
Abstract
Hydrodynamic gene delivery to the liver is a promising approach for liver gene therapy in the clinic, but levels of gene expression in larger species have been much less than in rodents. The development of surgical techniques for pressurizing individual liver segments and the establishment of whether hepatic vascular anatomy in fact permits pressurization of individual segments are critical issues that need to be addressed. We have evaluated these issues using hydrodynamic delivery to individual segments of the pig liver, via branches of both portal and hepatic veins. Our objective was to develop surgical techniques that achieve elevated vascular pressures within individual liver segments with small volumes, but without interruption of portal blood flow or reduction in venous return to the heart. We report that, without specific surgical interventions to obstruct outflow of DNA solution from the targeted liver segment, little or no increase in intrahepatic vascular pressure occurs. We demonstrate, for the first time, that selective pressurization of individual liver segments is possible without compromising portal venous flow or venous return to the heart. Thus, hydrodynamic gene delivery to individual liver segments is technically achievable in a clinical setting, but will require open abdominal surgery rather than minimally invasive techniques.
Introduction
As we have previously discussed (Sawyer et al., 2009), the driving force for swelling of the liver must come from raised intrahepatic vascular pressure (especially in the more easily distensible sinusoids and veins), and raised vascular pressure can occur only if there is resistance to flow. We have proposed that, during hydrodynamic gene delivery via the tail vein, the DNA solution passes retrograde through the liver into the portal vein and that the vascular resistance of the capillary bed of the gut, spleen, and pancreas provides the critical obstruction of outflow from the portal vein (Sawyer et al., 2009). The absence of any surgical intervention to block DNA outflow from the liver during hydrodynamic gene delivery via the tail vein has, misleadingly in our view, probably been responsible for the widespread idea that minimally invasive techniques will be possible in the clinic. These techniques involve insertion of balloon catheters into a femoral vein or the right internal jugular vein via a small skin incision, positioning the catheter in a branch of a hepatic vein, inflating the balloon to prevent backflow, and delivery of the DNA solution. No attempt is made to block outflow of the DNA solution from the targeted segment of liver.
Minimally invasive techniques have been attempted in pigs (Herrero et al., 2005; Yoshino et al., 2006; Aliño et al., 2007; Suda et al., 2008) and humans (Khorsandi et al., 2008). However, reporter gene expression in the pig studies was several hundred- to 10,000-fold lower than in the mouse, and there was no substantive evidence for gene expression in the clinical study. Perfusion pressures were not measured in the pig studies, and only syringe pressures (which are of little value, as most pressure loss occurs in the delivery lines and connections) were reported in the clinical study. Eastman et al. (2002) were conscious of the problem that minimally invasive techniques essentially involve injecting DNA solution into an open system. They demonstrated by fluoroscopy in a rabbit model that the DNA solution passed through the target segment of liver into the portal vein and then freely back through the other segments of the liver into the inferior vena cava (IVC). They attempted to block all hepatic venous outflow with a balloon in the IVC, but technical difficulties with available balloon catheters in the relatively small animals used (∼3-kg rabbits) meant that only one technical success (with poor gene delivery) was reported.
We have explored these problems using the model of regional hydrodynamic gene delivery to the pig liver, using branches of both portal and hepatic veins for DNA delivery. Our primary objective was the measurement of vascular pressures within the targeted segment of liver and the cardiovascular responses to the surgical procedures. Although the pGL3 plasmid was used in some hydrodynamic deliveries reported in this study, the luciferase levels obtained were low, as expected from previous studies in the pig (Fabre et al., 2008; Kamimura et al., 2009). Our objective was to develop surgical techniques that achieve elevated vascular pressures within individual liver segments, with small volumes (<1% of body weight) but without interruption of portal blood flow and without reduction in venous return to the heart (thereby avoiding gut congestion and ischemia, and cardiovascular problems) (Sawyer et al., 2007, 2008; Fabre et al., 2008).
Materials and Methods
Animals
Large White × Landrace cross female pigs (PSQ, Clifton, UK) were used. All procedures were approved by the King's College London Ethics Committee and by the U.K. Home Office.
DNA plasmid
The pGL3 plasmid containing the firefly luciferase gene (Promega, Madison, WI) was used. DNA came from a single, sterile, endototoxin-free batch of 600 mg (PlasmidFactory, Bielefeld, Germany). It was supplied frozen at 1 mg/ml in pure water and stored at −35°C. DNA was thawed and made up to 50 μg/ml in 0.15 M NaCl immediately before the perfusion, and any unused DNA was discarded.
