
Abstract
The efficacy of gene therapy for the treatment of inherited immunodeficiency has been highlighted in recent clinical trials, although in some cases complicated by insertional mutagenesis and silencing of vector genomes through methylation. To minimize these effects, we have evaluated the use of regulatory elements that confer reliability of gene expression, but also lack potent indiscriminate enhancer activity. The Vav1 proximal promoter is particularly attractive in this regard and may be useful in situations where high-level or complex regulation of gene expression is not necessary. X-linked severe combined immunodeficiency (SCID-X1) is a good candidate for such an approach, particularly as there may be additional disease-related intrinsic risks of leukemogenesis, and where safety is therefore a paramount concern. We have tested whether lentiviral vectors expressing the common cytokine receptor gamma chain under the control of the proximal Vav1 gene promoter are effective for correction of signaling defects and the disease phenotype. Despite low-level gene expression, we observed near-complete restoration of cytokine-mediated STAT5 phosphorylation in a model cell line. Furthermore, at low vector copy number, highly effective T- and B-lymphocyte reconstitution was achieved in vivo in a murine model of SCID-X1, in both primary and secondary graft recipients. This vector configuration deserves further evaluation and consideration for future clinical trials.
Introduction
In previous studies, we have shown that lentiviral vectors (LVs) harboring the proximal promoter of the Vav1 murine proto-oncogene (Vav) (Ogilvy et al., 1998) mediate a weak, though homogeneous and stable, expression of transgenes not only in vitro, but also in vivo (Almarza et al., 2007). This latter characteristic relates to the CpG island nature of the promoter, which is resistant to methylation in engrafted transduced murine HSCs (Grassi et al., 2003; Brooks et al., 2004; Krishnan et al., 2006). We have also previously shown that an enhancerless bidirectional CpG island promoter (A2UCOE) mediates effective gene therapy in the context of a murine model of SCID-X1 (Zhang et al., 2007). Because of their similar properties, and due to the fact that bidirectional activity may be undesirable, we have now evaluated whether the Vav1 gene promoter can correct signaling and immunological reconstitution of SCID-X1 both in vitro and in vivo.
Materials and Methods
Vector construction
The sequence of the IL2Rγ was cloned into a self-inactivating (SIN) LV under the control of the Vav1 proximal promoter (Almarza et al., 2007). The sequence of the mutated Wpre (woodchuck hepatitis post-transcriptional regulatory element) (Schambach et al., 2006) was cloned downstream of the gene. As control vectors, we used identical SIN LV backbones in which the IL2RG expression was under the control of the SFFV or UCOE promoters (Zhang et al., 2007).
Production and titering of LV supernatants
LV supernatants were produced by transient cotransfection of 293T cells in the presence of polyethylenimine (Sigma-Aldrich, Dorset, UK). The VSV-G envelope plasmid (pMD.G2) and second-generation packaging plasmid (pCMVΔ8.74) were produced by Plasmid Factory (Bielefeld, Germany) and kindly provided by Luigi Naldini [San Raffaele Telethon Institute for Gene Therapy (HSR-TIGET), Vita Salute San Raffaele University, San Raffaele Scientific Institute, Milano, Italy]. The viral supernatants were concentrated by ultracentrifugation at 98,000 g for 2 hr at 4°C. The viral titer was determined by transducing SC1 cells with serial dilutions of the viruses. The expression was monitored by flow cytometry using an anti-human CD132 antibody (BD Pharmingen, Oxford, UK).
Cell lines
ED-7R cells [derived from an adult T-cell leukemia line deficient in IL2RG gene expression (Kumaki et al., 1999)] were cultured in Roswell Park Memorial Institute (RPMI) 1640 medium (Biochrom, Berlin, Germany). HT1080, 293T, and SC1 cells were maintained in Dulbecco's modified Eagle's medium. Both media (already containing Glutamax) were supplemented with 10% fetal calf serum (FCS), 100 U/ml penicillin/streptomycin.
