
Abstract
Various plasmids were delivered into rodent limb muscles by hydrodynamic limb vein (HLV) injection of naked plasmid DNA (pDNA). Some of the pDNA preparations caused significant muscle necrosis and associated muscle regeneration 3 to 4 days after the injection whereas others caused no muscle damage. Occurrence of muscle damage was independent of plasmid sequence, size, and encoded genes. It was batch dependent and correlated with the quantity of bacterial genomic DNA (gDNA) that copurified with the pDNA. To determine whether such an effect was due to bacterial DNA or simply to fragmented DNA, mice were treated by HLV injection with sheared bacterial or murine gDNA. As little as 20 μg of the large fragments of bacterial gDNA caused muscle damage that morphologically resembled damage caused by the toxic pDNA preparations, whereas murine gDNA caused no damage even at a 10-fold higher dose. Toxicity from the bacterial gDNA was not due to endotoxin and was eliminated by DNase digestion. We conclude that pDNA itself does not cause muscle damage and that purification methods for the preparation of therapeutic pDNA should be optimized for removal of bacterial gDNA.
Introduction
During the course of more recent preclinical studies to treat muscular dystrophy with dystrophin-expressing pDNA, we were surprised to see significant muscle damage in rodents 3 days after HLV injection of pDNA because we also observed long-term expression from genes encoded in the pDNA (Zhang et al., 2010). Plasmid DNA had been reported to cause inflammation-induced damage and regeneration of skeletal muscles after direct intramuscular injection (McMahon et al., 1998) or electroporation of the naked pDNA into muscle (Hartikka et al., 2001; Durieux et al., 2004). Muscle damage correlated with the dose of pDNA delivered (Durieux et al., 2004) and in one report was reduced by methylation of the CpG sequences in the plasmid (McMahon et al., 1998).
Preparations of pDNA can contain fragments of bacterial genomic DNA (gDNA) that, depending on the type of pDNA production technology, copurify with the pDNA. We show here that the occurrence of substantial muscle damage after HLV injection was pDNA batch dependent and not an intrinsic characteristic of the pDNA, even though the plasmids all contained abundant CpG dinucleotides in the plasmid backbone. Toxicity of pDNA batches correlated strongly with the quantity of contaminating bacterial gDNA in them.
Materials and Methods
Animals
Outbred ICR mice, inbred C57BL/6 mice, and Sprague-Dawley rats were obtained from Harlan Laboratories (Indianapolis, IN). Male mice and female rats were used. Studies were conducted in accordance with the guidelines set forth by the Roche Madison (formerly Mirus Bio) Animal Care and Use Committees and by the Guide for the Care and Use of Laboratory Animals (National Institutes of Health, Bethesda, MD).
Hydrodynamic limb vein injection procedure
Mice were anesthetized with 1–2% isoflurane (Abbott Laboratories, North Chicago, IL). They were shaved and prepared for surgery and a small latex tourniquet was wrapped tightly around the upper hind limb (above the quadriceps) and secured in place with a hemostat. A small incision was made in the skin between the ankle and the knee to expose a segment of the distal great saphenous vein. A 30-gauge needle catheter (made by our laboratory as described in Zhang et al., 2004) was then inserted into the distal great saphenous vein of the mouse, advanced about 0.5 cm, and held in place during the injection. The catheter was connected to a Harvard PHD 2000 syringe pump (Harvard Apparatus, Holliston, MA) and 1 ml of DNA–saline solution containing 0.5 mg of pDNA was injected at a rate of 8 ml/min. In some animals the contralateral limb was injected with saline alone as a control. The tourniquet was removed approximately 2 min after injection and the skin was closed with 4-0 absorbable suture. The total tourniquet time was roughly 3 min and each procedure took 10 min to complete. The procedure was the same for rats except that a 25-gauge catheter was used and 2.7 mg of pDNA in a total volume of 3.5 ml of pDNA–saline solution was injected at a rate of 10 ml/min.
Plasmid DNA
Plasmid descriptions are in Table 1. Nonexpressing plasmids pUK21 and pMIR0266 (Zhang et al., 2010) contained no mammalian promoter. Promoters in plasmid constructs included the human major immediate-early cytomegalovirus (CMV) enhancer/promoter and the human desmin promoter (called DCR and described in Zhang et al., 2004). Coding sequences were canine, mouse, or human dystrophin; codon-optimized luciferase (Promega, Madison, WI); β-galactosidase; or CpG-free murine secreted alkaline phosphatase (mSEAP; InvivoGen, San Diego, CA). All plasmids contained the pUC-based ColE1 origin of replication. All plasmids were kanamycin resistant except pCI-Luc-17.1, pCI-Luc, and pCI-LacZ, which were constructed in the pCI mammalian expression vector (Promega); these were ampicillin resistant and previously described (Zhang et al., 2004). The CMV-luciferase expression cassette was the same in pCI-Luc-17.1, pCI-Luc, and pMIR0048. All preparations of pDNA contained less than 100 EU of endotoxin per milligram and most had at least one magnitude less (Table 1). Preparations of pDNA were commercially prepared by Aldevron (Fargo, ND) or PlasmidFactory (Bielefeld, Germany), using various procedures to yield pDNA with supercoiling, endotoxin, and bacterial genomic DNA levels indicated in Table 1.
