), aliquoted, and stored at −80°C. AAV9 could not be purified by ion-exchange chromatography and was further purified by a second round of ultracentrifugation on a continuous iodixanol gradient (Hermens et al., 1999). The vector-containing fraction was determined by quantitative real-time PCR, aliquoted, and stored at −80°C.
Virus quantification
Viral genome copies per milliliter (VG copies/ml) were determined by quantitative real-time PCR. Samples were treated with DNase I (Roche, Indianapolis, IN) and then digested with proteinase K (Roche) in 0.5% sodium dodecyl sulfate (SDS) and diluted 1:100 in water before PCR. Real-time quantitative PCR was performed on an Applied Biosystems 7900HT sequence detection system with Power SYBR green PCR master mix (Applied Biosystems, Foster City, CA) or EXPRESS SYBR GreenER qPCR supermix with premixed ROX (Invitrogen); primers targeting the woodchuck hepatitis virus posttranscriptional regulatory element (WPRE) (Mouravlev et al., 2007) or the bovine growth hormone polyadenylation signal (bGHpA) (5′-CTAGTTGCCAGCCATCTGTTGT, 5′-GGCACCTTCCAGGGTCAAG) were used. A standard curve was prepared by serial dilution of a linearized plasmid (Wang et al., 2006). Transduction units per milliliter (TU/ml) were determined by infection of HeLa cells and detection of eGFP-positive cells by flow cytometry as previously described (Reed et al., 2006); all serotypes and clones were able to transduce HeLa cells (data not shown).
Infection of monocytes and MoDCs
MoDCs were seeded at 1 × 105/well in a flat-bottomed 96-well plate (Greiner Bio-One) in serum-free sRPMI and infected at the indicated multiplicity of infection (MOI). In some experiments virus was preincubated in heparin (Sigma-Aldrich) at the concentrations indicated for 1 hr before infection. Three hours postinfection the medium was replaced with sRPMI with 10% fetal bovine serum, IL-4 (50 ng/ml), and GM-CSF (100 ng/ml). Infected MoDCs were cultured at 37°C/5% CO2 for 48 hr and then assessed for transgene expression ± immunophenotype by flow cytometry. In some experiments monocytes, seeded at 1 × 105 per well of a 96-well plate, were infected by the same method at the indicated MOI. Infected monocytes were cultured in sRPMI with 10% fetal bovine serum, IL-4 (50 ng/ml), and GM-CSF (100 ng/ml) at 37°C/5% CO2 for 48 hr before analysis by flow cytometry.
Flow cytometry
Flow cytometry was performed on a BD FACSCalibur (BD Biosciences, San Jose, CA) and data analyzed with CellQuest (BD Biosciences) or FlowJo (Tree Star, Ashland, OR). The fluorophore-conjugated antibodies against the following antigens were used: allophycocyanin-conjugated CD1a (CD1a–APC), HLA-DR–APC, peridinin–chlorophyll–protein-conjugated CD8 (CD8–PerCP), phycoerythrin-conjugated CD107a (CD107a–PE), and fluorescein isothiocyanate-conjugated interferon-γ (IFN-γ–FITC) from BD Biosciences; CD14–Alexa 647, CD14–FITC, CD83–PE, and CD86–PE from Serotec (Oxford, UK); CD83–PE, CD86–PE, and HLA-DR–APC from BioLegend (San Diego, CA). The viability stain 7
Generation of capsid mutants
Capsid mutants were generated by site-directed mutagenesis with a QuikChange II site-directed mutagenesis kit (Stratagene, La Jolla, CA) as per the manufacturer's instructions. To generate AAV6 capsid tyrosine mutants prospective sites for mutation were identified by sequence alignment of AAV6 and AAV2 cap genes and comparison made with previously reported mutations of the AAV2 cap gene (Zhong et al., 2008). The following primers were used (mutated bases in boldface, restriction sites introduced by silent mutations underlined): Y445F sense, CCTCTCATCGACCAGTAC
Infection of HeLa cells with capsid mutants
HeLa cells were seeded at a density of 4 × 104 cells per well of a flat-bottomed 96-well plate (Greiner Bio-One) in Dulbecco's modified Eagle's medium (DMEM; Invitrogen) supplemented with 10% FBS, penicillin (60 μg/ml) (Sigma-Aldrich), and streptomycin (140 μg/ml) (Sigma-Aldrich). The next day half the medium was removed and the cells were infected in triplicate at an MOI of 1 × 103 or 1 × 104 with vector pseudotyped with either the wild-type AAV6 capsid or with one of the three mutants (Y445F, Y731F, double mutant). After 1 hr an additional 50 μl of medium was added per well. Two days after infection cells were harvested with trypsin–EDTA (Invitrogen), stained with 7AAD (BD Biosciences) to exclude nonviable cells, and assessed by flow cytometry for eGFP expression.
Stimulation of HBc18–27-specific CD8+ T cell clone
Day 6 MoDCs were seeded at 1 × 105 cells per well in a U-bottomed 96-well plate (Greiner Bio-One) and infected at an MOI of 1 × 105 with AAV6/HBcAg or AAV6/eGFP as described previously. After 24 hr the 10 mM HBc18–27 peptide FLPSDFFPSV (Sigma-Aldrich) was added to the positive control. Two days postinfection the cells were washed twice in RPMI, and then resuspended in 100 μl of sRPMI–10% FBS. Cells (1 × 105) of the HBc18–27 (clone C18-3)-specific CD8+ T cell clone (Gehring et al., 2007), kindly provided by A. Bertoletti and A. Gehring (Singapore Institute for Clinical Sciences, Singapore), were added in 100 μl of sRPMI–10% FBS and anti-CD107a–PE (BD Biosciences) was added. After a 1-hr incubation at 37°C/5% CO2 brefeldin A (Sigma-Aldrich) and monensin (Sigma-Aldrich) were added to 5 μg/ml and the plate was incubated for a further 5 hr at 37°C/5% CO2. Samples were stained with anti-CD8–PerCP (BD Biosciences), fixed with Cytofix/Cytoperm solution (BD Biosciences), and stained with anti-IFN-γ–FITC (BD Biosciences). Samples were analyzed by flow cytometry as outlined previously.