Hydrodynamic gene delivery
Pigs were given azaperone (2 mg/kg), ketamine (10 mg/kg), and atropine (60 μg) (all intramuscularly). Isoflurane was used for general anesthesia. A standard three-lead electrocardiogram (left front leg, right front leg, left hind leg) was used. For postoperative analgesia, buprenorphine (5 μg/kg) was given intramuscularly every 8 hr.
The MK5 angiographic injector syringe pump with a 200-ml syringe (Medrad, Ely, UK) was used for the perfusions. When a single pump was used, it was linked to the perfusion catheter via a high-pressure contrast injection line (1,000 psi rated) (Medrad). For volumes from 200 to 400 ml, two pumps were used in parallel, linked by a high-pressure (400 psi-rated) Y connector (Kimal, Uxbridge, UK). The pumps have a maximum speed of 50 ml/sec, thus giving a maximum capacity of 400 ml and a maximum flow rate of 100 ml/sec (6 liters/min). The rise time (time to reach the set flow rate) was set at 0.1 sec.
Perfusion catheters
Retrograde coronary sinus perfusion catheters (Medtronic, Minneapolis, MN) were used for most procedures. The 15 French gauge was used for delivery into the portal vein or into the IVC, and the 13 French gauge for delivery into a branch of the portal vein. For delivery into a hepatic vein via the right internal jugular vein, a long catheter was required, and we used the 9 French Cardiovasion catheter (Edwards Life Sciences, Irvine, CA).
Occlusion catheters
For selective occlusion of either the left/middle or right hepatic veins by a balloon in the IVC, the equalizer balloon catheter (product number M001171120, Boston Scientific, Natick, MA) (balloon diameter, 40 mm) was used. For occlusion of both hepatic veins by a balloon in the IVC, the Atlas PTA balloon dilatation catheter (balloon diameter, 22 mm; balloon length, 4 cm) (catalog number AT-75224, Bard Peripheral Vascular Inc., Tempe, AZ) was used. This provided the required length and cylindrical shape of balloon for optimal occlusion of both hepatic veins.
Pressure measurements
Cannulas (20 French gauge) (Abbott Laboratories, Queensborough, UK) were placed in the aorta, IVC, and portal vein as required. These (and the pressure lines incorporated in the perfusion catheters) were connected via 200 cm of saline-filled high-pressure manometer tubing 1 mm in diameter (to avoid damping effects) (Vygon, Cirencester, UK) to an electrical pressure transducer system (LogiCal single line transducer monitoring kit, Medex Medical, Strathclyde, UK). The transducers were placed at the level of the heart in the supine pig and opened to atmospheric pressure to obtain the zero baseline. This ensured that pressure measurements were obtained with reference to a constant baseline, namely, the level of the heart. The pressure signal was converted to a digital signal, as previously described (Sawyer et al., 2007). The digital signal was transferred to a laptop computer where the pressure recordings and electrocardiogram were acquired and displayed graphically using AcqKnowledge waveform analysis software, version 3.9 (BioPac Systems, Linton Instrumentation, Norfolk, UK). The software allowed all recordings to be displayed in a simultaneous time domain and for event markers to be placed in real time to highlight critical interventions. The heart rate (pulse) was derived from the arterial pressure trace.
In the earliest of the studies reported here (see pigs 8–10, Table 2), pressures were measured using a PVB pressure transducer (Peter von Berg Medizintechnik, Eglharting, Germany), and the pressure signals were digitized using the processing apparatus on the Deltastream pump (Medos Medizintechnik, Aachen, Germany). The digitized signals were exported to Microsoft (Redmond, WA) Excel for data analysis.
Harvesting of liver for assay of luciferase reporter gene expression
This was performed 1 day after gene delivery. The hepatic artery and portal vein were clamped with the animal under isoflurane general anesthesia, and 60 mg/kg pentobarbitone sodium was injected intravenously to kill the pig. A cut was then made in the IVC, and a 14 French gauge cannula was inserted into the portal vein for perfusion of 1.5 liters of 0.15 M NaCl. This was to remove blood, as hemoglobin can interfere with luciferase assays (Smith and Trempe, 2000). The liver was then removed, and ∼10 biopsy specimens (each ∼8 × 8 × 15 mm) from different lobes were taken and frozen.