CMV, cytomegalovirus; EU, endotoxin unit; N/D, not determined; SEAP, secreted alkaline phosphatase.
Supercoiling was determined by capillary gel electrophoresis at PlasmidFactory as previously described (Schleef and Blaesen, 2009).
The quantity of E. coli genomic DNA in the preparation is shown as a percentage of the total DNA.
Damage was quantitated by morphometry as the mean area ± SD for all evaluated muscle specimens per animal that contained damaged, necrotic, and/or regenerating muscle, n = 3–11. For a few of the pDNA batches the muscle damage was estimated (est.) from microscopic inspection of histology samples without morphometric quantitation.
Only one mouse was tested with this batch.
Endotoxin and genomic DNA
Endotoxin (lipopolysaccharide, product L4516; Sigma-Aldrich, St. Louis, MO) was resuspended in sterile 0.9% saline solution (Phoenix Pharmaceuticals, Burlingame, CA) at 1 mg/ml, which was equal to 500,000 EU/mg, and was further diluted in saline. Bacterial genomic DNA (gDNA) was either isolated from Escherichia coli DH10B with a genomic DNA purification kit (Gentra Systems, Minneapolis, MN), or purchased from USB (Cleveland, OH) (product 14380). The Gentra Systems purification kit included an RNase A treatment to degrade RNA. Mammalian genomic DNA was purified from the liver of an ICR mouse, using the same kit. Both preparations were then treated for endotoxin removal, using a MiraCLEAN kit (Mirus Bio, Madison, WI), followed by ethanol precipitation. The DNA was resuspended in 0.9% NaCl (normal saline). It was sheared by passage through a 30-gauge needle twice. Before shearing, the bacterial and mouse genomic DNA were both primarily high molecular weight fragments larger than 23 kb. The sheared fragments ranged in size from approximately 1 kb to very large (remaining in the well when run on a 0.8% agarose gel); the majority of fragments migrated with DNA markers of 9–20 kb. To denature, DNA samples were heated for 10 min at 95°C and then snap cooled on ice. The purchased bacterial gDNA was extracted twice with phenol–chloroform–isoamyl alcohol (25:24:1), ethanol precipitated, resuspended, extracted twice for endotoxin removal, ethanol precipitated, and resuspended in TE (10 mM Tris-HCl, 0.1 mM EDTA; pH 8). It was then sheared by passage through a 30-gauge needle five times. TE was replaced with saline, using a Centriprep YM-10 for filtration (Millipore, Billerica, MA). The gDNA fragments ranged in size from approximately 1 kb to very large; the majority were about 9 kb. Digested gDNA was prepared from the sheared gDNA in TE with 100 U of DNase I per milligram of DNA in 10 mM Tris-HCl, 2.5 mM MgCl2, and 0.5 mM CaCl2 and incubated at 37°C overnight. DNase was removed from the digested gDNA by centrifugation through a Centriprep YM-10 column. The filtration device allows passage of double-stranded DNA (dsDNA) fragments of less than 20 bp and single-stranded DNA (ssDNA) fragments of less than 30 bases. Thorough digestion was confirmed by absorbance at 260 nm and by electrophoresis showing no detectable fragments on an ethidium bromide-stained agarose gel. Endotoxin levels for the sheared and digested E. coli gDNA were less than 0.4 EU/mg DNA. The recovered digested gDNA was diluted in saline before HLV injection.
Fragmented pDNA
The pUK21 preparation described in Table 1 was digested separately with EcoRI, BamHI, AvaI plus XhoI, or HaeII plus NdeI to result in fragments of the indicated sizes: 3090 bp (EcoRI digest); 3090 bp (BamHI); 1673, 771, 617, and 29 bp (AvaI plus XhoI); and 2006, 370, 316, 245, 145, and 8 bp (HaeII plus NdeI). Equal molar amounts of fragments from each digest were combined. This mixture was phenol extracted twice, chloroform extracted, ethanol precipitated, and then treated for endotoxin removal with MiraCLEAN in accordance with the manufacturer's protocol, ethanol precipitated again, and finally resuspended in saline. For comparison with fragmented pDNA, supercoiled pUK21 was subjected to the same purification steps before injection into mice.
Real-time quantitative polymerase chain reaction of Escherichia coli genomic DNA
Amplification and data analysis were performed with a 7500 Fast real-time PCR system and Fast SYBR green PCR master mix (Applied Biosystems, Foster City, CA), according to the manufacturer's instructions. PCR primers were designed to amplify a 128-bp fragment of the single-copy 1-deoxy-
Morphological analysis and morphometry
Rodents were anesthetized with 3 to 5% isoflurane and killed. The skin was then removed from the hind limb to expose the muscles. The medial group of muscles of the upper leg, which includes the gracilis and adductor muscles and comprises a total of 24% of the hind leg muscle, and the posterior group of muscles of the lower leg, which includes the triceps surae: gastrocnemius, soleus, and plantaris and comprises 20% of the leg muscle, were evaluated for histology. Each of these groups of muscles was cut into two parts and immediately placed in a 10% buffered formalin solution. Specimens were then further processed, embedded in paraffin, sectioned, and stained with hematoxylin and eosin (H&E). The four pieces of muscle were each sectioned at three or occasionally at two levels. The levels were 0.5–1 mm apart. Two cross-sections were taken at each level. In most cases, this resulted in 24 muscle specimens sampled from a broad area of the highly transfected muscles of each injected hind leg. These sections were evaluated by a pathologist blinded to sample identity. Morphometry (SigmaScan Pro imaging program; Systat Software, Chicago, IL) was conducted to quantitate the area of damage. For this, slides of H&E-stained cross-sections of muscle specimens were scanned and digitized with a photographic film scanner. Muscle damage was determined by the pathologist (V.S.) viewing each slide under a microscope. The corresponding whole slide-scanned image was then opened with SigmaScan Pro software and enlarged to the size of a computer screen so that the damaged areas could be outlined. The total area of the muscle specimens and the damaged areas were quantitated with this software to determine the percentage of damaged and regenerating muscle (see Fig. 2).