Statistical analysis
Probability testing was performed with SigmaPlot 11.0 (Systat Software, San Jose, CA). Multiple groups were compared by one-way analysis of variance (ANOVA). If a statistically significant difference was detected between the groups (p < 0.05) then pair-wise multiple comparison was performed by the Holm–Sidak method. Capsid mutants and the effect of heparin were compared with wild type by a one-sample t test. Means (or geometric means) and 95% confidence intervals are presented throughout the paper.
Results
rAAV2/6 exhibits superior transduction of human MoDCs
Expansion of the number of AAV serotypes has enabled the development of rAAV vectors suited to the delivery of genes to distinct cells and tissue. Previous studies into the ability of rAAV to transduce human MoDCs have used a limited range of serotypes. We sought to determine whether AAV serotypes previously untested in these cells might exhibit greater transduction ability. To identify the serotype or clone of AAV with optimal tropism for human MoDCs we initially infected cells with rAAV2 pseudotyped with capsids from six distinct serotypes (AAV serotype 1, 2, 5, 6, 8, 9) or three variants (rh8, rh10, rh13) at an MOI of 1 × 105 VG copies per cell. In addition, we examined whether the tropism was different when undifferentiated monocytes were infected (i.e., on day 0) compared with immature (day 6) MoDCs. Transduction was assessed 48 hr after infection by detection of eGFP expression by flow cytometry (Table 1). Only rAAV2/1, rAAV2/2, rAAV2/5, and rAAV2/6 vectors transduced more than 0.5% of day 6 MoDCs and only rAAV2/6 more than 0.5% of monocytes (Table 1). All serotypes and variants except AAV2/9 were purified from transfected cell lysates by ultracentrifugation on iodixanol gradients and ion-exchange chromatography. This method has previously been shown to maintain maximal transducing activity of vectors with a lower particle-to-infectious unit ratio compared with CsCl gradient purification (Zolotukhin et al., 2002). All vector stocks, including rAAV2/9, were of high purity as assessed by SYPRO Ruby-stained SDS–polyacrylamide gel electrophoresis (data not shown). The titers (VG copies/ml) and potency (the ratio of viral genome copies to transduction units) of vector stocks are presented in Table 2. Although it is not possible to compare vector potency between different pseudotypes or with previous reports that use a different transduction assay (Aucoin et al., 2008), the potency within a pseudotype was reasonably consistent between preparations (Table 2). Further, the potency of rAAV2/2 was similar to that described in a previous report that used the same transduction assay (Reed et al., 2006). In addition, in the initial screen of pseudotypes two independent batches of rAAV2/2, rAAV2/5, and rAAV2/rh8 were used with similar results. Therefore our findings are unlikely to be influenced by significant differences in vector potency between preparations.
Transduction defined as fluorescence in FL-1 > 99.9% of the uninfected control.
rAAV2/6 vs. rest, p < 0.001, other differences not significant.
rAAV2/6 vs. rest, p < 0.001, other differences not significant.
CI, confidence interval; rAAV, recombinant adeno-associated virus.
Viral genome (VG) copies per milliliter were determined by real-time quantitative PCR and transduction units (TU) per milliliter were determined by a transduction assay in HeLa cells, as outlined in Materials and Methods.
The ability of rAAV2/1, rAAV2, rAAV2/5, and rAAV2/6 to transduce MoDCs and monocytes was further assessed with cells isolated from three donors on two different occasions (Fig. 1). Consistent with the preliminary screen, rAAV2/6 was significantly more effective at transducing MoDCs than the next most effective vector, rAAV2 (14.2% mean transduction efficiency vs. 3.3%; p = 0.005), whereas rAAV2/1 and rAAV2/5 gave transduction efficiencies of 1.0 and 1.6%, respectively. All serotypes exhibited a greatly reduced ability to transduce monocytes compared with MoDCs when transgene expression was determined after 48 hr (Fig. 1B). Only a modest decline in transduction efficiency was observed with rAAV2/6 when the MOI was reduced to 1 × 104 (Fig. 1C); transduction efficiency with rAAV2/6 at an MOI of 1 × 104 remained higher than with 10-fold more rAAV2/1, rAAV2, or rAAV2/5 (Fig. 1A and C). These experiments were performed with three independent preparations of rAAV2/6, two independent preparations of rAAV2/2 and rAAV2/1, and a single preparation of rAAV2/5.
