
Abstract
Our objective was to achieve the enhanced delivery of vascular endothelial growth factor (VEGF) to ischemically disordered brain through transferrin-coupled liposomes (Tf-PLs) via intravenous administration, and to observe the effect of Tf-VEGF-PLs on ischemic brain neuroprotection and angiogenesis. Cerebral VEGF overexpression was achieved with Tf-PLs by intravenous injection 48 hr after an acute stroke. β-Galactosidase expression was monitored; saline was injected as a control. The success of postischemic gene transduction was confirmed by β-galactosidase staining and by increased VEGF mRNA and protein in ischemic brain. Vascular density, neurological recovery, and ischemic area calculation were performed to evaluate the effect of Tf-VEGF-PLs. The positive expression of β-galactosidase indirectly indicated that VEGF was successfully delivered into brain by Tf-VEGF-PLs. VEGF mRNA in the Tf-VEGF-PL group 24 hr after injection was significantly higher than in the control group (p < 0.05). Western blot analysis showed that postischemic Tf-VEGF-PLs resulted in increased VEGF protein levels compared with VEGF-PLs and saline-administered rats (p < 0.05) 48 hr after administration. At 21 days after drug injection, we observed a significant decrease in infarct volume and better neurological function in the Tf-VEGF-PL-treated group, compared with the VEGF-PL group. FITC–dextran marking showed increased vascular density in the penumbra of Tf-VEGF-PL-treated hemispheres (245,873.9, number of microvessels per field) compared with that in VEGF-PL-treated hemispheres (139,801.3) or saline-treated hemispheres (102,175.5) (p < 0.05). The remainder of the cerebral blood flow after ischemia in the Tf-VEGF-PL group was significantly more than in the control groups (0.35 vs. 0.29, 0.21; p < 0.05). We conclude that the VEGF gene can be delivered noninvasively into the brain by Tf-VEGF-PLs. Postischemic treatment with Tf-VEGF-PLs effectively promoted neuroprotection and vascular regeneration in the chronic stage of cerebral infarction.
Introduction
The blood–brain barrier (BBB) protects the CNS from potential harmful exogenous and endogenous molecules (Smith and Gumbleton, 2006). Because of the poor permeability of the BBB, gene therapy drugs cross it only with difficulty. In addition, most current gene vectors do not cross the BBB after intravenous administration and must be given via craniotomy or intracerebral injection, both considered highly invasive and unable to deliver exogenous genes to global areas of the brain (Pardridge, 2002; Schlachetzki et al., 2004). Given the evident side effects of viral vectors (Pardridge, 2001), the goal of brain-targeting technology may benefit from an efficient, noninvasive, and nonviral method of gene therapy.
The BBB possesses specific receptor-mediated transport mechanisms that potentially can be exploited as a means to target drugs to the brain (Qian et al., 2002; Hatakeyama et al., 2004). Some receptors expressed on the BBB, for example, the transferrin (Tf) receptor and insulin receptor, have been used for brain-targeting drug delivery research (Pardridge, 1999). Liposomes, a kind of drug carrier, have attracted increasing interest not only for the enhancement of therapeutic efficacy but also for the reduction of systemic side effects. Previous studies have reported that PEGylated immunoliposomes could cross the BBB via Tf receptor-mediated transcytosis and deliver exogenous genes into the brain without damaging the BBB (Shi and Pardridge, 2000; Zhang et al., 2003).
In this study, VEGF-loaded liposomes stabilized by PEGylation (VEGF-PLs) were prepared and Tf was conjugated to the surface of VEGF-PLs to formulate Tf-targeted VEGF-PLs (Tf-VEGF-PLs). The effects of Tf-VEGF-PLs and VEGF-PLs were both studied in permanent focal cerebral ischemic rat models by intravenous administration via the femoral vein, rather than by craniotomy or intracerebral injection. Tf-targeted, LacZ-loaded PLs (Tf-LacZ-PLs) was used to ensure that PLs had been successfully delivered into the brain by positive expression of the LacZ gene. The same volume of saline was injected as control. We hypothesized that animals treated with Tf-VEGF-PLs would regain most neurological functions on recovery.
Materials and Methods
Materials
1-Palmitoyl-2-oleoyl-sn-glycerol-3-phosphocholine (POPC), didodecyl dimethyl ammonium bromide (DDAB), polyethylene glycol distearoylphosphosphatidylethanolamine (PEG2000-DSPE), and maleimide-derivatized PEG2000-DSPE (Mal-PEG2000-DSPE) were purchased from Avanti Polar Lipids (Alabaster, AL). Cholesterol (Chol) was bought from Tokyo Kasei Kogyo (Tokyo, Japan). A NanoOrange protein quantitation kit was purchased from Invitrogen (Carlsbad, CA). 2-Iminothiolane (Traut's reagent) and Sepharose CL-4B were obtained from Sigma-Aldrich (St. Louis, MO). Polycarbonate membranes (pore size, 400, 200, and 100 nm; Isopore) were purchased from Millipore (Cork, Ireland). Sephadex G-25 was purchased from Amersham Biosciences (Baied'Urfe, Quebec, PQ, Canada). All other chemicals were of the highest grade possible.
Methods
Preparation of PLs
PEGylated liposome was prepared by the thin-film hydration method as described previously (Zhao et al., 2009b). POPC, DDAB, lipid PEG2000-DSPE, and Mal-PEG2000-DSPE were first dissolved in chloroform and then evaporated under an N2 gas stream to produce a thin-layer lipid film. The dried lipid film was dispersed in 200 μl of 0.05 M Tris-HCl buffer (pH 7.0) and sonicated for 2 min with a model 3210 sonicator (Branson, The Hague, The Netherlands), followed by 2 min of vortexing. An appropriate amount of plasmid (detected with a NanoOrange kit) was added to the liposome dispersion. The liposome dispersion was frozen in ethanol–dry ice for 5 min and thawed at room temperature for 25 min. This freeze–thaw cycle was repeated six times. The liposome dispersion was diluted to a lipid concentration of 40 mM by adding 0.05 M HEPES buffer (pH 7.0), followed by extrusion three times through 400-, 200-, and 100-nm pore size filters under nitrogen gas, using an extruder (Northern Lipids, Burnaby, BC, Canada). Nonencapsulated plasmid DNA was detected with a NanoOrange kit and digested by adding pancreatic DNase (10 U), exonuclease (50 U), and 5 mM MgCl2. After incubation for 1 hr at 37°C, the reaction was stopped by adding 20 mM EDTA. The sample was applied to a 1.5 × 10 cm Sepharose CL-4B column, and the digested plasmid DNA was separated from the DNA encapsulated into liposomes. The efficiency of encapsulation of VEGF plasmid into liposomes was determined by the total amount of plasmid added in the formulation minus the amount that was not encapsulated.