Statistical analysis
Correlation between muscle damage and bacterial gDNA content and between muscle damage and endotoxin content was determined with two-tailed p values from XY tables, using the Spearman correlation coefficient. Contingency analysis (chi square) was performed with Fisher's exact test. Prism software (GraphPad, San Diego, CA) was used for these analyses.
Results
Batch-dependent muscle damage from plasmid DNA
Rodents were given one HLV injection of cytomegalovirus (CMV) promoter-driven human dystrophin-expressing pDNA (CMV-hDys) or empty vector pUC19 in the hind leg to determine whether any adverse effects of the foreign human dystrophin expression would be observed (Fig. 1). Rats were injected with 2.7 mg of pDNA. ICR and C57BL/6 mice were injected with 0.5 mg of pDNA. Some contralateral limbs were injected with saline as a control (Fig. 1b). CMV promoter-driven genes encoded in pDNA typically reach maximal expression 7 days after naked pDNA delivery to muscle (Zhang et al., 2001; and data not shown), and therefore animals were killed 3, 4, 5, 7, 14, and 21 days after pDNA injection. Muscle sections were taken from large muscle groups of the leg that transfect well: the medial muscles of the thigh, including the biceps, and the posterior muscles of the lower leg, including the gastrocnemius. These muscle specimens were evaluated for evidence of pathology by histological examination. The limb muscles of rats and mice that were killed 3 or 4 days after HLV injection of either type of plasmid had significant muscle damage (Fig. 1c, f, and g). More than 50% of some localized areas of the muscle specimens appeared to be damaged and/or undergoing muscle regeneration, as evidenced by necrotic myofibers, an infiltration of mononuclear cells (Fig. 1k), and fusion of satellite cells into myofibers (Fig. 1j). Muscle damage was much lower on day 5, and by day 7 the muscles of pDNA-injected mice and rats appeared to be nearly normal except for central nuclei in the muscle fibers, which indicated the muscle had recently regenerated (data not shown). Muscles from rodents injected with either CMV-hDys or pUC19 and evaluated on day 14 or 21 appeared normal except for centralized nuclei (Fig. 1d and e; and data not shown). Muscle damage followed the same time course in mice and in rats, regardless of whether the injected pDNA was an empty vector or if it expressed human dystrophin. The apparent muscle toxicity did not follow the expected time course of plasmid-encoded gene expression. Muscle damage did not correlate with an elevation in serum creatine kinase levels because the volume of muscle damage was insufficient to raise serum levels (data not shown).