Quantitative PCR
To determine the vector copy number in transduced cells, the genomic DNA was extracted using the DNAeasy extraction kit (Qiagen, West Sussex, UK). The quantitative PCR was performed in an ABI 7000 Sequence Detection System (ABI, Applied Biosystems, Warrington, UK). To amplify the vector, we used sequences annealing the mutant Wpre region. As an endogenous two-copy gene control, we used the titin gene for murine cells and β-actin gene for human cells. The primer sequences for mutant Wpre were as follows: forward, 5′-TGGATTCTGCGCGGGA-3′; and reverse, 5′-GAAGGAAGGTCCGCTGGATT-3′. The mutant Wpre probe sequence was 5′-FAM_CTTCTGCTACGTCCCTTCGGCCCT_TAMRA-3′. The primer sequences for the murine Titin were as follows: forward, 5′-AAAACGAGCAGTGACCTGAGG-3′; and reverse, 5′-TTCAGTCATGCTGCTAGCGC-3′. The Titin probe sequence was 5′-FAM_TGCACGGAATCTCGTCTCAGTC_TAMRA-3′. The primer sequences for the human β-actin gene were as follows: forward, 5′-TCACCCACACTGTGCCCATCTACGA-3′; and reverse, 5′-CAGCGGAACCGCTCATTGCCAATGG-3′. The β-actin probe sequence was 5′-FAM_ATGCCCTCCCCCATGCCATCCTGCGT_TAMRA-3′. Primer/probe sequences and standards for the mutant WPRE and titin gene were a kind gift from Dr. Anne Galy (Genethon, Evry, France).
Animals
γc–/– mice (very low percentage of T cells, no NK cells, and presence of nonfunctional B cells) were generated by crossing γc–/–Rag2–/– mice (no T, no B, and no NK cells) with Balb/c mice (γc+/+Rag2+/+ ; Harlan UK, Bicester, UK). For the UCOEIL2RG-LV transplant experiments, we used the same mice, but generated in the C57BL/6 strain. As previously described, similar phenotypes were observed in these knockouts in Balb/c and B6 strains (Custer et al., 1985). All experimental procedures were approved by the Institutional Research Ethics Committee (Institute of Child Health, University College London, UK) and performed according to UK Home Office Animal Welfare Legislation.
Ex vivo gene therapy of the SCID-X1 murine model
Bone marrow (BM) cells were harvested from the femurs and tibias of γc–/– or Balb/c mice, and the lineage negative (lin−) progenitors were isolated using a StemSep mouse progenitor enrichment cocktail and negative selection columns following the manufacturer's instructions (StemCell Technologies, London, UK). Lin− cells were transduced for a total period of 20 hr [multiplicity of infection (MOI): 10 transducing units (TU)/cell] in StemSpan SFEM serum-free medium (StemCell Technologies) supplemented with 1% penicillin/streptomycin and cytokines (100 ng/ml murine stem cell factor, 100 ng/ml mFlt3L, 100 ng/ml hIL-11, 20 ng/ml mIL-3). Transduced cells were then harvested, and 2–3 × 105 transduced lin− cells were injected intravenously into the tail veins of sublethally irradiated (6 Gy) γc–/–Rag2–/– mice. For the secondary transplants, 107 total BM cells from primary recipients were transplanted per mouse. Two secondary mice were transplanted when possible.
STAT5 phosphorylation assay
This assay was performed as previously described (Zhang et al., 2007). The control ED-7R-γc cells have been described previously (Kumaki et al., 1999) and were maintained in RPMI 1640 medium supplemented with 10% FCS, 100 U/ml penicillin/streptomycin, and 2 mmol/L glutamine.
Immunoglobulin (Ig) ELISA
Serum isolated from the peripheral blood of transplanted mice was diluted appropriately and added to NUNC-Immuno ELISA plates (NUNC, Fischer Scientific UK, Loughborough, UK) precoated with capture antibody—IgG (AbD Serotec, Oxford, UK), IgG1 (BD Pharmingen), or IgG2a (BD Pharmingen). Plates were incubated for 1–2 hr at room temperature, washed three times with phosphate-buffered saline/0.05% Tween 20, and incubated with a horseradish peroxidase–conjugated detection antibody at room temperature for 1 hr. For the IgG ELISAs, a biotinylated detection antibody was used followed by 30-min incubation with a strepavidin–horseradish peroxidase tertiary antibody. Plates were washed a further three times in phosphate-buffered saline/0.05% Tween 20, after which substrates A and B from an ELISA Duo-set substrate reagent pack (R&D Systems, Abingdon, UK) were mixed in equal quantities and 100 μl of the mixed solutions was added to each well. Plates were incubated at room temperature in the dark for ∼10 min to allow color change, at which time the reaction was stopped by adding 50 μl of stop solution/well (2 N H2SO4). The absorbance of each plate was then read at 405 nm using a FLUOstar Optima plate reader (BMG Labtech).