Preparation of Tf-PLs
Tf was thiolated with Traut's reagent as described (Lu et al., 2005), and the extent of thiolation was determined with Ellman's regent (Bradford, 1976) (about three thiol groups per Tf). Unreacted Traut's reagent was removed by chromatography on a Sephadex G-25 column with degassed Hanks' balanced saline solution (HBSS, pH 7.4). Thiolated Tf and Mal-PEG2000-DSPE were incubated overnight at 4°C in HBSS. The mole percentage of linker lipid (Mal-PEG2000-DSPE) compared with phospholipids in the PLs was 0.2 and 2.0 mol%, respectively, whereas the amount of Tf relative to phospholipids was 100–500 μg of Tf per micromole of phospholipids. The amounts of conjugated and free Tf were indirectly measured in a protein assay (bicinchoninic acid [BCA] protein assay reagent; Pierce, Rockford, IL) (Ito et al., 2004). Tf-PL was prepared by overnight incubation of Tf-PEG2000-DSPE with preformed PLs at 4°C for 24 hr. Tf-PL was separated from unincorporated Tf by gel chromatography, using the Sepharose CL-4B column; fractions were collected and spectrophotometrically analyzed for Tf and then freeze-dried to obtain the liposome powder, as previously described (Yang et al., 2007a,b).
Characterization of Tf-PLs
The mean particle size, particle size distribution, and ζ potential of PLs were determined with a dynamic light-scattering spectrophotometer (Zetasizer ZEN3500; Malvern Instruments, Malvern, UK) after diluting with phosphate-buffered saline (PBS, pH 7.4) at room temperature. The appearance of Tf-PLs was detected by transmission electron microscopy (JEM-1010; Jeol, Tokyo, Japan). Liposome stability was determined by investigating the aggregation of liposomes in PBS, PBS with Tween 80 (3%, v/v), and PBS with 50% fetal bovine serum (FBS) at 37°C. The change in mean particle size was measured before and after a 1-hr incubation (Maeda et al., 2004).
Rat middle cerebral artery occlusion
One hundred and fifty-two specific pathogen-free (SPF)-grade adult male Sprague-Dawley rats (230–250 g) were purchased from the Animal Experimental Center of Peking Union Medical College Hospital (license CXK [Beijing], 2007-0001). Rats were housed in cages and the room was kept at a temperature of 24 ± 1°C and 50–60% humidity, under a 12 hr:12 hr light–dark cycle and with access to food and water ad libitum. Aseptic procedures were performed in the Barrier Animal Experimental Facility of Peking Union Medical College Hospital (license SYXK [Beijing], 2005-0008). Experiments were carried out in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publication No. 80-23: revised 1996). Every effort was made to minimize pain and discomfort.
Rats were anesthetized with 10% chloral hydrate (350 mg/kg, intraperitoneally). The right middle cerebral artery (MCA) was occluded by means of a monofilament suture as described previously (Longa et al., 1989; Zhao et al., 2009a). Briefly, a 4-0 surgical monofilament nylon suture with rounded tip was introduced into the right internal carotid artery through the external carotid artery stump, and advanced 1.8 cm past the carotid artery bifurcation to occlude the origin of the right MCA. Rectal temperature was monitored and maintained at 37°C with heating pads throughout the surgical procedure. The animals were allowed free access to water and food at ambient temperature (21–23°C).
Rat groupings
Two days after MCA occlusion (MCAO), rats were randomized into three groups—Tf-VEGF-PLs (1 ml, encapsulating 80 μg of VEGF), VEGF-PLs (1 ml, encapsulating 80 μg of VEGF), and saline (1 ml)—and were injected via the femoral vein. Tf-LacZ-PL (1 ml, encapsulating 80 μg of LacZ plasmid) was injected into eight rats as an indicator. Body temperature was maintained with a heating pad and rectal thermal probe set at 37°C.
Histochemical staining
β-Galactosidase histochemistry in eight rat brains removed 48 hr after intravenous injection of Tf-LacZ-PLs was determined with a Beta-Glo assay system kit (Promega, Madison, WI) according to the manufacturer's instructions. Results were recorded with a Magnifire digital charge-coupled device camera (Optronics, Goleta, CA). Controls included omitting or preabsorbing primary antibody and omitting secondary antibody.
Quantitative polymerase chain reaction
Quantitative polymerase chain reaction (PCR) was performed 24 hr after drug injection, using the SYBR green real-time PCR method. Total RNA was extracted from 24 adult rat ischemic brains (n = 8 for each group), using an Absolutely RNA microprep kit according to the manual of the manufacturer (Stratagene, La Jolla, CA). Quantitative RT-PCR was performed with an ABI 7000 PCR instrument (Applied Biosystems, Foster City, CA), using a two-stage amplification program provided by the manufacturer as follows: preliminary denaturation at 95°C for 10 min, followed by 40 cycles of denaturation at 95°C for 15 sec, annealing and elongation at 60°C for 60 sec. The specificity of the amplifications was confirmed by examination of the dissociation plot; a single peak indicated that a single DNA sequence was amplified during PCR. Each sample was tested in triplicate, and the samples obtained from three independent experiments were used for the analysis of relative transcription data, using the 2–ΔΔC T method (Livak and Schmittgen, 2001). To verify the molecular size, PCR products were run on 2% agarose gels. The primers for real-time PCR were designed with Primer Express software (Applied Biosystems).
Western blot assay
Western blotting was performed 48 hr after drug injection. Twenty-four rats (n = 8 for each group) were transcardially perfused with heparinized saline. Immediately after death, the brains were removed and homogenized. The pellet was resuspended in extraction buffer and used for Western blot analysis. Protein concentration was determined by BCA protein assay. Equal amount of proteins were loaded on a 10% acrylamide gel for electrophoresis, and were electroblotted onto a polyvinylidene difluoride (PVDF) membrane. The membrane were then probed with mouse anti-VEGF antibody (diluted 1:1000; Abcam, Cambridge, UK), which reacts specifically with VEGF that originates from humans. After probing, the membranes were then incubated with horseradish peroxidase-conjugated sheep anti-mouse IgG (Sigma-Aldrich). Protein expression was detected with an enhanced chemiluminescence detection system (GE Healthcare Biosciences, Piscataway, NJ). The image was scanned with a Canon imaging densitometer (Canon, Lake Success, NY), and protein band densities were quantified on the basis of fluorescence intensity. VEGF band intensities were normalized relative to the band intensities of β-actin, which was used as a loading control.
Neurological severity score evaluation
The modified neurological severity scores (mNSS) test (normal score, 0; maximal deficit score, 18) for 24 rats (n = 8 for each group) was performed 1, 7, 14, and 21 days after MCAO by an investigator blinded to the experimental groups as previously described (Chen et al., 2001a). This score is derived by evaluating animals for hemiparesis (response to raising the rat by the tail or placing the rat on a flat surface), abnormal movements (immobility, tremor, seizures), sensory deficits (placing, proprioception), and absent reflexes (pinna, corneal, startle), as described in detail elsewhere (Schallert et al., 1982; Li et al., 2001).