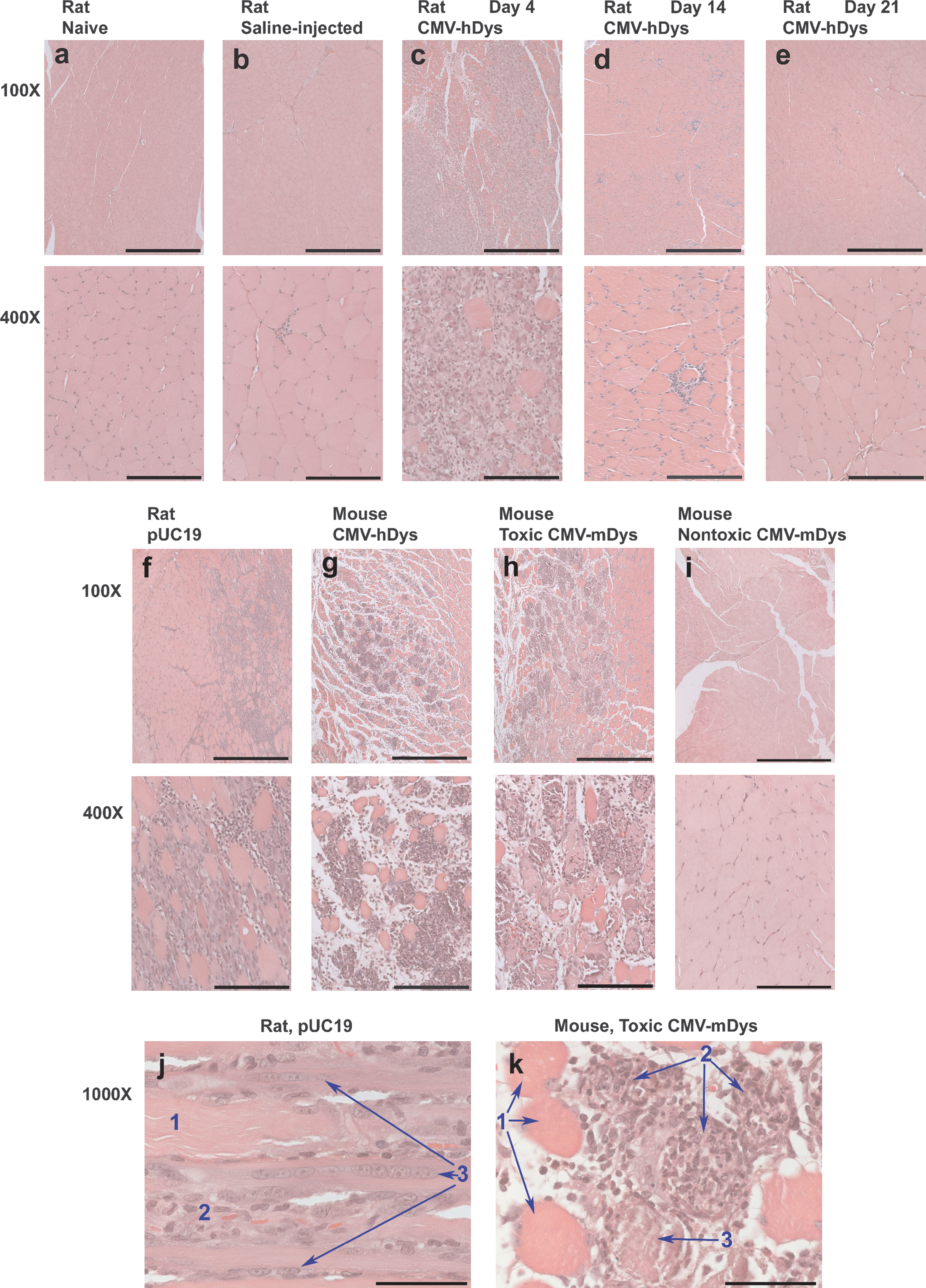
Muscle damage after hydrodynamic limb vein (HLV) injection of some batches of plasmid DNA (pDNA) into rodents. Muscle sections were taken from the posterior lower leg including the gastrocnemius, processed for histology, and were stained with hematoxylin and eosin (H&E). Rat muscle specimens were taken from (
We then tested more than 26 pDNA preparations to determine whether all caused muscle damage 3 days after HLV injection. In most cases, muscle damage was quantitated by morphometry from 16 to 24 muscle specimens per animal (Fig. 2). Outbred ICR mice consistently had more muscle damage than the inbred C57BL/6 mice (data not shown). Sprague-Dawley rats had similar levels of damage as ICR mice. Therefore, ICR mice were used to test all batches of plasmids for associated toxicity (Table 1). HLV injection of saline alone caused negligible muscle damage (Fig. 1b and Fig. 3a), with an average of 1.2 ± 1.6% of the muscle fibers in ICR mice showing damage. Muscle damage was occasionally higher because the injection pressure is difficult to control in mice on account of the tourniquet being tightened by hand. HLV injection of some preparations of pDNA caused significant muscle damage, but injection of other preparations of even the same plasmid construct caused no more damage than injection of saline alone (Fig. 1h and i, Fig. 3, and Table 1). Multiple preparations of pDNA expressing mouse dystrophin from two different constructs caused highly variable amounts of muscle damage in mice (Fig. 1h and i, Fig. 3a and b, and Table 1). Some batches caused damage to an average of more than 20% of all the muscle specimens from some limbs with localized areas in which nearly all muscles were damaged, whereas other batches caused no damage. The pDNA batches that caused no damage contained either no expression cassette or expressed mouse dystrophin, luciferase, mouse secreted alkaline phosphatase (mSEAP), or β-galactosidase. They ranged in size from 2.3 kilobase pairs (kb) to 21.8 kb. Most contained a kanamycin resistance gene for drug selection during bacterial growth, but some contained the ampicillin resistance gene. All constructs had a pUC-based plasmid backbone and origin of replication.