The liver pieces were thawed, weighed, and minced with scissors in a Petri dish, and 2 ml of lysis buffer (Promega) was added per gram of tissue. The tissue was then homogenized with a ground glass homogenizer. The aqueous tissue extract was prepared and assayed for luciferase activity and protein concentration as previously described (Sawyer et al., 2007). Luciferase activity was expressed as relative light units per milligram of protein per 10 sec, using a luciferase assay kit from Promega, and a 10-sec measurement of luminescence (Anthos Lucy 1, Labtech International, Lewes, UK).
Fluoroscopy
Omnipaque contrast medium (Amersham, Oslo, Norway) in 0.15 M NaCl was used, and the fluoroscopic image was obtained using a Stenoscop 2 fluoroscope (General Electric, Les Moulineux, France).
Results
The critical importance of outflow obstruction for pressurization of the liver
In a previous study in the pig model, we reported that a volume corresponding to 2% of body weight, when delivered via the isolated IVC segment retrograde to the whole liver at 6 liters/min (100 ml/sec), generated intrahepatic vascular pressures of 101–126 mm Hg. We suggested that this was a consequence of the natural obstruction to outflow of the DNA solution from the portal vein, mediated by the capillary beds of the intestines, spleen, and pancreas (Fabre et al., 2008). In the present study, we delivered the 2% volume to the whole liver at the same flow rate, but in the opposite direction (i.e., via the portal vein). In this direction of flow, on exiting the liver into the hepatic veins, there is no natural obstruction to flow.
Two of the four pigs in this group showed pressure profiles as in Fig. 1. In the pig illustrated, the portal venous pressure (top panel) dropped dramatically immediately once the infusion was started and remained well below the initial pressure for the duration of the 3.7–3.8-sec infusion (370 ml at 100 ml/sec, plus rise time of the pump of 0.1 sec). This negative pressure is a direct consequence of Bernoulli's principle and indicates that the flow of fluid through the liver met essentially no resistance. Thus, in these two pigs, rather than pressurizing the liver, the rapid infusion actually created a negative pressure.
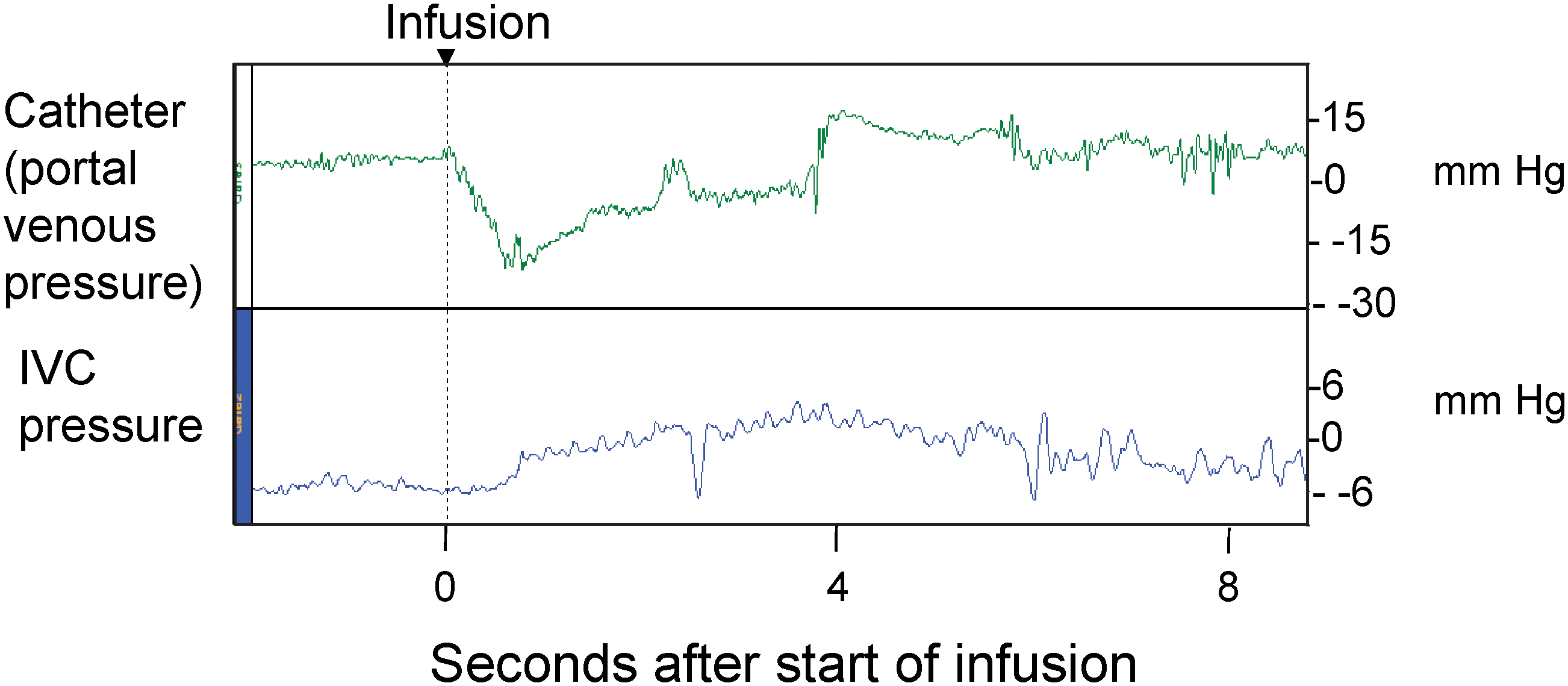
Vascular pressures during hydrodynamic delivery into the portal vein. An 18.5-kg pig was given a 370-ml infusion of 0.15 M NaCl (2% of body weight) into the portal vein at 100 ml/sec (6 liters/min). (