Analysis of immune reconstitution
At 5 months after transplantation, animals were culled and peripheral blood, BM, spleens, thymus, and lymph nodes were isolated and analyzed for immunoreconstitution as previously described (Zhang et al., 2007).
Statistical analysis
The statistical analysis was done using the GraphPad Prism4 software. For all the analyses, we performed a nonparametric Mann-Whitney test. The significances are expressed as p < 0.01(**) or p < 0.05(*).
Results
Restoration of IL-2-mediated signaling in vitro
As a first approach to evaluate the efficacy of VavIL2RG-LV, we tested its capacity to restore γc-mediated signaling in vitro in human ED-7R cells, deficient in γc. ED-7R cells were transduced with VavIL2RG-LVs, and also with SFFVIL2RG- and UCOEIL2RG-LVs, as control vectors previously shown to correct the deficiency (Zhang et al., 2007; Thornhill et al., 2008). A schematic of the LVs used in this study and the Vav HS1 promoter with most significant transcription factor binding sites are shown in Fig. 1A and B, respectively. Seven days after transduction, cells were stained with an anti-γc antibody and analyzed by flow cytometry. As shown in Fig. 1C, most ED-7R cells exhibited strong expression of γc after transduction with SFFVIL2RG-LVs. In contrast, ED-7R cells transduced with similar MOIs of UCOEIL2RG-LVs and of VavIL2RG-LVs expressed much lower levels. Because phosphorylation of STAT5 (signal transducer and activator of transcription 5) by JAK3 (Janus kinase 3) is a key step in the signaling pathway activated by IL-2, levels of phosphorylated STAT5 (pSTAT5) were determined in unstimulated and IL-2-stimulated ED-7R cells that had been transduced with the different LVs. As expected, no pSTAT5 was detected in unstimulated ED-7R transduced cells. However, after stimulation with IL-2, similar levels of pSTAT5 were observed in ED-7R cells transduced with all tested vectors, suggesting that the expression of other members of this receptor complex (either the alpha or the beta chains) may limit the extent of signaling once threshold levels of expression of the γc are achieved. We deduce from these experiments that low-level expression of γc is sufficient to restore the biochemical pathway mediated by the interaction of IL-2 with its receptor.