Morphometric quantitation of muscle damage. Muscle sections were taken from the gastrocnemius, processed for histology, and stained with H&E. (
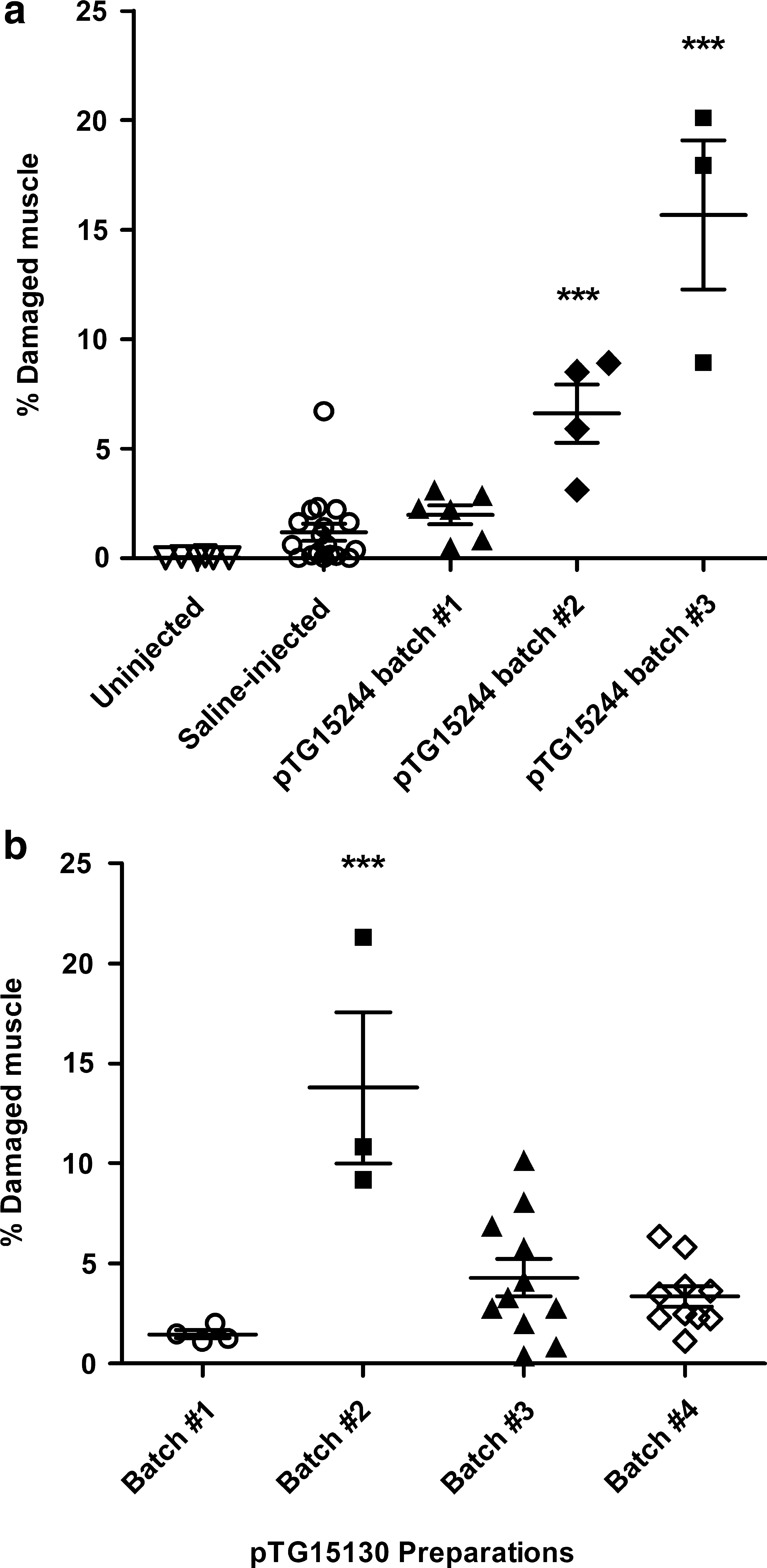
Batch-dependent muscle damage after HLV delivery of pDNA. (
Endotoxin contamination and muscle damage
A number of components of the pDNA preparation were considered as the source of toxicity, beginning with endotoxin. All preparations had lower levels of endotoxin than the common manufacturers' cutoff of 100 EU/mg DNA (Table 1). The endotoxin levels in the batches of pDNA that were tested varied from less than 0.005 EU/mg DNA to more than 34 EU/mg DNA. Muscle damage and endotoxin levels did not correlate significantly (n = 115 mice; Spearman correlation coefficient r = 0.05, p = 0.57), although nontoxic pDNA preparations generally had low endotoxin levels. The effects of endotoxin on the muscle were directly evaluated by HLV injection of 0.5, 50, or 500 EU of lipopolysaccharide endotoxin in saline. Day 3 muscle specimens from mice given 50 EU of endotoxin in an HLV injection had the same appearance as muscle from mice injected with saline alone (data not shown). HLV injection of 500 EU of endotoxin (1 μg) resulted in minimal infiltration in the interstitial space, but otherwise the muscle had a similar appearance to that of mice given an HLV injection of saline (data not shown). Therefore, endotoxin levels do not account for the observed batch-dependent muscle damage.
Bacterial genomic DNA contamination and muscle damage
The amounts of contaminating Escherichia coli genomic DNA (gDNA) present in the pDNA preparations were determined by quantitative PCR (Table 1). Muscle damage correlated strongly with bacterial gDNA content in the pDNA preparations (Fig. 4a). This correlation was evident whether mice were evaluated as individuals (Fig. 4a) or on the basis of the group means of mice injected with the same pDNA batch (Fig. 4b). Evaluating all mice together with each individual as one data point increased the statistical power of the correlation.

Muscle damage correlates with bacterial genomic DNA (gDNA). ICR mice were given one HLV injection of 400–500 μg of pDNA. Plasmids expressed no gene; mouse, canine, or human dystrophin; luciferase; β-galactosidase; or mouse secreted alkaline phosphatase (SEAP). (
Preparations of pDNA with less than 1.3% bacterial gDNA administered by HLV injection into mice, or 6.5 μg of gDNA in a 500-μg dose of pDNA, rarely caused muscle damage (p < 0.0001). Of 70 mice injected with pDNA containing less than 7 μg of bacterial gDNA, 60 (86%) had no significant muscle damage. Most of the exceptions were from one batch of pDNA. In contrast, 31 of 45 mice (69%) injected with 14 μg or more of bacterial gDNA had more than 3% muscle damage, thus significantly more than mice given an HLV injection of saline alone. Mice injected with saline had 1–3% muscle damage (Fig. 3a); therefore, we define a toxic batch of pDNA as one that causes more than an average of 3% muscle damage.