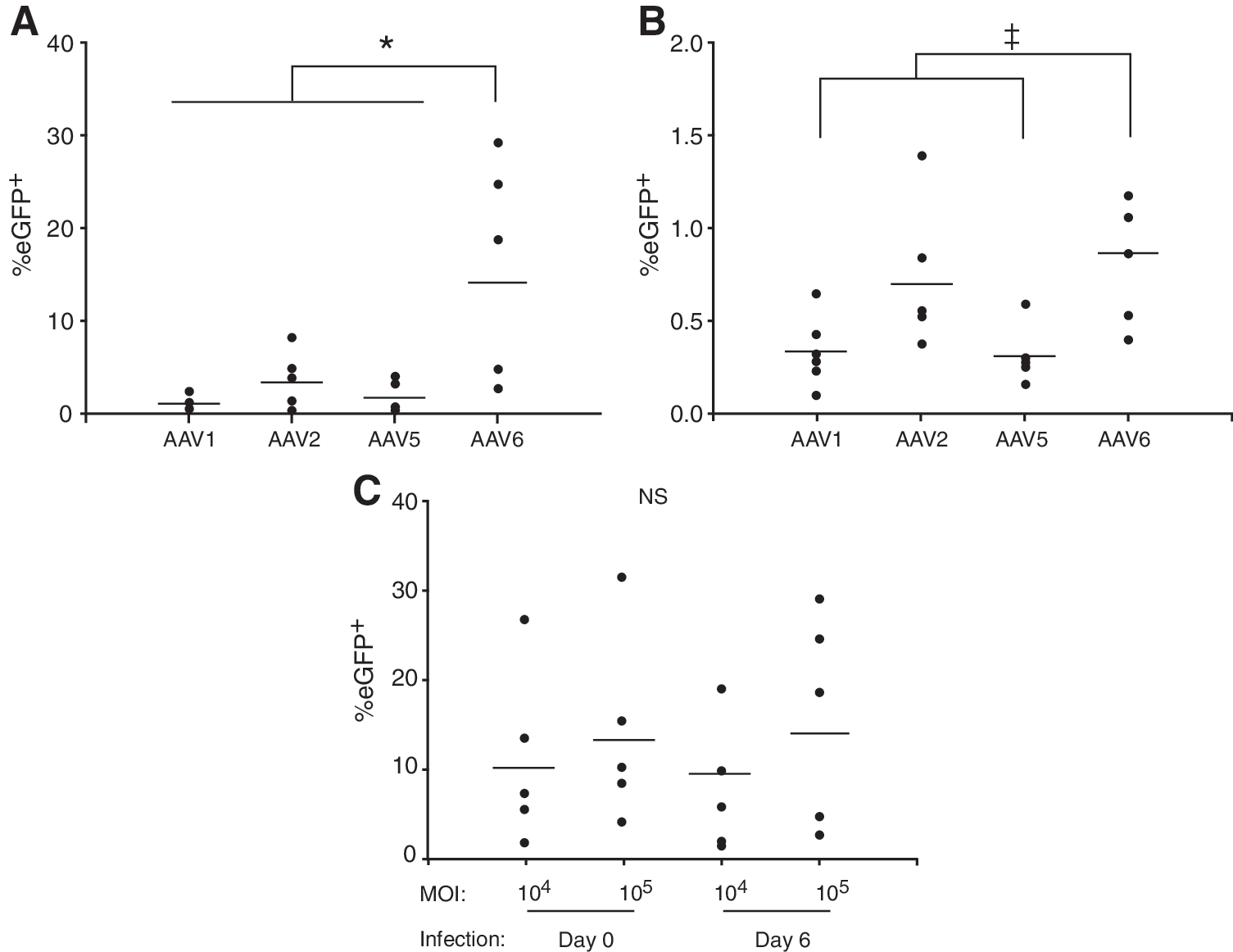
Effect of rAAV serotype and multiplicity of infection (MOI) on the transduction of human monocyte-derived dendritic cells (MoDCs).
Transduction of MoDCs by rAAV2/6 is abolished by a lysine-to-arginine mutation in the capsid. Despite 99.2% sequence similarity between the capsids of AAV1 and AAV6, rAAV2/1 failed to transduce MoDCs in our experiments. Wu and colleagues demonstrated that a single lysine-to-glutamate mutation (K531E) in the capsid of AAV6 reduced both liver transduction efficiency and the ability to bind heparin whereas the converse mutation in the AAV1 capsid led to the acquisition of these properties (Wu et al., 2006a). To probe the molecular basis for the distinct tropism of rAAV2/6 for MoDCs we assessed the effects of the K531E mutation in the AAV6 capsid and the converse mutation in AAV1 on the ability of vectors to transduce MoDCs. Figure 2A indicates that this mutation significantly reduces the transduction of MoDCs by rAAV2/6 whereas the converse mutation in AAV1 (E531K) restored a level of transduction comparable to that of the wild-type rAAV2/6 vector. Soluble heparin only partially inhibited the transduction of MoDCs by rAAV2/6 at the highest concentration tested (1 mg/ml). By contrast, transduction of MoDCs with rAAV2 was susceptible to inhibition by soluble heparin at a 10-fold lower concentration (Fig. 2B). These experiments suggest that rAAV2/6 does not use heparin as a receptor on MoDCs despite being able to bind immobilized heparin.

The tropism of rAAV2/6 for MoDCs is determined by Lys-531, not by its ability to bind heparin.