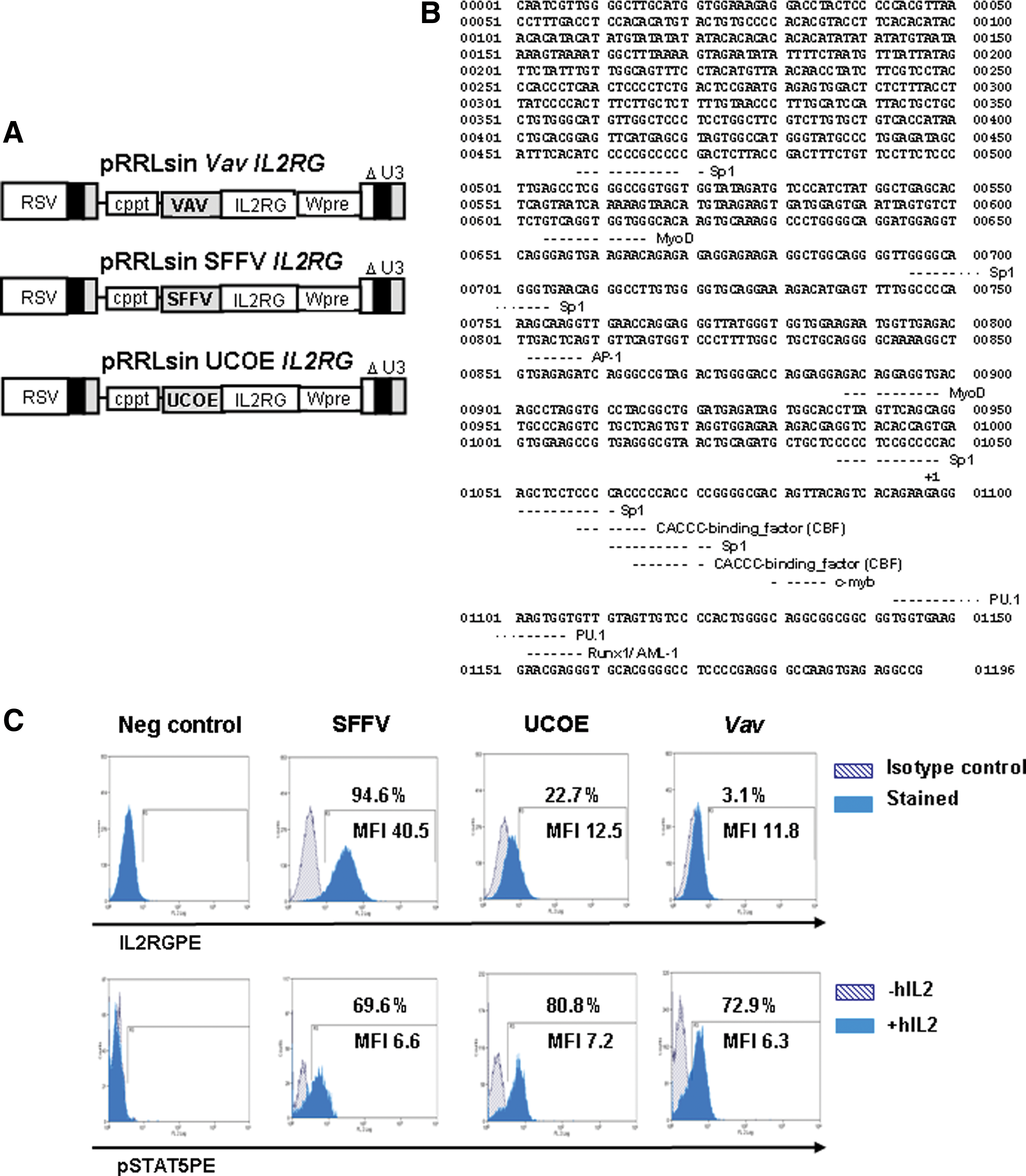
Restoration of IL-2-mediated signaling in γc-deficient ED-7R cells. (
Ex vivo gene therapy of the SCID-X1 murine model with VavIL2RG-LV
Lin− BM cells from γc-deficient mice were transduced with VavIL2RG-LV (MOI: 10 TU/cell). Transduced cells were cultured in methylcellulose medium for 7 days. Individual colonies were picked, and DNA was extracted to analyze the mean transduction efficiency by quantitative PCR. The percentage of colonies positive for the integrated provirus was 74%, the mean vector copy number per transduced cell being 1.2 copies/cell. To investigate the therapeutic effect of VavIL2RG-LVs, transduced γc-/- cells were transplanted into sublethally irradiated immunodeficient γc–/–rag2–/– recipients (Otsu et al., 2000). These double-knockout mice were used as recipients because they have a stable, alymphoid phenotype, in contrast to mice deficient only in γc, which contain a residual number of T lymphocytes and are susceptible to inflammatory bowel disease (DiSanto et al., 1995).
Mean vector copy numbers determined in the spleen of γc–/–rag2–/– recipients transplanted with γc–/– corrected cells were 0.6 ± 0.4 at 5 months post engraftment. Representative histograms of the immunophenotype of cells obtained from peripheral blood [1 month post bone marrow transplantation (BMT)] and from the spleen, thymus, and lymph nodes of γc–/–rag2–/– recipients (5 months post BMT) transplanted with transduced γc–/– cells are shown in Fig. 2. Data obtained from a negative control γc–/– and two positive controls [untransplanted wild-type (WT) mice and γc–/–Rag2–/– mice transplanted with WT cells] are also shown. Mean values obtained from analysis of all mice are shown in Fig. 3. The analysis of peripheral blood from γc–/–rag2–/– mice transplanted with VavIL2RG-transduced γc–/– cell mice indicates good restoration of CD3+ and B220+ cell populations, similar to that achieved following transplantation of WT cells (see Figs. 2 and 3). Spleen and lymph nodes from γc–/–rag2–/– recipients transplanted with γc–/– VavIL2RG-transduced cells contained normal numbers of mature CD4 and CD8 T cells, and mature B220+ lymphocytes were also restored in spleens. In the thymus, similar numbers of CD4+ and CD8+ cells were observed compared with mice transplanted with WT cells.