2,3,5-Triphenyltetrazolium chloride staining
To quantify infarct volume, 24 rats (n = 8 for each group) were overanesthetized with 10% chloral hydrate (400 mg/kg, intraperitoneally) 21 days after MCAO, and killed by decapitation. Each brain was rapidly taken from the skull and cooled in cold saline for 10 min. The brains were then coronally sectioned into five 2-mm sections, using a Brain Matrix blade (Electron Microscopy Sciences, Hatfield, PA). The brain slices were incubated for 30 min in a 2% solution of 2,3,5-triphenyltetrazolium chloride (TTC) at 37°C, and fixed by immersion in a 4% buffered formalin solution. Infarction size was quantified with Image-Pro Plus version 6.0 software (Media Cybernetics, Bethesda, MD). To compensate for the effect of brain edema, the corrected infarct volume was calculated according to the following formula as previously described (Swanson et al., 1990; Chen et al., 2001b): corrected infarct volume (%) = [left hemisphere volume − (right hemisphere volume − infarct volume)]/left hemisphere volume × 100. (Hemispheric infarcted areas were calculated separately on each coronal slice, from 1 to 5; each such area was defined as a percentage of the affected hemisphere [Reddy and Labhasetwar, 2009]).
Fluorescein isothiocyanate–dextran labeling and microvessel density calculation
Two days after MCAO, 24 rats (n = 8 for each group) were subjected to fluorescein isothiocyanate (FITC)–dextran (molecular weight, 2 × 106 [Sigma-Aldrich]; 1 ml of a 50-mg/ml preparation) administration via the left ventricle 1 min before the animals were killed, under deep anesthesia, by decapitation. The brains were rapidly removed and placed in 4% paraformaldehyde at 4°C for 48 hr. Green fluorochromes (FITC–dextran) on the sections were excited with a laser beam at 488 nm. Five 50-μm coronal sections at 2-mm intervals from each rat and eight views in each section adjacent to the ischemic lesion were selected for observation by laser-scanning microscopy. The segment lengths, diameters, and number of microvessels were analyzed with Image-Pro Plus version 6.0 software.
Cerebral blood flow measurement
A laser Doppler flowmetry (LDF) monitor equipped with a small-caliber probe (0.7 mm in diameter) was used to measure the surface cerebral blood flow (CBF) of 24 rats (n = 8 for each group). The laser Doppler probe was in contact with the surface of the rats' skull bone during measurement. Blood flow was recorded in the ischemic penumbral area, which is 1.0 mm lateral to the sagittal suture and 1 mm posterior to the coronal suture. Baseline blood flow was recorded 5 min before MCAO. Changes in CBF were measured 21 days after drug injection. Blood flow values were calculated and expressed as percentages of the baseline values (Shen et al., 2006).
Statistical method
All data were analyzed with the SPSS version 13.0 statistical package (SPSS, Chicago, IL). All data are presented as means ± standard deviation. Data were processed by one-way analysis of variance (ANOVA) with post hoc multiple comparisons using the Bonferroni method. Differences were considered statistically significant at p < 0.05.
Results
Characterization of Tf-PLs
The mean particle size of liposomes prepared in this study was about 100 nm (Fig. 1). The size before conjugation of Tf was in the range of 95 nm. After the addition of Tf, the average size increased between 5 and 10 nm for samples under investigation (Fig. 2). The polydispersity index (PI) did not show any significant alteration, indicating that the stability of the liposomes had not been negatively affected. The incorporation of LacZ or VEGF did not result in any significant increase in particle size when incubated for 1 hr in PBS or in PBS including 3% Tween 80, or in a solution composed of equal volume of FBS and PBS at 37°C. The mean ζ potential value was 30.2 mV. The entrapment efficiency of liposomes was 86%, the concentration of LacZ plasmid and VEGF plasmid was 80 μg/ml, and the amount of Tf in correlation to the amount of total phospholipid was 97.5 μg of Tf per micromole of phospholipid.

Particle size distribution of Tf-PLs. The average size was about 100 nm.

Transmission electron microscopy view of the PEGylated liposomes (PLs; arrows). Magnification bar: 100 nm.
Postischemic gene delivery and expression with Tf-liposomes
We first examined the level of LacZ expression after femoral injection of Tf-LacZ-PLs to determine whether postischemic trans-BBB gene delivery was successful. LacZ expression was clearly observed in the brain 48 hr after MCAO (Fig. 3). LacZ expression was also observed in the contralateral hemisphere. This demonstrated that the LacZ-encoding gene could distribute extensively in the brain with Tf-LacZ-PLs rather than being limited in a focal location with adeno-associated viral vector (Zhu et al., 2009). Real-time PCR showed that postischemic Tf-VEGF-PLs resulted in increased VEGF mRNA levels compared with VEGF-PL- and saline-administered rats (Fig. 4, p < 0.05) 24 hr after administration. Western blot analysis showed that postischemic Tf-VEGF-PLs resulted in increased VEGF protein levels compared with VEGF-PL- and saline-administered rats (Fig. 5, p < 0.05) 48 hr after administration. Together, our results showed that the VEGF gene was effectively delivered by Tf-PLs after ischemia to an extensive area of the brain, with sustained expression.