Mutation of surface-exposed tyrosine residues improves transduction of MoDCs by rAAV2/6
“Next-generation” rAAV2 vectors have been described in which surface-exposed tyrosine residues in the capsid were mutated to block phosphorylation of capsids by epidermal growth factor receptor protein tyrosine kinase (EGFR-RTK) (Zhong et al., 2008). These mutations led to increased transduction of HeLa cells in vitro and murine hepatocytes in vivo through a reduction in the ubiquitination of capsids, leading to increased nuclear translocation. Alignment of the sequences encoding polypeptide VP1 from AAV2 and AAV6 revealed a high degree of conservation; six of the seven surface-exposed tyrosine residues from AAV2 are conserved in AAV6. Of the three tyrosine residues in the AAV2 sequence associated with the greatest improvement in transduction efficiency when mutated (Y444, Y500, Y730), Y500 (Y501 in AAV6) encodes phenylalanine in the AAV6 VP1 polypeptide (Fig. 3A). To investigate whether mutation of surface-exposed tyrosines would improve transgene expression in rAAV2/6-transduced MoDCs, Y445 and Y731 in the AAV6 capsid were mutated to phenylalanine and an additional double mutant (Y445F + Y731F) was generated. Transduction of HeLa cells with vectors containing these mutations confirmed their ability to increase the rate of transduction, albeit at a reduced level compared with that reported previously for rAAV2 (Fig. 3B). When the effect of tyrosine mutation on the transduction of MoDCs was assessed, a modest (approximately 2-fold) increase in transgene expression was observed with the Y731F mutant. Surprisingly, both the Y445F mutant and double mutant resulted in a significant decrease in transduction (Fig. 3C). We were unable to assess whether this decrease in transduction efficiency was due to proteasomal degradation, as the proteasome inhibitors were highly toxic to MoDCs (data not shown).
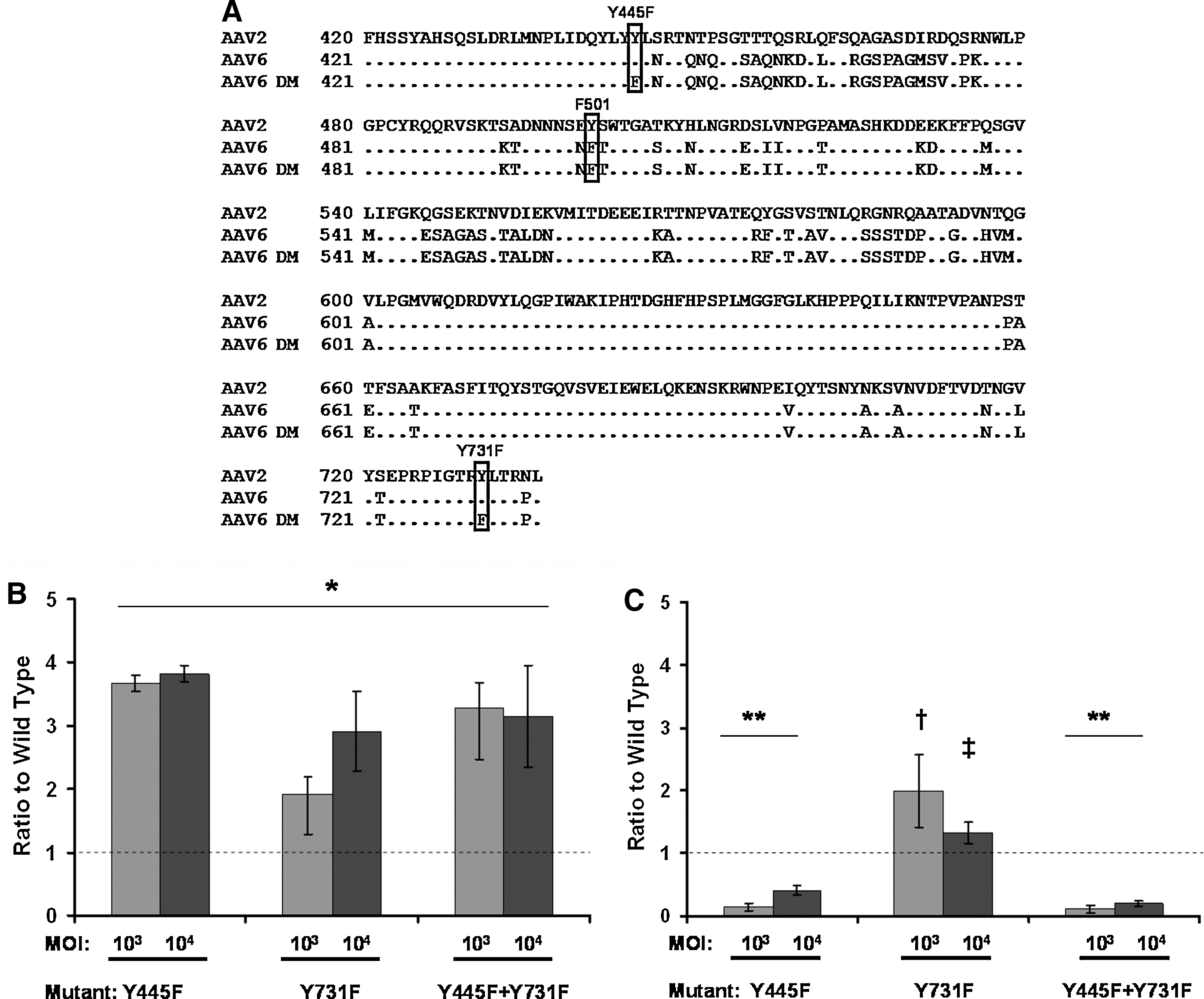
Mutation of surface Tyr-731 improves the transduction of HeLa cells and MoDCs by rAAV2/6.
Immunostimulatory potential of MoDCs transduced with rAAV2/6 (Y731F)
We next examined the effect of infection with rAAV2/6 (Y731F) on the immunophenotype of MoDCs. Day 8 MoDCs that had been infected with rAAV2/6 (Y731F) on day 6 were stained for immunophenotypic markers and assessed by flow cytometry. On day 8 the expression levels of CD83, CD86, and HLA-DRα, which are upregulated on DC maturation, were only partially upregulated in infected compared with uninfected MoDCs, suggesting that infection with rAAV2/6 (Y731F) is unable to trigger MoDC maturation (Fig. 4A and B).