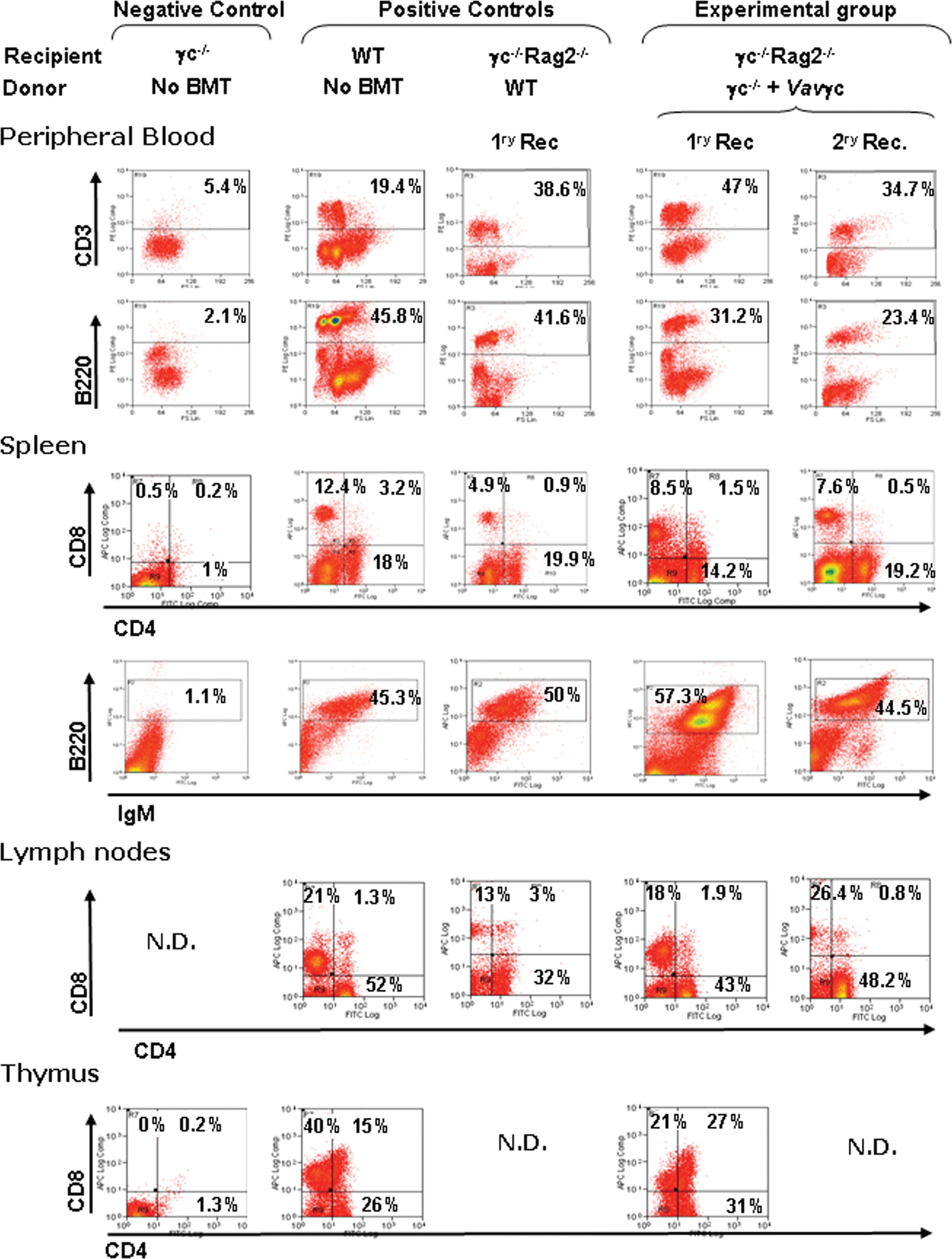
Ex vivo gene therapy of mice with the VavIL2RG-LV. In the experimental group, Lin− cells from γc–/– mice were transduced with the VavIL2RG-LV and transplanted into sublethally irradiated γc–/–Rag2–/– mice [primary recipients (1ry Rec.)]. Secondary transplants were carried out with BM cells from Vavγc primary recipients (2ry Rec.). As positive controls, we used cells from untransplanted WT mice as well as from sublethally irradiated γc–/–Rag2–/– mice transplanted with total BM cells from WT mice. As negative control, cells from untransplanted γc–/– mice were used. Dot plots show representative flow