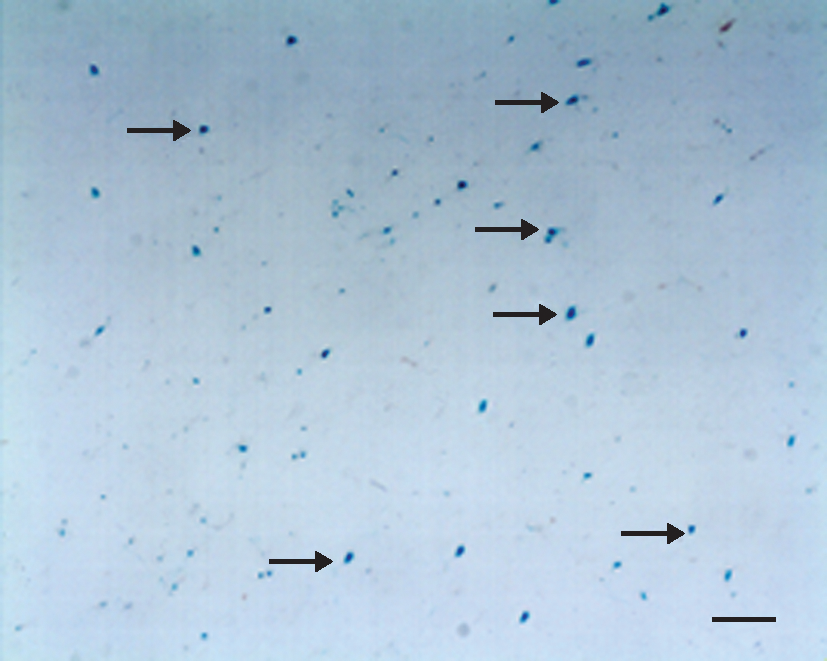
β-Galactosidase histochemistry in rat brain removed 48 hr after intravenous injection of transferrin-coupled PEGylated liposomes encapsulating the LacZ-encoding gene (Tf-LacZ-PLs). Positive blue staining (arrows) indirectly indicated that transferrin-coupled PEGylated liposomes encapsulating vascular endothelial growth factor (Tf-VEGF-PLs) had been delivered into rat brain. Magnification bar: 50 μm. Color images available online at
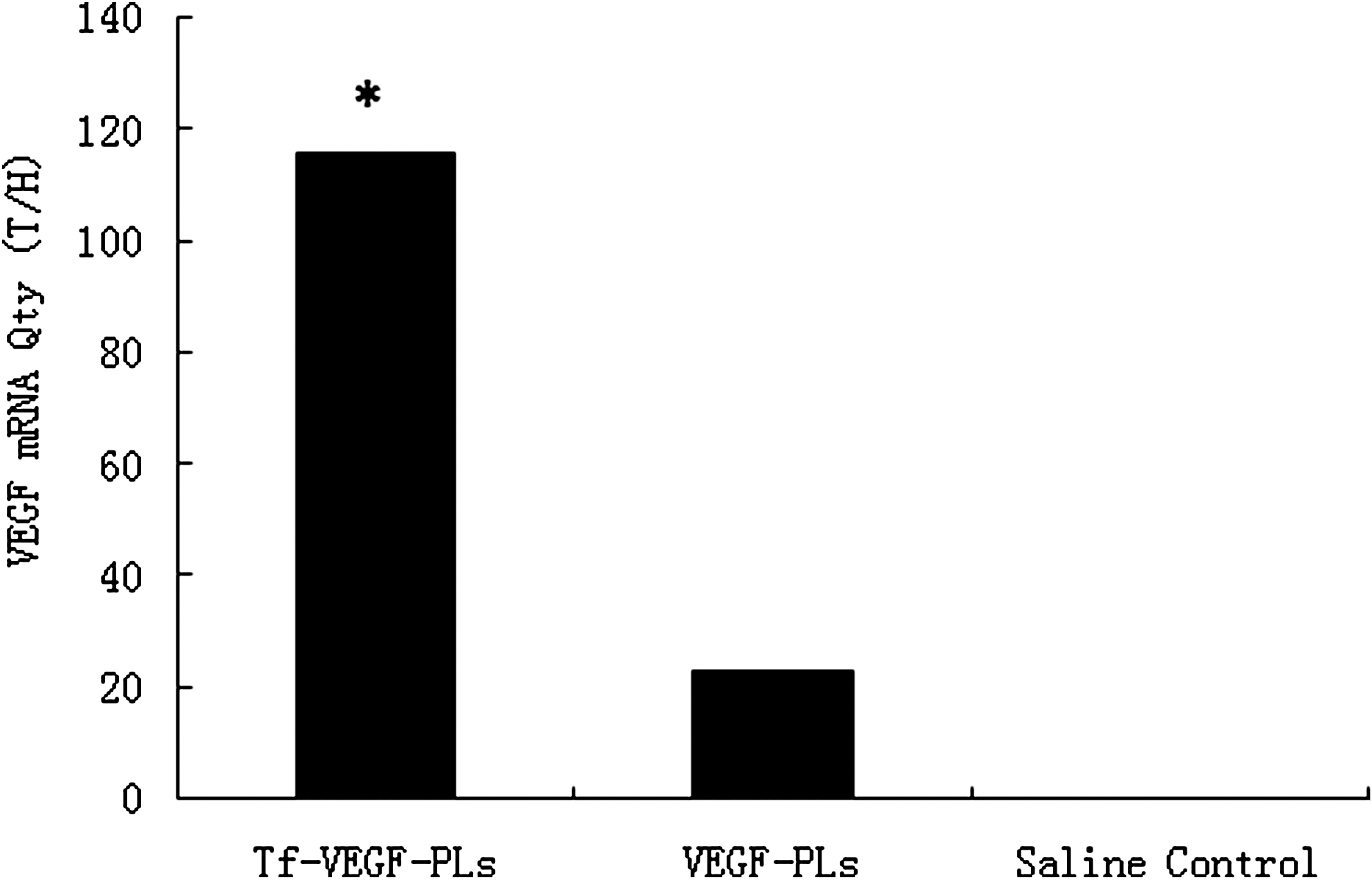
Relative quantities of vascular endothelial growth factor (VEGF) mRNA in the brain 24 hr after injection of transferrin-coupled PEGylated liposomes encapsulating vascular endothelial growth factor (Tf-VEGF-PLs), PEGylated liposomes encapsulating vascular endothelial growth factor (VEGF-PLs), and saline control. The VEGF mRNA level in ischemic brain treated with Tf-VEGF-PLs was significantly higher than that in the VEGF-PL and saline control groups (*p < 0.05) (n = 8 rats per group). T/H, concentration ratio of VEGF gene and Actin gene expression.
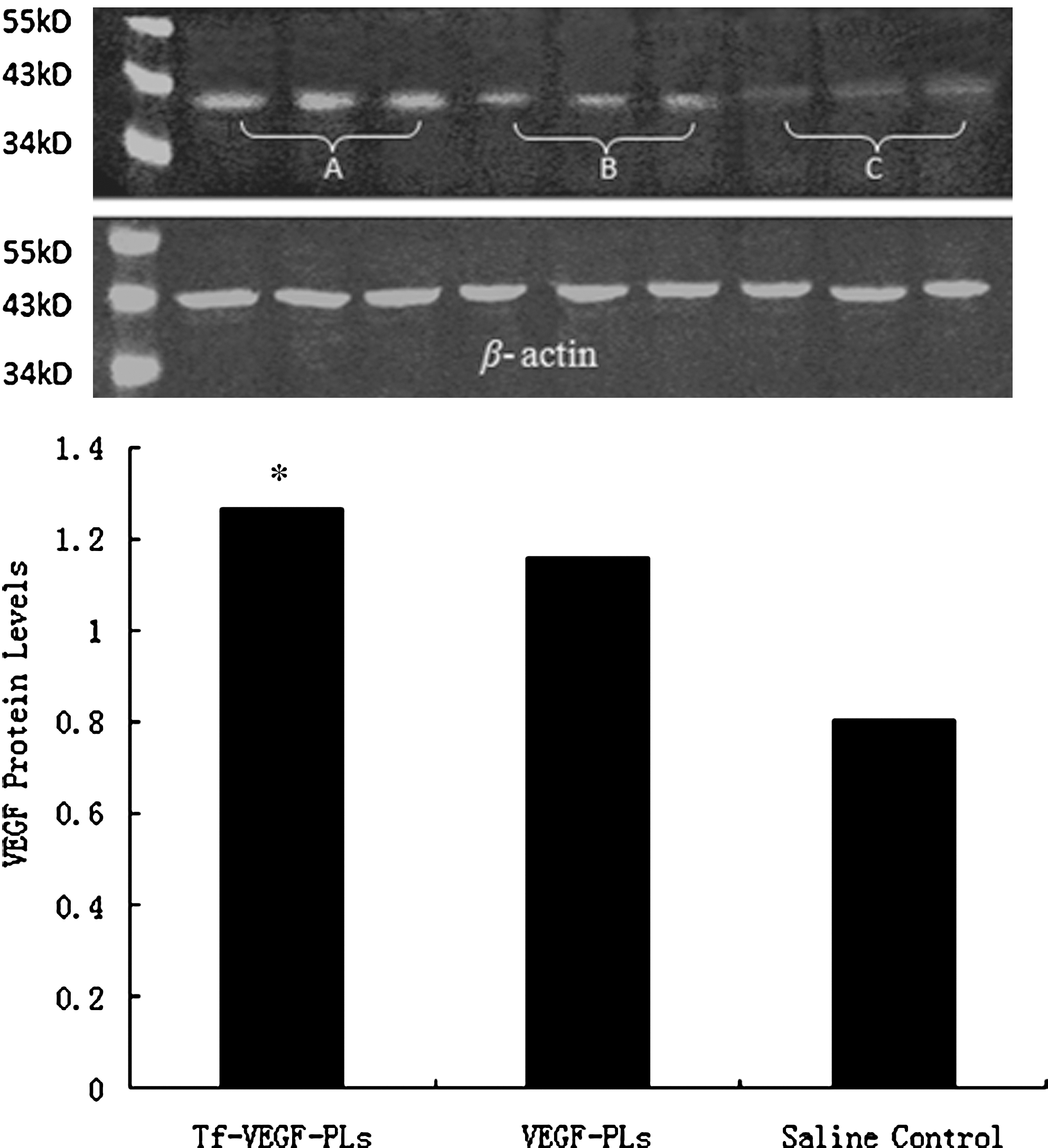
Expression of vascular endothelial growth factor (VEGF) protein in rat brain 48 hr after injection of transferrin-coupled PEGylated liposomes encapsulating vascular endothelial growth factor (Tf-VEGF-PLs), PEGylated liposomes encapsulating vascular endothelial growth factor (VEGF-PLs), and saline control; determined by Western blotting. VEGF protein in ischemic brain treated with Tf-VEGF-PLs was significantly higher than in the VEGF-PL and saline control groups (*p < 0.05) (n = 8 rats per group). (