To confirm that bacterial gDNA caused muscle damage and to investigate any effects of the DNA structure, we tested sheared, double-stranded and single-stranded (heat-denatured) fragments of bacterial gDNA, and compared these with sheared, double-stranded, and single-stranded mammalian gDNA (Fig. 5). The allowable level of bacterial gDNA in commercial pDNA preparations (5%) would be 25 μg in a 0.5-mg dose of injected pDNA. Injection of a higher dose of double-stranded bacterial gDNA, 200 μg, caused muscle damage in an estimated average of 5–15% of the cross-sectional area of all the evaluated hind leg muscles (posterior lower leg muscles and medial thigh muscles) from three mice. The 20-μg dose caused damage in an estimated average of 1–10% of the cross-sectional area of all the specimens from the mice. A 200-μg dose of heat-denatured bacterial gDNA caused approximately 5-fold less damage than the same dose of double-stranded gDNA, an average of less than 1–5% of the cross-sectional area of all specimens from three mice. The morphological appearance of the damaged muscle was the same in mice injected with bacterial gDNA or with toxic batches of pDNA. In contrast, mice injected with the mammalian gDNA all had approximately 1–3% damaged muscle in the cross-sectional area, similar to injection of saline alone, regardless of the dose and whether the DNA was double-stranded or single-stranded (a total of 11 mice). Muscle damage was associated with the bacterial origin of the gDNA; it was not simply the result of double-stranded or single-stranded DNA fragments.

HLV injection of mammalian compared with bacterial genomic DNA fragments. ICR mice were injected via the hind leg with 200 or 20 μg of sheared bacterial or mammalian gDNA that was either double-stranded or heat-denatured to be single-stranded. Mice were killed 3 days after injection and muscles of the posterior lower leg and medial upper leg were evaluated by histological examination at two levels; thus, a total of eight muscle specimens from one hind leg of each mouse was examined. Each column represents the average percentage of damaged and/or regenerating muscle in cross-sectional areas of all specimens from a mouse (n = 3 mice injected with each type of sample; one mouse died). Error bars indicate the estimated range of damage in all examined muscle specimens of each animal.
Bacterial gDNA was then fully digested with DNase I to confirm that the toxic component in the bacterial gDNA was sequence and/or length dependent and not a contaminant in the preparation (Fig. 6). The digested gDNA was from the same preparation as the sheared DNA. After the DNase digestion, the digested sample was spun through a filtration device with a molecular mass cutoff of 10 kDa to remove the 31-kDa DNase I and to recover fragments that were small. Full digestion with DNase I results in single-stranded fragments of approximately 4 bases (Matsuda and Ogoshi, 1966). HLV-injected DNase-digested bacterial gDNA (200 μg) caused no muscle damage, whereas sheared bacterial gDNA fragments did. Muscle damage resulted from even 20 μg of bacterial gDNA spiked into 200 μg of a pDNA preparation that itself caused no toxicity (Fig. 6).
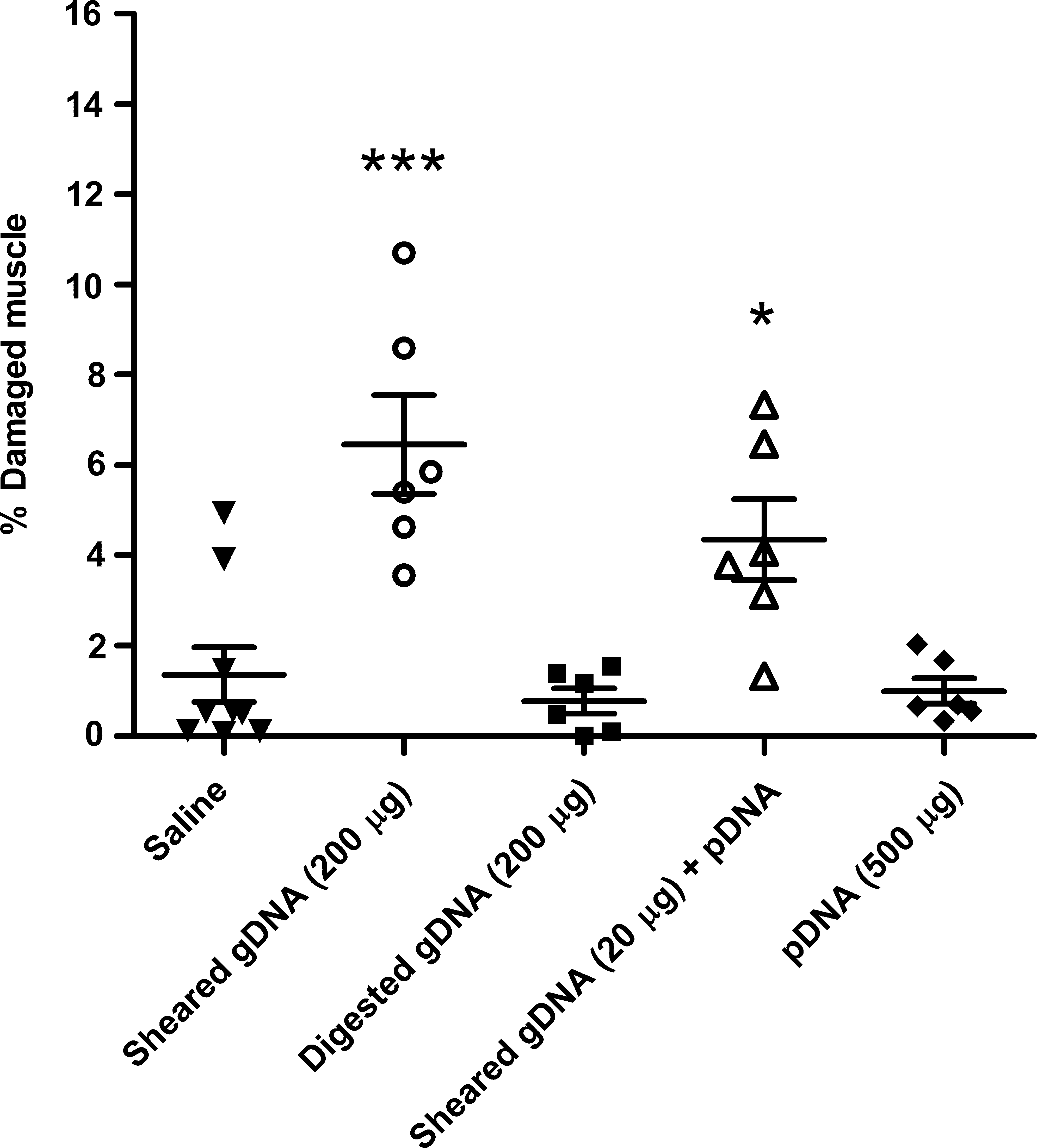
DNase digestion of bacterial gDNA eliminated its toxicity. ICR mice were treated by HLV injection of saline alone; 200 μg of sheared bacterial gDNA; 200 μg of DNase I-digested, sheared bacterial gDNA; 20 μg of sheared bacterial gDNA plus 200 μg of pMIR0603 that expressed mSEAP; or 500 μg of pMIR0603 alone (n = 6 mice injected with each sample and n = 9 mice injected with saline alone). Each data point represents all specimens evaluated from one leg; also shown are the group averages ± SEM (***p < 0.0001, *p < 0.05).