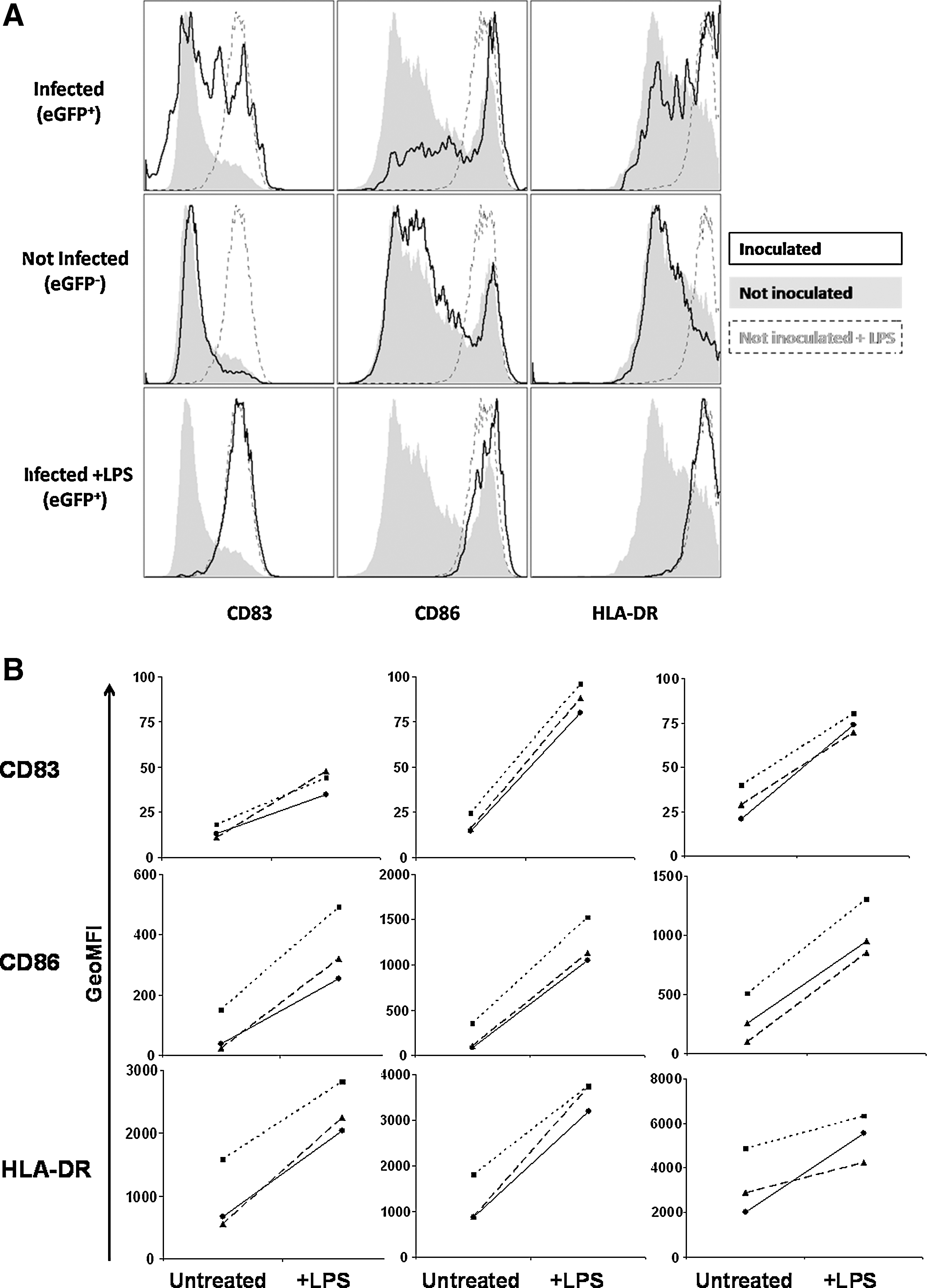
Partial upregulation of costimulatory markers in MoDCs transduced with rAAV2/6 (Y731F).
Stimulation of a naive T cell response by dendritic cells requires the increased surface expression of costimulatory molecules and secretion of cytokines that are associated with maturation of the DCs (Banchereau et al., 2000). To establish whether rAAV2/6 (Y731F)-transduced MoDCs were capable of undergoing maturation necessary for T cell stimulation, cells were infected with rAAV2/6 (Y731F) expressing eGFP and, 24 hr later, treated with the Toll-like receptor-4 (TLR4) agonist lipopolysaccharide (LPS). Immunophenotype was assessed after a further 24 hr by flow cytometry. Infection with rAAV2/6 (Y731F) had no effect on the ability of MoDCs to mature (Fig. 4A and B). Last, we confirmed that antigen encoded by rAAV2/6 (Y731F) was correctly processed and presented to T cells by transduced MoDCs. Day 6 MoDCs isolated from HLA-A2-positive donors were infected with rAAV2/6 (Y731F) encoding the core antigen from hepatitis B virus (HBcAg). Forty-eight hours after infection rAAV-infected MoDCs were incubated with a CD8+ T cell clone specific for the HLA-A2 HBcAg epitope 18–27. Stimulation of the T cell clone was assessed by CD107a expression and flow cytometry. Antigen-specific CD107a expression was seen with AAV2/6 (Y731F) vectors expressing HBcAg (Fig. 5). Of note, as this clone is not antigen naive only T cell receptor signaling is necessary for activation; costimulatory signals are not required (Gehring et al., 2007).