Tf-VEGF-PL enhances neuroprotection after cerebral ischemia
Cerebral infarcts in Tf-VEGF-PL-treated rats were reduced in size, however, especially regarding the extent to which they involved the dorsomedial cortex in the region of the anterior cerebral artery–MCA border zone, or penumbra (Fig. 6). Sun and colleagues (2003) documented that the infarct volume at 3 days was reduced approximately 35% in VEGF-treated rats, and this function of VEGF persisted for at least 4 weeks. In our study, 21 days after drug injection, we observed a significant decrease in infarct volume in the Tf-VEGF-PL-treated group, compared with the VEGF-PL group. Over 21 days, rats that received Tf-VEGF-PLs displayed consistently better neurological function (Fig. 7). Neurological improvement in Tf-VEGF-PL-treated rats was attributed primarily to improvement in two motor tests (head movement on raising the rat by the tail and circling behavior toward the paretic side on placing the rat on the floor) and one sensory test (proprioception). In contrast, there was little difference between treated and untreated rats with respect to abnormal movements or reflex function.
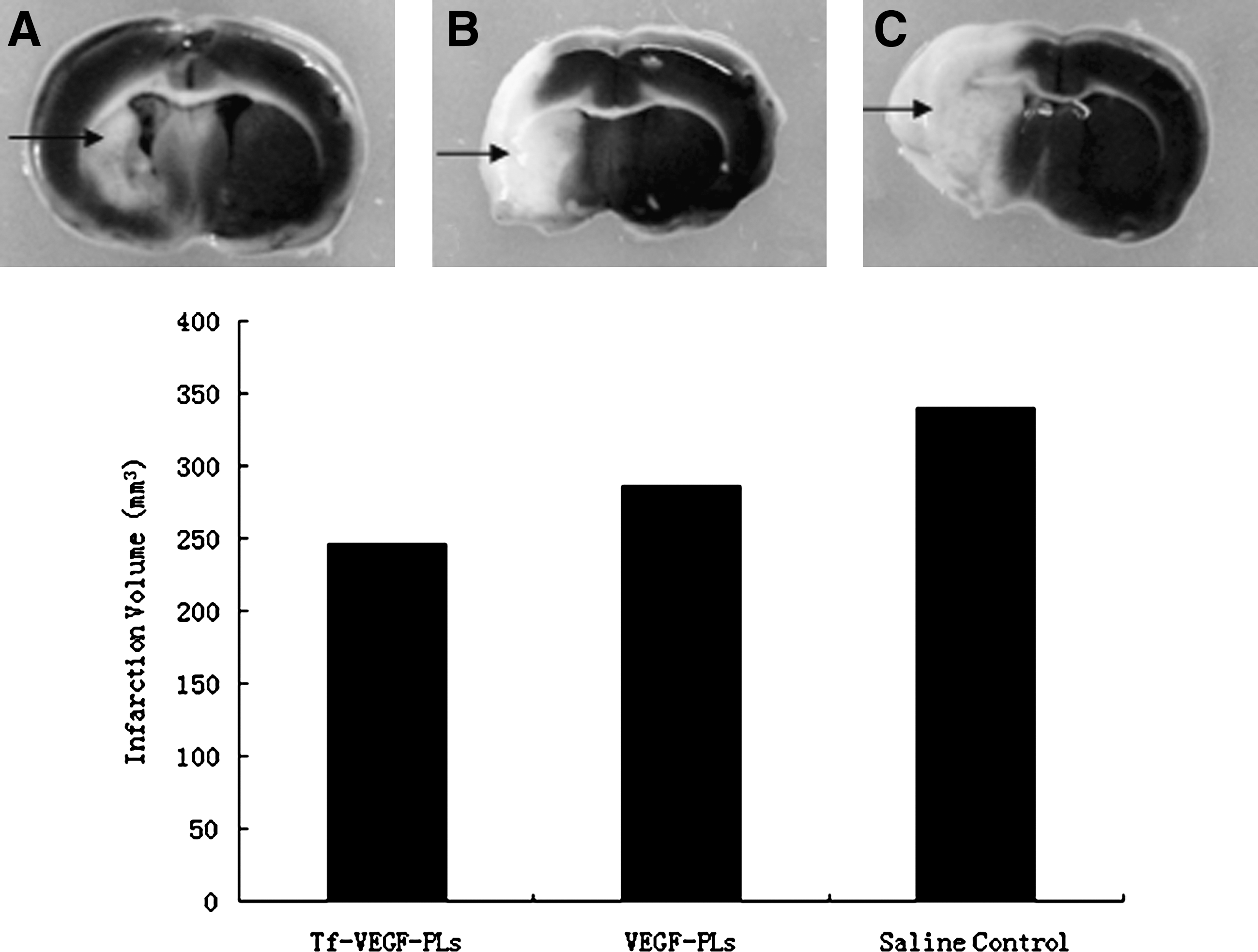
Effect of treatment with intravenous transferrin-coupled PEGylated liposomes encapsulating vascular endothelial growth factor (Tf-VEGF-PLs) on infarct size after middle cerebral artery occlusion (MCAO). Rats underwent MCAO and were treated with intravenous Tf-VEGF-PLs, VEGF-PLs, or saline control, and infarct size (arrows) was measured on brain sections stained of 2,3,5-triphenyltetrazolium chloride (TTC; white staining). The results are shown for (