Plasmid DNA is bacterially produced, and the pUK21 vector is composed entirely of bacterially derived sequence. To determine whether linear fragments of a nontoxic preparation of supercoiled pUK21 would cause toxicity, we digested it with restriction enzymes to produce DNA fragments that varied in length from 3090 bp down to 8 bp, as described in Materials and Methods. ICR mice underwent one HLV injection of 500 μg of either the supercoiled or digested pUK21 and were then evaluated for muscle damage on day 3. None of the animals had any significant muscle damage (data not shown). Taken together, these observations confirm that the muscle damage was caused by large fragments of contaminating bacterial gDNA (more than 3 kb) in the pDNA preparations.
Discussion
In the process of performing preclinical gene therapy studies in rodents, we observed that some batches of pDNA caused significant muscle damage whereas other batches did not. The plasmids themselves did not cause muscle damage, nor did fragments of pDNA that were 3 kb or smaller, even though these contained an abundance of unmethylated CpG motifs in the plasmid backbone and variable amounts in the coding sequence. CpG-containing sequences appear to play a more important role when pDNA is formulated in liposomes and to be less critical for naked pDNA delivery (Yasuda et al., 2004, 2005; Zhang et al., 2005).
This pDNA batch-dependent toxicity could not be attributed to an immune response to expression of the encoded gene because some preparations of empty vector and of plasmids expressing the murine dystrophin gene in mice also caused muscle damage. Multiple batches of pDNA expressing nonmurine genes caused no damage. It is not surprising that we observed no toxicity related to expression of these foreign proteins, because maximal expression in muscle from plasmid vectors generally requires about 1 week and immune responses typically appear after 10 days. The batches of pDNA were evaluated for toxicity much earlier, on day 3 or 4 after injection.
Endotoxin levels and muscle damage did not correlate, although preparations with higher bacterial DNA content sometimes also had higher endotoxin levels. Even at a dose higher than in any of the pDNA preparations, endotoxin alone caused no muscle damage. Large bacterial DNA fragments containing negligible endotoxin caused muscle damage, but not after DNase digestion. Consistent with our results, McMahon and colleagues previously observed that endotoxin alone caused less muscle pathology than intramuscular injection of pDNA (McMahon et al., 1998).
The factor that consistently correlated with toxicity of pDNA preparations was the content of bacterial genomic DNA. Muscle-damaging preparations of pDNA contained bacterial gDNA and the severity of the toxicity correlated with the quantity of bacterial gDNA in the preparation. Initially the toxicity appeared to be related to the pDNA size, but even large plasmids could be purified as nontoxic batches, using a procedure that removes linear fragments of DNA (Schleef and Blaesen, 2009). Removal of contaminating gDNA from pDNA is achieved by a multistep process, beginning with a chromatographic capture step (for DNA binding) and followed by at least one polishing step to remove not only lipopolysaccharide endotoxins but also nonsupercoiled open circular and linear forms of plasmids and at the same time the linear bacterial gDNA fragments (Schleef and Schmidt, 2004). Separation of genomic DNA from larger plasmids is more challenging than from small plasmids, which could account for the greater number of large plasmid preparations causing toxicity. Nontoxic preparations of pDNA contained less than 0.5 EU of endotoxin per milligram of pDNA and less than 1.3% contamination with bacterial gDNA.