cytometry analyses of T (CD4+, CD8+, and double-positive) and B cells (B+IgM+) from the peripheral blood and the lymphoid organs of gene therapy-treated SCID mice, and their respective controls. N.D., not done because no material could be collected. Color images available online at
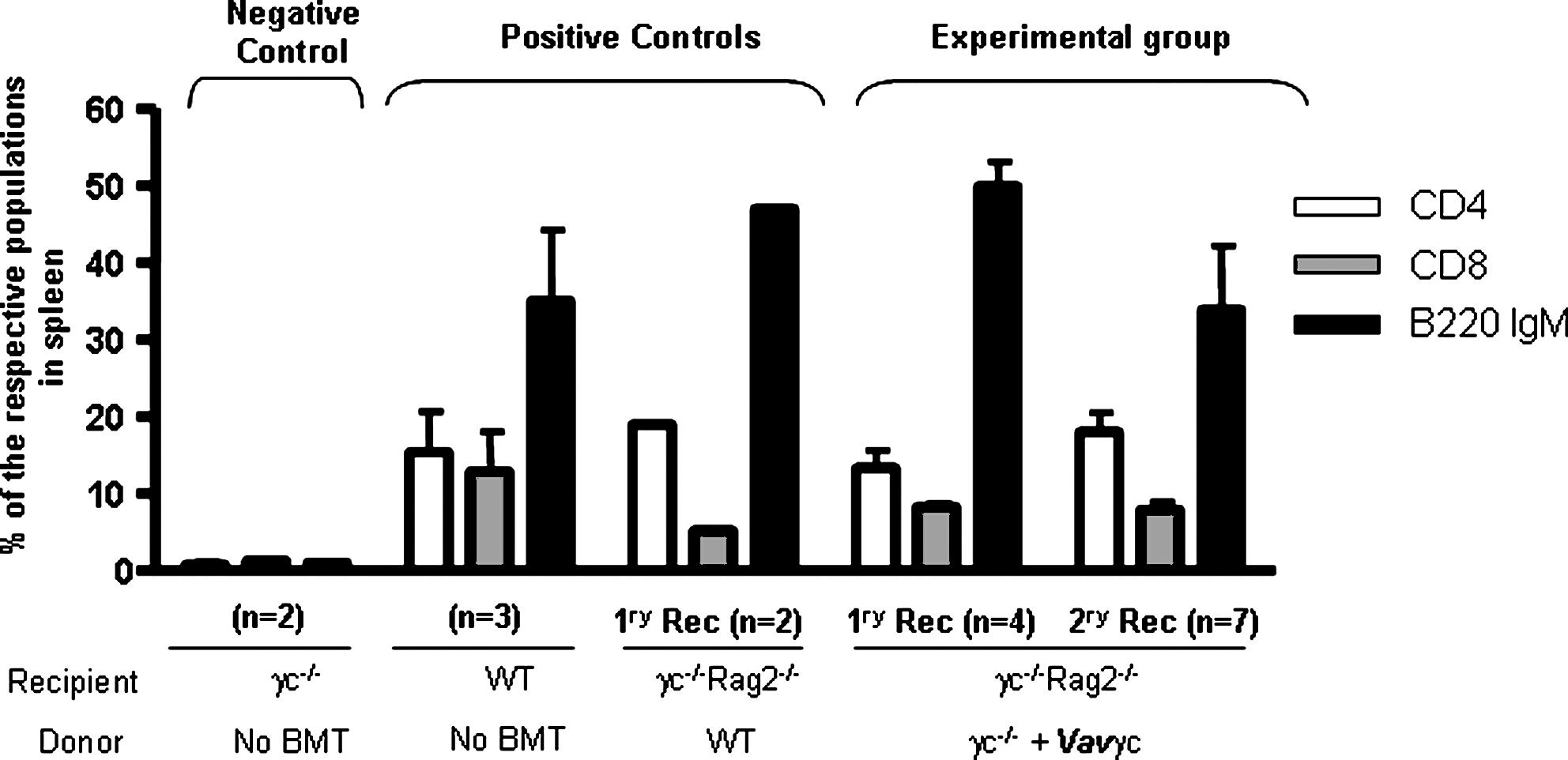
Lymphoid reconstitution in the spleen of mice subjected to gene therapy. Mean values corresponding to the different lymphoid populations present in the spleen of mice treated by gene therapy and their respective controls are shown. The represented values, shown as means ± SD, were obtained at 5 months after transplant.