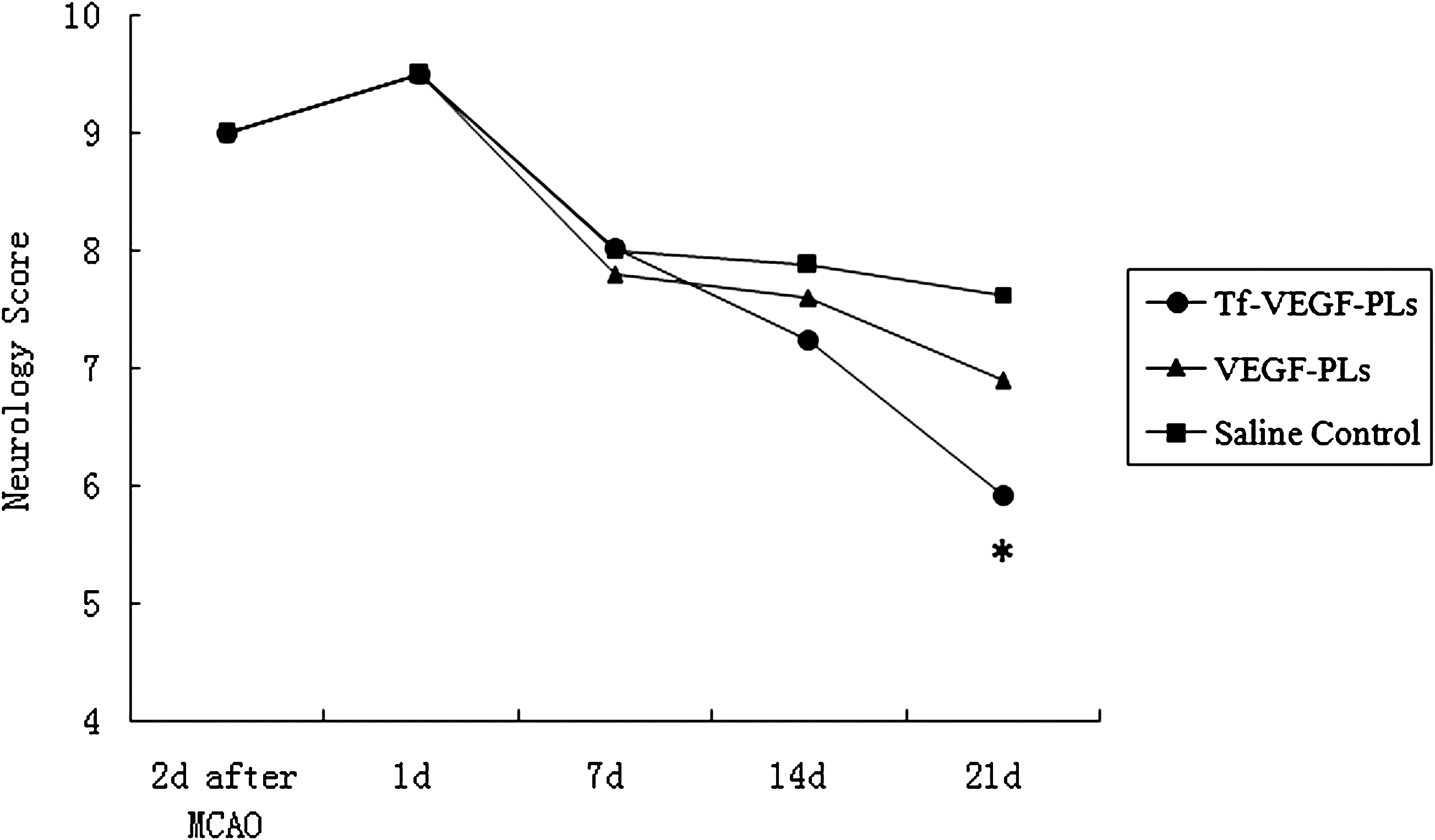
Effects of intravenous Tf-VEGF-PL treatment on neurological recovery after middle cerebral artery occlusion (MCAO). Rats underwent MCAO and were treated with intravenous Tf-VEGF-PLs, VEGF-PLs, or saline control. Similar neurological deficient rat models were chosen 2 days after MCAO. The neurological severity score (NSS) was not significantly different 1, 7, and 14 days after drug injection. On day 21, the NSS in the Tf-VEGF-PL-treated group was 8, which was significantly lower than in the VEGF-PL and saline control groups (7 and 7.5, respectively; *p < 0.05).
Tf-VEGF-PL promotes neovascularization
The VEGF gene has been shown to be a potent angiogenic factor required for cerebral angiogenesis (Zhang et al., 2000). We determined whether VEGF overexpression would induce neovascularization. FITC–dextran marking showed increased vascular density in the penumbra of Tf-VEGF-PL-treated hemispheres (245,873.9, number of microvessels per field) compared with that in VEGF-PL-treated hemispheres (139,801.3) or saline-treated hemispheres (102,175.5) (Fig. 8, p < 0.05). VEGF gene transduction also increased segment lengths and diameters. To determine whether the newly formed vessels were functional, we measured CBF by laser Doppler flowmetry. VEGF gene transfer in the Tf-VEGF-PL-treated group (35%) significantly enhanced vascular perfusion 21 days after drug injection compared with the VEGF-PL-treated group (29%) and the saline control group (21%) (Fig. 9, percentages of baseline values, p < 0.05).