The toxic effect from large fragments of bacterial gDNA could be mediated by Toll-like receptors (TLRs), which are part of the innate immune system. TLR9 elicits an inflammatory response after detecting, in the endosome of certain immune cells, DNA with unmethylated CpG motifs. TLR9 is not expressed in muscle (see references in Bauer et al., 2008). TLR9 involvement would be consistent with our observation that the mammalian DNA fragments did not cause muscle damage. The immunostimulatory activity of vertebrate DNA is reduced by having fewer CpG motifs, methylation of these motifs when they are present, inclusion of motifs that inhibit immune activation, and a saturable DNA uptake mechanism in macrophages (Stacey et al., 2003).
Bacterial DNA could also have a direct effect on muscle cells. In contrast to sensing by TLR9, almost all nucleated cells, including muscle, respond to viral infection or the presence of dsDNA in the cytoplasm, such as from environmentally induced cell damage. The presence of the B-form of dsDNA in these nonimmune cells activates the cytokine cascade and results in an inflammatory response (Suzuki et al., 1999). If muscle damage were the result of sequence-independent immune stimulation by dsDNA, we would expect mammalian gDNA to also cause muscle damage, but it did not.
The batch-dependent toxicity of some preparations of pDNA can have a critical impact on pDNA-based gene therapy. It resulted in low and inconsistent expression of a therapeutic dystrophin gene in dystrophic mice that received multiple injections to increase the amount of gene product (data not shown), whereas mice injected with nontoxic batches of pDNA showed more consistent and additive amounts of gene expression from multiple injections (Zhang et al., 2010; and data not shown). Consistent with our results, batch-dependent differences in plasmid-encoded gene expression after intramuscular injection were previously noted by Manthorpe and colleagues (1993). To compare experimental results from different pDNA constructs or preparations, or from different experiments, or even to compare the published results from different research groups, we suggest that the amount of contaminating bacterial gDNA in pDNA preparations be reported along with the experimental results.
Unmethylated CpG dinucleotides act like an adjuvant for immunization (Krieg, 2002). Contaminating bacterial gDNA could serve this purpose during genetic immunization with naked pDNA. However, antibodies were efficiently generated in mice after a single delivery to the liver of a fairly small dose (approximately 30 μg) of naked pDNA from a vector that gives long-term liver expression (Bates et al., 2006). High titers of antibodies were also generated after two HLV injections, with only 10 μg of pDNA per injection. In these cases no significant amount of bacterial gDNA was likely to be present during the injection. The antibody response to the pDNA-encoded protein was most likely due simply to expression of the foreign protein. Therefore, although bacterial DNA can serve as an adjuvant, it is probably not essential for eliciting an immune response after genetic immunization.
Plasmid DNA manufacturers typically stipulate that the chromosomal DNA must be less than 5% of the total DNA preparation, and agarose gel electrophoresis is used for the quantitation. Large chromosomal fragments are generally detected as slowly migrating nucleic acid when the quantity is sufficient for visualization. These large chromosomal fragments can comigrate with large plasmids and are difficult to distinguish on a gel. Genomic DNA fragments can also be a variety of sizes, making them more difficult to detect. Therefore, agarose gel analysis is not satisfactory for quantitation of bacterial gDNA. Quantitative PCR is much more accurate for assessing the quality of the pDNA preparation. Furthermore, our results show that contamination of pDNA preparations with 5% bacterial gDNA is sufficient to cause extensive muscle toxicity. Batches of pDNA contaminated with bacterial gDNA may damage other tissues and organs and/or cause immunological effects after delivery by other methods. Therefore, we suggest that pDNA manufacturers redefine the allowable level of contaminating bacterial gDNA and implement quantitative PCR as a quality control assay.
In summary, plasmids can be purified in a manner such that they cause no muscle damage. Naked pDNA is not inherently toxic. Batches of pDNA can be tested for toxicity by HLV injection into rodent muscle, but PCR quantitation of bacterial gDNA is a simpler method of assessment that we strongly recommend. Preparations containing less than 1.3% bacterial gDNA in an HLV injection of pDNA at 0.5 mg/ml to the mouse limb rarely caused any significant muscle damage. Our results emphasize the importance of removing bacterial gDNA from pDNA preparations for gene therapy.
Footnotes
Acknowledgments
The authors thank Mark Noble, Tracie Milarch, Mavis Eldridge, Brett Hermanson, and Linda Goth for technical assistance with animals, Julia Hegge for helpful discussions, and Steven Coakley and Jeffrey Kim for histology (all from Roche Madison); Nicolas Fritsch (Transgene) and Katie Scarpace (Roche Madison) for endotoxin quantitation; and Jens Eickhoff (Colorado State University) for assistance with statistical analysis. Funding was provided by the Association Française contre les Myopathies.
Author Disclosure Statement
M. Schleef is CEO of the plasmid-manufacturing company PlasmidFactory.