Finally, to demonstrate effective transduction of primitive HSCs, BM cells from primary recipients were retransplanted into secondary γc–/–rag2–/– recipients. As observed in primary recipients, good T- and B-cell reconstitution occurred (see representative histograms in Fig. 2 and mean values in Fig. 3). Interestingly, the effectiveness of reconstitution was similar to that achieved with the bidirectional alternative CpG island promoter (A2UCOE), supporting the fact that low-level gene expression from both constructs is sufficient for rescue of the immunological phenotype (Supplementary Fig. 1A; supplementary data are available online at
Functional reconstitution of primary and secondary recipients
To test whether T-lymphocyte populations generated in mice transplanted with transduced samples were functional, we performed in vitro proliferation assays with splenocytes obtained 5 months after transplantation. Data in Fig. 4 show relative values of lymphocyte proliferation after stimulation with concanavalin A (ConA) alone or combined with IL-2, compared with basal levels obtained in γc–/– mice. As positive controls, lymphocytes from γc–/–rag2–/– mice transplanted with WT cells and lymphocytes from untransplanted WT animals were used. Lymphocytes from recipients transplanted with BM cells transduced with either Vav- (Fig. 4A) or A2UCOE-regulated LVs (Supplementary Fig. 1B) showed proliferation indexes significantly increased compared with values obtained from the negative control. These values were also not significantly different from those obtained from untransplanted WT animals (p > 0.05). Finally, we investigated humoral immunity in mice transplanted with γc–/– cells genetically treated with VavIL2RG-LVs. In contrast to the very low levels of immunoglobulins IgG1, IgG2a, and total IgG found in untreated γc–/– mice, in mice receiving gene-corrected cells, levels were restored in both primary and secondary recipients (Fig. 4B). Altogether these results confirm that low levels of γc expression are sufficient to mediate functional correction of γc-deficient primary lymphocytes in vivo.

Functional reconstitution of transplanted mice subjected to gene therapy with VavIL2RG-LV. (
Discussion
The first successful attempts to treat monogenic hematopoietic disorders used technology based on gammaretroviral vectors that employed potent viral regulatory enhancer elements to mediate transgene expression. These elements are recognized as mutagenic both in their natural WT viral configuration and, more recently, when used in the context of gene transfer vectors. To improve the safety of gene therapy, new vector types and configurations have been developed. Among them, SIN LVs have been proposed as one of the most promising. Apart from their capacity to transduce minimally stimulated HSCs, LVs do not integrate preferentially in the region of transcription factor binding sites and have a more limited potential to generate leukemias in mice when compared with LTR-intact gammaretroviral vectors (Montini et al., 2006). Similarly, a recently developed in vitro transformation assay has demonstrated that internal promoter sequences in the context of SIN LVs or gammaretroviral vectors are much less mutagenic, particularly if weaker elements are used to direct transgene expression (Modlich et al., 2009). We have previously shown that the proximal promoter of the Vav1 proto-oncogene (the Vav promoter) is able to direct homogeneous and stable gene expression of either marker or therapeutic transgenes both in vitro and in vivo (Almarza et al., 2007). Furthermore, LVs harboring this element induce no more than background levels of transformation in hematopoietic progenitor cells in vitro (Modlich et al., 2009). Recent unpublished studies from our laboratory have demonstrated that the Vav promoter also maintains these characteristics in the context of SIN gammaretroviral vectors. Based on these observations, we concluded that the use of the Vav promoter may improve the safety of gene therapy strategies in which moderate or low levels of the therapeutic transgene product are required (Zhang et al., 2007).