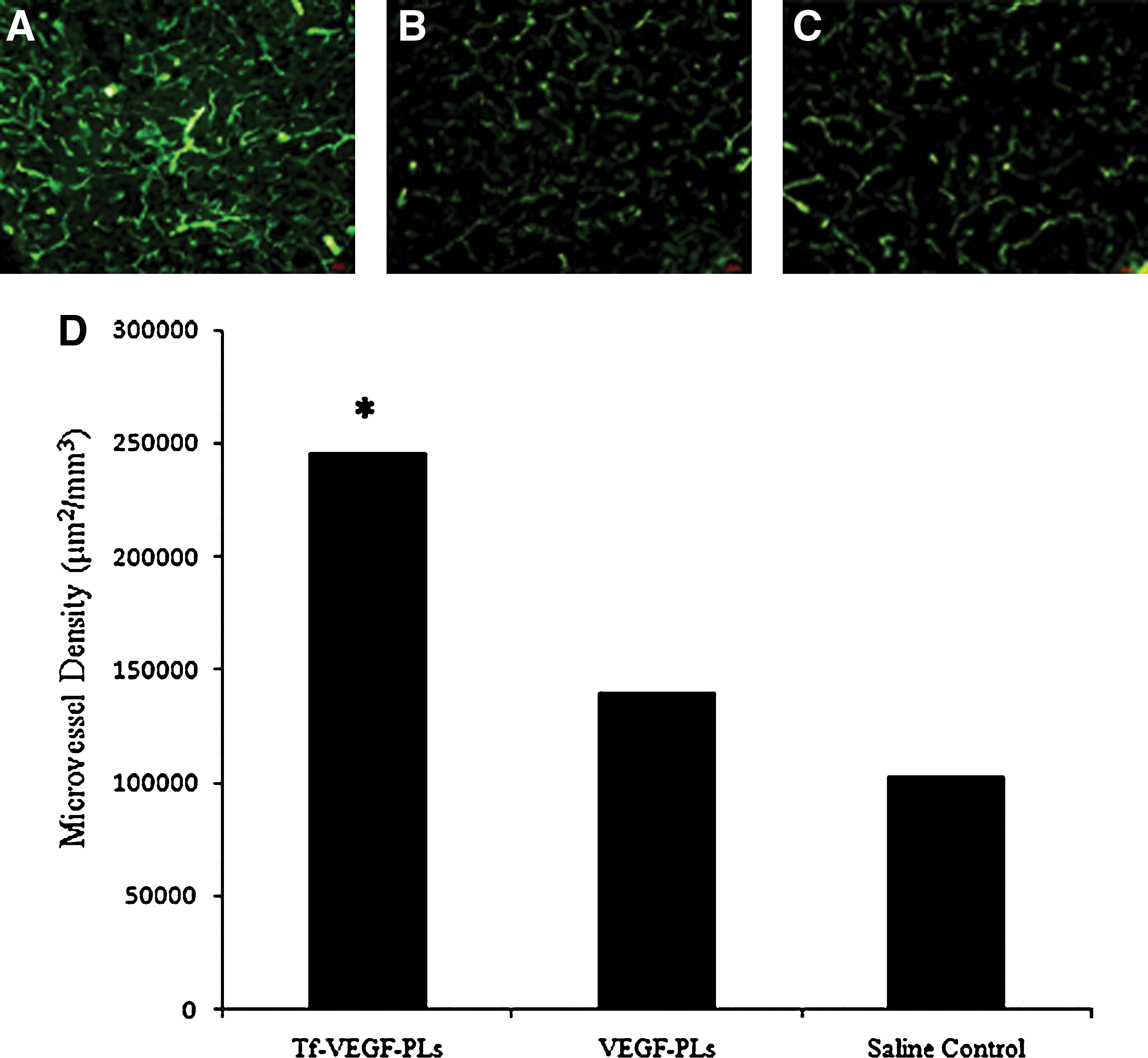
VEGF overexpression induces angiogenesis. (
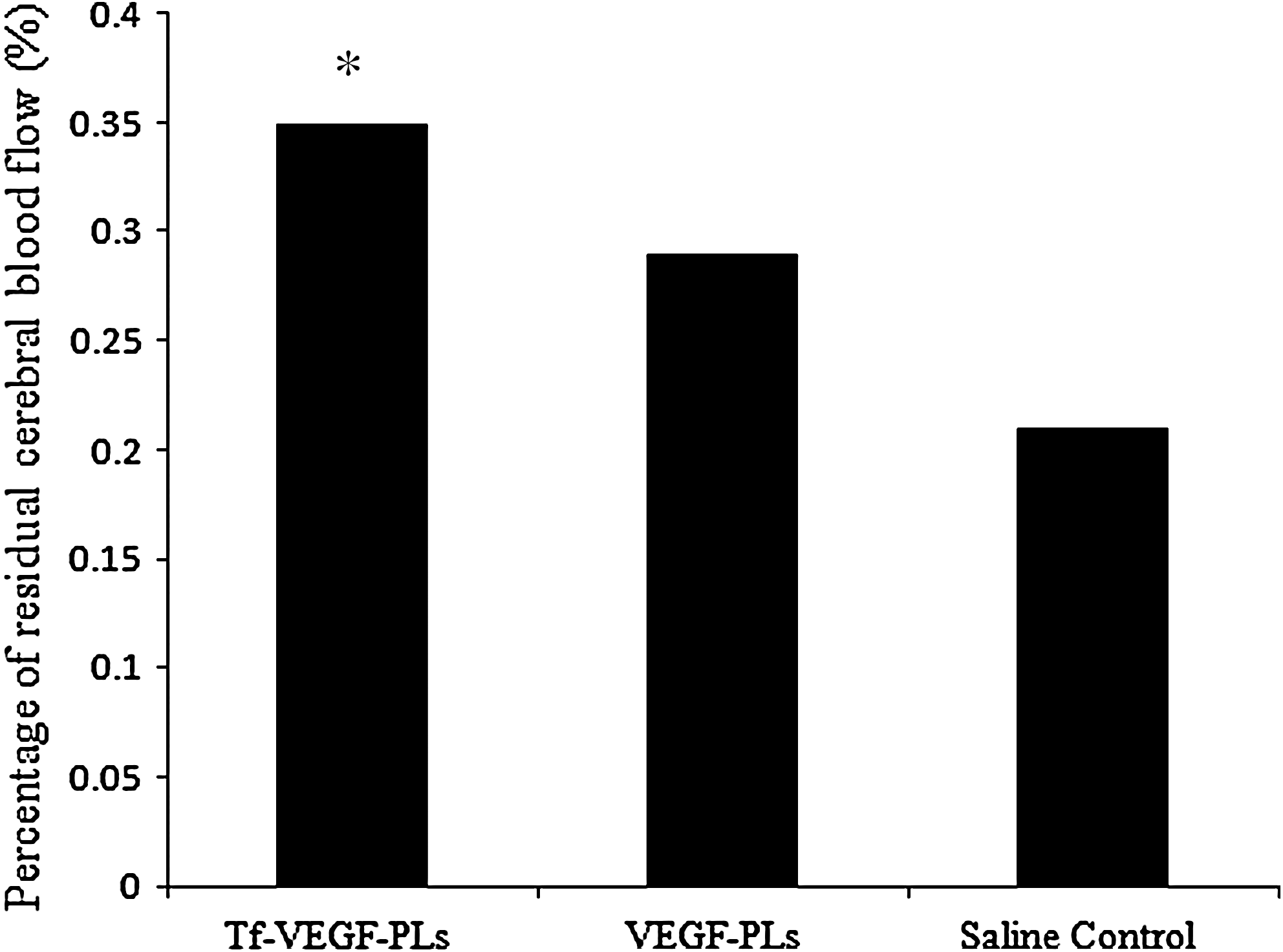
Percentage of residual cerebral blood flow. *p < 0.01 versus VEGF-PLs and saline control, n = 8 for each group.