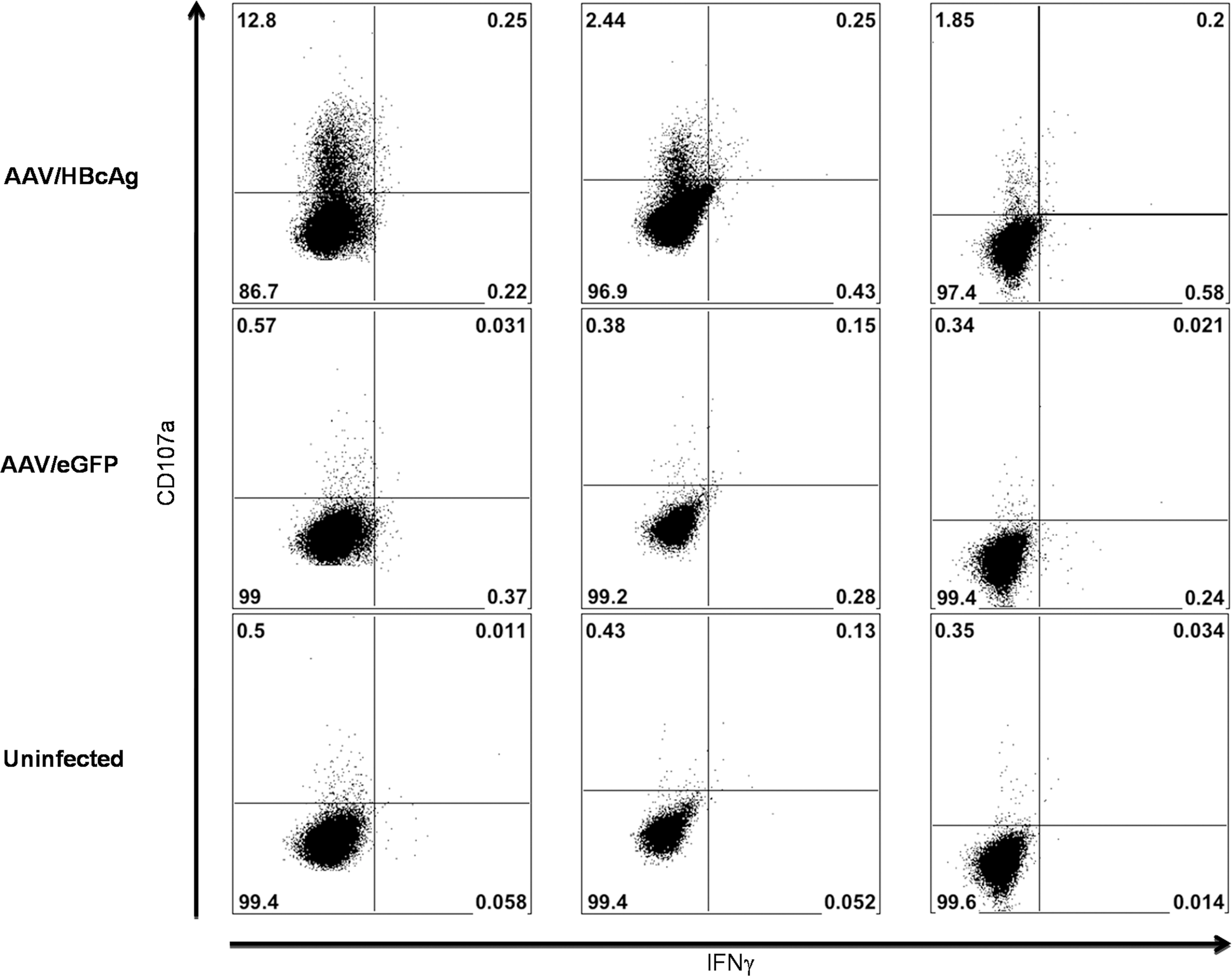
Vector-encoded antigen is correctly processed and presented to a CD8+ T cell clone by transduced MoDCs. MoDCs from HLA-A2+ donors were either infected with rAAV2/6 (731F) encoding HBcAg or eGFP or mock infected. Forty-eight hours postinfection MoDCs were incubated with a costimulation-independent CD8+ T cell clone specific for the HLA-A2-restricted HBcAg 18–27 epitope. Cells were stained for surface expression of CD107a and CD8 and intracellular expression of IFN-γ and the cells were analyzed by flow cytometry. Expression of CD107a and IFN-γ by CD8+ cells is shown (n = 3).
Discussion
The availability of AAV serotypes with distinct tissue tropism has fostered their widespread use as gene transfer vectors in vitro and in vivo (Gao et al., 2005). The pivotal role played by dendritic cells in the initiation and regulation of immune responses has prompted several groups to examine their permissiveness to transduction by rAAV vectors. In this study we examined the tropism of rAAV vectors pseudotyped with capsids derived from AAV serotypes 1, 2, 5, 6, 8, and 9 and the variants rh8, rh9, and rh10 from nonhuman primates for human peripheral blood monocytes and immature monocyte-derived dendritic cells (MoDCs). Undifferentiated (day 0) monocytes were markedly less permissive than immature MoDCs (cultured for 6 days with GM-CSF and IL-4) (1.95 vs. 9.85% transduction with rAAV2/6). In a subsequent assessment with cells from three donors rAAV2/6 mean transduction levels at an MOI of 105 per cell reached 13.8% compared with 3.6% for rAAV2 and 1.3% for rAAV2/1. Significant donor-to-donor variation was observed with all serotypes, consistent with previous studies (Ponnazhagan et al., 2001; Shin et al., 2008).