We have therefore generated a SIN LV in which the Vav promoter regulates the expression of the IL2RG transgene. We tested the ability of VavIL2RG-LVs to reconstitute signaling in a model cell line, and also in a murine model of SCID-X1 in both primary and secondary transplant recipients. Overall, this vector was able to mediate complete restoration of function despite relatively low potency in terms of gene expression. The fact that secondary recipients achieved normal levels of reconstitution supports the reliability of this promoter and its resistance to silencing in primitive cell populations such as HSCs, although more extensive preclinical testing will be required before clinical application with this element can be considered. The results obtained here are consistent with our previous studies using an alternative CpG island-based promoter (A2UCOE), which shares a lack of potent enhancer activity (Zhang et al., 2007). However, in its basic configuration, the A2UCOE element has bidirectional activity and has residual splice donor sites that could promote dysregulation of genes in the surrounding environment. Studies are ongoing to correct these potential deficiencies, but the Vav promoter represents a viable and comparable alternative regulatory sequence.
Although CpG island-based regulatory elements such as the Vav1 gene proximal promoter mediate relatively weak gene expression, for some applications this may be advantageous and could obviate transgene-mediated toxicity (Baum et al., 2004, 2006; Kustikova et al., 2005; Montini et al., 2006). The lack of potent enhancer sequences (Ogilvy et al., 1998) also means that inadvertent transactivation of other genes is less likely. Patients with SCID-X1 treated by gene therapy using LTR-intact gammaretroviral vectors have been shown to be susceptible to leukemogenesis primarily as a result of insertional mutagenesis, although other factors, including γc overexpression, replicative stress, and disease background, have been suggested to operate as cofactors in this process (Woods et al., 2003; Nienhuis et al., 2006; Shou et al., 2006; Ginn et al., 2010). At present, these remain controversial and undefined, but deserve consideration for the design of future protocols. Replication of natural gene regulation would seem to be an obvious goal, but is complicated by the fact that even natural regulatory sequences may behave aberrantly when integrated in different regions of the chromosome. This was highlighted recently in a study in which a genomic IL2RG fragment containing natural promoter and enhancer sequences was shown to transactivate LMO2 (similar to an LTR-based vector) when placed close by, whereas a housekeeping EF1α promoter did not have this effect (Zhou et al., 2010). Development of endonuclease-enhanced gene-targeting strategies are therefore of considerable interest (Lombardo et al., 2007).
Overexpression of γc has been postulated as a cofactor in leukemogenesis on the basis of lentiviral studies in SCID-X1 murine models and in murine tumor models containing coincident retroviral insertions at the LMO2 genomic locus and near the IL2RG locus (Woods et al., 2006; Dave et al., 2009). In EBV-immortalized cell lines derived from patients with SCID-X1 and in which γc expression was suppressed by short interfering RNA, it was found that the amount of γc influenced spontaneous proliferation and JAK3 phosphorylation, but also growth hormone-induced cell proliferation (Amorosi et al., 2009). It was therefore suggested that expression levels are important for control of cell growth. Despite these findings, there has been no evidence for overexpression or constitutive activation of STAT5 phosphorylation in patients in either gene-transduced T cells or leukemic clones (Howe et al., 2008).
Taken together, data obtained in this study suggest that SIN LVs harboring CpG island-based promoters such as that obtained from the Vav1 proto-oncogene provide effective and reliable gene expression for long-term correction of disease phenotypes such as SCID-X1, particularly where high-level or complex regulation is not required. Furthermore, they offer distinct advantages with respect to safety and deserve further evaluation for clinical use.
Footnotes
Acknowledgments
Research at the Institute of Child Health and Great Ormond Street Hospital for Children NHS Trust benefits from R&D funding received from the NHS Executive. A.J.T. is a Senior Wellcome Trust Clinical fellow. E.A. was supported partially by the Spanish network CIBERER plus a grant given by the José Castillejo program of the Spanish government. G.S. was supported by the CGD Research Trust, and M.P.B. by the Wellcome Trust.
The authors also wish to thank the technical assistance of Aurora de La Cal and Sergio Losada (CIEMAT/CIBERER). This work was supported by grants from the European Program FP7 (PERSIST; Ref 222878), Centro de Investigación en Red de Enfermedades Raras (CIBERER), International Cooperation on Stem Cell Research Plan E (Ref PLE 2009/0100); Comisión Interministerial de Ciencia y Tecnología (SAF 2009-2103). The authors also thank the Fundación Marcelino Botín for promoting translational research at the División de Hematopoyesis y Terapia Génica at the CIEMAT and the Centro de Transfusiones de la Comunidad de Madrid for providing cord blood samples. CIBERER is an initiative of the Instituto de Salud Carlos III.
Author Disclosure Statement
All authors have nothing to disclose.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.