Discussion
This study indicates that the administration of Tf-VEGF-PLs in a nonintracranial way reduces infarct size, enhances neuroprotection, and promotes cerebral angiogenesis and neurogenesis in ischemic rats undergoing MCAO. Of particular note, these effects were observed with (1) venous administration of PLs, which could cross the BBB to deliver VEGF gene into brain; and with (2) delayed administration of VEGF, which was given beginning on day 2 after MCAO. Hermann and Zechariah (2009) summarized 22 animal studies investigating the therapeutic effects of VEGF. These studies adopted recombinant VEGF, VEGF-secreting cells, AAV vectors, and plasmids carrying VEGF cDNA to evaluate VEGF the mechanism of action, but none of studies involved liposomes. In our study, liposome was chosen as the basic vector and further modified with PEG and Tf to form a brain-targeting drug carrier, and successfully carried the exogenous gene into brain.
It is well known that particle size plays an important role in the alteration of pharmacokinetics by affecting tissue distribution and clearance. Vehicles with small particle size (<200 nm) are known to increase the accumulation of drug in the tissue via an enhanced permeability and retention (EPR) effect (Moreira et al., 2001; Xu et al., 2005). In our study, the average liposome particle size was 100 nm, ensuring accumulation of the drug. In addition, aggregation of liposomes in the bloodstream can accelerate the uptake of liposomes by the reticuloendothelial system (RES). In our study, however, liposome aggregation did not occur in any of three solutions (PBS, PBS including 3% Tween 80, or a solution composed of equal volumes of FBS and PBS at 37°C). This suggested that these liposomes would be stable and not aggregate in the bloodstream, thereby decreasing their elimination from the body by the RES.
Tf is considered to possess great potential as a brain-targeting ligand (Li and Qian, 2002). The Tf receptor is expressed both on the BBB and neuronal cell membrane (Schlachetzki et al., 2004). The transvascular delivery of genes is possible by accessing Tf-conjugated transport systems, which might cross the BBB and deliver genes to the doorstep of every neuron in the brain (Shi and Pardridge, 2000; Zhang et al., 2003; Schlachetzki et al., 2004). However, little information is presently available about the application of Tf as a ligand for gene delivery to the brain in vivo. In our study, PEG served as a spacer in Tf-modified conjugates. The conjugation of Tf at the tip of the PEG tail profoundly increased the targeting capability of Tf-VEGF-PLs to the brain.
There are two main barriers to in vivo gene targeting to organs, such as the brain, that have continuous capillaries of restricted permeability: the microvascular endothelial barrier and the plasma membrane barrier of the target cells (Shi and Pardridge, 2000). The presence of the two barriers profoundly limits the expression of exogenous genes after transvascular administration. It was reported that exogenous gene expression could be observed in limited regions around the cerebral ventricles because of their relatively weak BBB (Sako et al., 2005). In our study, the conjugation of Tf with PLs greatly increased gene expression in the brain. The high expression of the Tf receptor at both barriers (the BBB and the neuronal plasma membrane) enables the widespread expression of an exogenous VEGF gene encapsulated in Tf-PLs after intravenous administration. This is in harmony with the fact that OX26 PEGylated immunoliposomes exhibited extended gene expression in brain via intravenous administration (Shi and Pardridge, 2000). Omori and colleagues (2003) intravenously administered PEG-liposomes conjugated with Tf to rats after 90 min of transient middle cerebral occlusion, and their results showed that Tf-PEG fluorescence was remarkably increased, with a peak 2 days after injection. This was in concordance with our results, that is, that high expression of VEGF in ischemic brain was recorded 48 hr after Tf-VEGF-PL administration and guaranteed the function of VEGF in brain.
Our previous study showed that VEGF transferred with liposomes into cultured C6 glioma cells in vitro could achieve higher VEGF expression at 48 hr, and demonstrated the feasibility of choosing the glial cell as the target cell for gene transfer (Wang et al., 2000). The role of VEGF in ischemic and hypoxic conditions has been confirmed by experiments performed in vitro. It has been observed that VEGF protects primary cultures of rat cortical neurons from hypoxia and glucose deprivation. All of these studies represent a strong foundation for applying Tf-VEGF-PLs in the gene therapy of cerebral ischemia in rats.
In our study, we chose delayed administration of VEGF, which was given beginning on day 2 after MCAO. This regimen was adopted because early VEGF treatment promoted brain edema, hemorrhagic transformation of the infarct, and spread of the ischemic lesion (Zhang et al., 2000). Chu and colleagues (2005) used a postacute delivering strategy, in which VEGF was applied starting 2 days after the stroke. The results showed an unequivocally beneficial effect of VEGF after systemic administration. This suggests that the time of administration, in addition to route of delivery, might have a role in the actions of VEGF in the ischemic brain. That delayed treatment of cerebral ischemia may enhance neuroprotection has been reported in several papers; Sun and colleagues (2003) reported that VEGF produced a strikingly delayed effect on neurogenesis, which was apparent only after 28 days.
We observed a significant reduction in infarct volume at 21 days with Tf-VEGF-PLs, suggesting that most of the ischemic penumbra was rescued because of the direct effect of VEGF. It is possible that the infarcted tissue could have been cleared by phagocytes, an important process of brain repair after stroke (Kitagawa et al., 2001), which is facilitated by the process of neovascularization. Although further studies are needed to completely understand the mechanism of VEGF, we found that VEGF is effective in achieving the overall goal of neurological recovery and increasing survival of animals (Fig. 6). Rats treated with Tf-VEGF-PLs in our study regained almost all vital neurological functions by 21 days, with the exception of slight impairment in forelimb movement and paresis of the eye muscles on the ipsilateral side. In our study we also found that FITC–dextran-marked microvessels showed increased vascular density in the penumbra of Tf-VEGF-PL-treated hemispheres (Fig. 7), and the result of LDF demonstrated that increased vascular density enhanced perfusion of the infarcted side of the brain.
In conclusion, brain-specific expression of an exogenous gene is possible with the combined use of a brain-targeting ligand and a cationic gene delivery system such as liposomes stabilized by PEGylation. In addition to successful brain transfer of VEGF, intravenous administration of VEGF-loaded Tf-PLs also achieved high VEGF expression in the ischemic brain, enhanced neuroprotection, and promoted neovascularization. Although the mechanisms of this efficient gene delivery vector are not fully understood and are under investigation, we believe that Tf-VEGF-PLs hold great promise for efficient, noninvasive, and brain-targeting gene delivery.
Footnotes
Acknowledgments
This study was supported by a grant from the National Natural Science Foundation of China (grants 30571901 and 30772227) and the National High Technology Research Project (2006AA02A115). The authors greatly appreciate the work of Prof. Xin-ru Li (Department of Pharmaceutics, Peking University School of Pharmaceutical Sciences, Beijing, China), who prepared the liposomes and immunoliposomes.
Author Disclosure Statement
No competing financial interests exist.