The transduction of mature DCs was not assessed. In vivo, immature DCs represent a more relevant target for vaccination. Immature DCs are resident in the peripheral tissues, where they first encounter antigen; maturation is a continuous process that, while initiated in the periphery, continues during migration to the lymph nodes and is completed on interaction with T cells (Banchereau et al., 2000). Indeed, preactivation of DCs impairs MHC presentation of exogenous and endogenous antigens (Young et al., 2007). Whereas the tropism of various serotypes might differ with the stage of differentiation of MoDCs no difference was seen in the hierarchy of serotypes between monocytes and immature MoDCs.
There are conflicting reports concerning which AAV serotype exhibits the greatest tropism for human MoDCs. Initial reports of DC transduction by rAAV used rAAV2 (Liu et al., 2000; Ponnazhagan et al., 2001). Subsequent studies showed that the transduction efficiency of MoDCs by rAAV2 and rAAV2/1 was less than 1% but increased 10-fold when vectors with self-complementary genomes were used, indicating that second-strand synthesis might be a limiting factor in transduction, consistent with results obtained in other cell types (Veron et al., 2007). A comparison of AAV serotypes in human and mouse DCs revealed rAAV5/5 to be significantly more effective than rAAV2 and all other serotypes tested (serotypes 1, 2, 5, 7, and 8) (Xin et al., 2006). Most recently, a comparison of self-complementary vectors pseudotyped with serotypes 1–8 identified rAAV serotypes 2, 5, and 6 as possessing a similarly high transduction ability (>20%) even at an MOI 100-fold lower than in the studies mentioned previously (Shin et al., 2008). Notably, Shin and colleagues assessed eGFP expression at 4 days postinfection whereas our assessments were made at 48 hr. On the basis of our experiments we conclude that rAAV2/6 exhibits significantly higher transduction efficiencies in human MoDCs that the other serotypes and variants examined here, with an efficiency approximately 5-fold greater than that of rAAV2 and 10-fold greater compared with all other serotypes tested. The rates of transduction seen with rAAV2/1, rAAV2/2, and rAAV2/8 in our study were similar to those seen in previous studies using the same MOI (Xin et al., 2006; Veron et al., 2007). However, in contrast to a previous report (Xin et al., 2006), we found rAAV pseudotyped with AAV5 capsid to be no more efficient at transducing MoDCs than rAAV2/1 and rAAV2. This discrepancy may be due to the use of pseudotyped vectors in our study whereas Xin and colleagues made rAAV5/5 in which the genome had AAV5 ITRs, which may influence other aspects of the viral transduction. Indeed, a further study (Aldrich et al., 2006) reported poor transduction of murine DCs by rAAV2/5 whereas Xin and colleagues found high levels of transduction with rAAV5/5.
The molecular basis for the superior tropism of rAAV2/6 for MoDCs is intriguing. The genome sequence of AAV6 suggests that it is a hybrid that arose through recombination between AAV1 and AAV2 (Xiao et al., 1999). Despite the high degree of sequence similarity between the capsids of AAV1 and AAV6 (99.2%), a study observed that these viruses vary significantly in their ability to transduce liver in vivo (Wu et al., 2006a). These authors demonstrated that a single lysine-to-glutamate mutation (K531E) in the capsid of AAV6 reduced both liver transduction efficiency and the ability to bind heparin whereas the converse mutation in AAV1 increased liver transduction and imparted heparin-binding affinity (Wu et al., 2006a); K531 is unique to AAV6 among other AAV serotypes (Rutledge et al., 1998; Wu et al., 2006a). We speculated that the affinity of rAAV2/6 for heparin may be responsible for its superior transduction of MoDCs. A study that evaluated natural and engineered AAV variants showed direct correlations between heparin binding, uptake into MoDCs, and activation of a capsid-specific T cell response, although rAAV2/6 was not included in this study (Vandenberghe et al., 2006). Interestingly, the immunogenicity and toxicity of vectors based on human adenovirus type 5 have also been correlated with its ability to use a heparin-sensitive receptor on dendritic cells (Cheng et al., 2007). We confirmed that K531 is important for the ability of rAAV2/6 to transduce MoDCs, with the E531K mutation in AAV1 conferring levels of transduction similar to that of rAAV2/6. Surprisingly, however, heparin did not inhibit transduction of MoDCs by rAAV2/6 except at 1 mg/ml, where only partial inhibition was seen; in contrast, rAAV2 was inhibited at 0.1 mg/ml. This suggests that although residue K531 is critical for efficient MoDC transduction heparan sulfate proteoglycans are unlikely to act as a receptor for rAAV2/6. We cannot exclude the possibility that other positively charged glycans other than heparin sulfate are involved in the binding of AAV6 to MoDCs. Alternatively, K531 may be important in the recognition of a protein receptor that is present on MoDCs or may influence intracellular trafficking. These issues should be addressed in future studies.
Although rAAV2/6 exhibits a high affinity for MoDCs, we sought to determine whether its intrinsic tropism could be further increased by the selective mutation of amino acid residues in the capsid. A study demonstrated that mutation of surface-exposed tyrosines in the capsid of AAV2 led to increased transduction in vitro and in vivo (Zhong et al., 2008). Six of the seven tyrosine residues are conserved between AAV2 and AAV6, suggesting that their functional significance is also likely to be conserved. We selected those residues reported to have the greatest effect on rAAV2 transduction titers after mutation to phenylalanine (Zhong et al., 2008). Transduction of HeLa cells with mutant rAAV2/6 confirmed that the mutants increased the transduction efficiency, although by a significantly lower amount than reported for rAAV2. The relatively small increase in transduction efficiency may be due to the phenylalanine at position 501 in AAV6 as opposed to the tyrosine in AAV2; no further increase in transduction was seen with the rAAV2/6 double mutant, suggesting that the effect of mutation of multiple tyrosine residues is not necessarily additive. This is consistent with a report of multiple mutations of the AAV2 capsid (Zhong et al., 2009). We then examined the potential of mutants to increase the transduction of MoDCs (Fig. 3C). Surprisingly, two capsid mutations that increased the transduction of HeLa cells caused a significant decrease in the transduction of MoDCs, whereas the remaining mutant (Y731F) resulted in an approximate 2-fold increase in transduction. Attempts to determine whether transduction efficiency could be increased by proteasome inhibition were unsuccessful because of the toxicity of inhibitors to the cells. The effect on transduction of mutations at selected capsid tyrosine residues differed between HeLa cells and MoDCs. The precise reason(s) for this difference is not immediately obvious from our experiments. One plausible explanation is that epidermal growth factor receptor protein tyrosine kinase (EGFR-PTK), which is responsible for the phosphorylation of capsid tyrosine residues leading to ubiquitination and proteasomal degradation (Zhong et al., 2008), is differentially expressed in these two cell types. Indeed, there are conflicting reports as to whether EGFR is expressed in MoDCs (Tillman et al., 1999; Cao et al., 2008). Thus the greater effect of these mutations in HeLa cells may reflect greater EGFR-PTK activity compared with DCs. Alternatively, AAV6 may use distinct cell surface receptors on HeLa cells and DCs and the effect of mutating capsid tyrosine residues could impact selectively on receptor binding on DCs alone. Notably, EGFR has been identified as a coreceptor for AAV6 (Weller et al., 2010). It has also been reported that the Y445F mutation in AAV6 has no effect on the transduction of human CD34+ hematopoietic stem cells whereas the Y731F and Y705F mutations led to a 2- to 3-fold increase over wild type (Jayandharan et al., 2009). The corresponding Y704F mutation in the AAV2 capsid resulted in only a modest increase in transduction efficiency in HeLa cells compared with Y444F, Y500F, and Y730F (2-fold vs. 8- to 11-fold) (Zhong et al., 2008). Therefore the effect of mutation of surface-exposed tyrosine residues on transduction efficiency is likely dependent on both the virus serotype and target cell. In future studies it would be interesting to compare the transduction efficiency of Y731F and Y705F mutants in MoDCs.
Antigen presentation to naive T cells by mature dendritic cells stimulates proliferation and memory differentiation; in contrast, presentation by immature dendritic cells leads to tolerance (Steinman and Nussenzweig, 2002; Mescher et al., 2006). Mature dendritic cells provide signals 1, 2, and 3 to T cells, presenting antigen via MHC class I and II molecules (signal 1), expressing high levels of costimulatory molecules such as CD80 and CD86 (signal 2), and secreting cytokines, such as interleukin-12 or type I interferons (signal 3) (Curtsinger et al., 1999; Mescher et al., 2006). We assessed the immunophenotype of MoDCs transduced with rAAV2/6 (Y731F) and found only minor upregulation of CD86 and other markers of maturation (HLA-DRα and CD83; Fig. 4A and B), consistent with previous findings (Veron et al., 2007; Shin et al., 2008). Dendritic cells mature in response to signals via innate pathogen recognition receptors, such as the Toll-like receptors (TLRs) (Banchereau et al., 2000). Studies of gene expression in human cell lines and in mice in vivo suggest that infection with rAAV causes minimal activation of innate immune signaling pathways (Zaiss et al., 2002; McCaffrey et al., 2008). Importantly, however, MoDCs transduced by rAAV2/6 (Y731F) are still able to mature in response to stimulation by the TLR-4 ligand LPS (Fig. 4A and B). Thus coadministration of an adjuvant may improve the immunogenicity of rAAV as a vaccine vector. Alternatively, unadjuvanted rAAV may be a suitable vaccine vector for induction of tolerance (Han et al., 2005). Last, we confirmed that MoDCs transduced with rAAV2/6 (Y731F) are able to process and present the vector-encoded transgene on MHC class I molecules as assessed by their ability to stimulate a costimulation-independent CD8+ T cell clone, which requires only signal 1 to respond (Fig. 5) (Gehring et al., 2007).
In conclusion, this study reports the novel finding that rAAV vectors pseudotyped with serotype 6 are significantly more efficient at transducing human MoDCs. Lysine-531 is a critical determinant of MoDC tropism through a mechanism that is independent of its effect on heparin binding. Transduction efficiency can be increased further by mutation of the capsid tyrosine residue at position 731, consistent with a study on rAAV/2 (Zhong et al., 2008). Transduction of MoDCs with rAAV2/6 led to only minor upregulation of surface markers of MoDC maturation. Therefore future vaccine studies with rAAV should use rAAV2/6 (Y731F) in combination with an adjuvant.
Footnotes
Acknowledgments
This research was conducted during the tenure of a Clinical Research Training Fellowship from the Health Research Council of New Zealand (J.U.). The authors thank Dr. Deborah Young (University of Auckland) and Prof. James Wilson (University of Pennsylvania) for the provision of plasmids used in the production of rAAV, and Dr. Antonio Bertoletti and Dr. Adam Gehring (Singapore Institute for Clinical Sciences) for providing the HBc18–27-specific CD8 T cell clone.
Author Disclosure Statement
No competing financial interests exist.