In the third pig (weighing 19 kg) the portal pressure recording fluctuated wildly for the first 2.05 sec (data not shown). The portal vein wall was observed to flap during the infusion, presumably because of negative pressures generated. The pressure recordings almost certainly represent artifacts caused by the wall of the portal vein hitting the tip of the pressure line. This needs to be avoided in the clinical situation because perfusion pressure will almost certainly control the perfusion pump. In the last pig (weighing 18.6 kg), the portal pressures were much greater than the IVC pressures, suggesting vascular resistance within the liver, possibly due to vasospasm.
Hydrodynamic delivery via a hepatic vein branch, without outflow obstruction (“minimally invasive” technique)
A critical evaluation of this technique is important, as it has already been used in a clinical study (Khorsandi et al., 2008). The perfusion catheter (9 French, Cardiovasion) was introduced into the right internal jugular vein, through the right atrium into the IVC, and positioned in a branch of a hepatic vein. On balloon inflation, the region of liver drained by the hepatic vein branch became distinctly cyanotic (blue), with a sharp demarcation line. A standard volume of 200 ml was delivered over 4 sec (3 liters/min) to 28–38-kg pigs. Care was taken that the region of liver targeted was reasonably large (17–26% of the whole liver). These conditions correspond to ∼1 ml/g of targeted liver and a flow rate of ∼15–20 ml/min/g of targeted liver (Table 1), similar to the optimal hydrodynamic conditions in the rat (Sawyer et al., 2008). By comparison, in the clinical study, 50–300 ml was delivered to adult patients over a period reported as less than 1 min (Khorsandi et al., 2008).
Measured at day 1, at time of harvest.
Right lateral lobe.
Saline without DNA was delivered.
Medial half of right median lobe.
Equipment malfunction.
Left lateral lobe.
RLU, relative light units.
The pressures generated were very low (Table 1 and Fig. 2). In pig 46 (Fig. 2a, top panel), the pressure in the hepatic vein branch fell throughout the infusion, suggesting little resistance to flow; portal venous pressure (Fig. 2a, bottom panel) rose by only 6.2 mm Hg. In pig 47, the pressure in the hepatic vein branch (Fig. 2b, top panel) initially fell during the infusion and then rose to 23.3 mm Hg, a rise of only 8.5 mm Hg; the portal venous pressure (Fig. 2b, bottom panel) rose by only 5.2 mm Hg. In pig 43, the pressure in the hepatic vein branch rose to a peak of 26.6 mm Hg, a rise of 16.5 mm Hg (Fig. 2c, top panel); the portal pressure (Fig. 2c, bottom panel) rose by 5.1 mm Hg. Thus, the minimally invasive technique causes little or no pressurization of the targeted segment of liver and is therefore unlikely to give rise to a hydrodynamic effect.
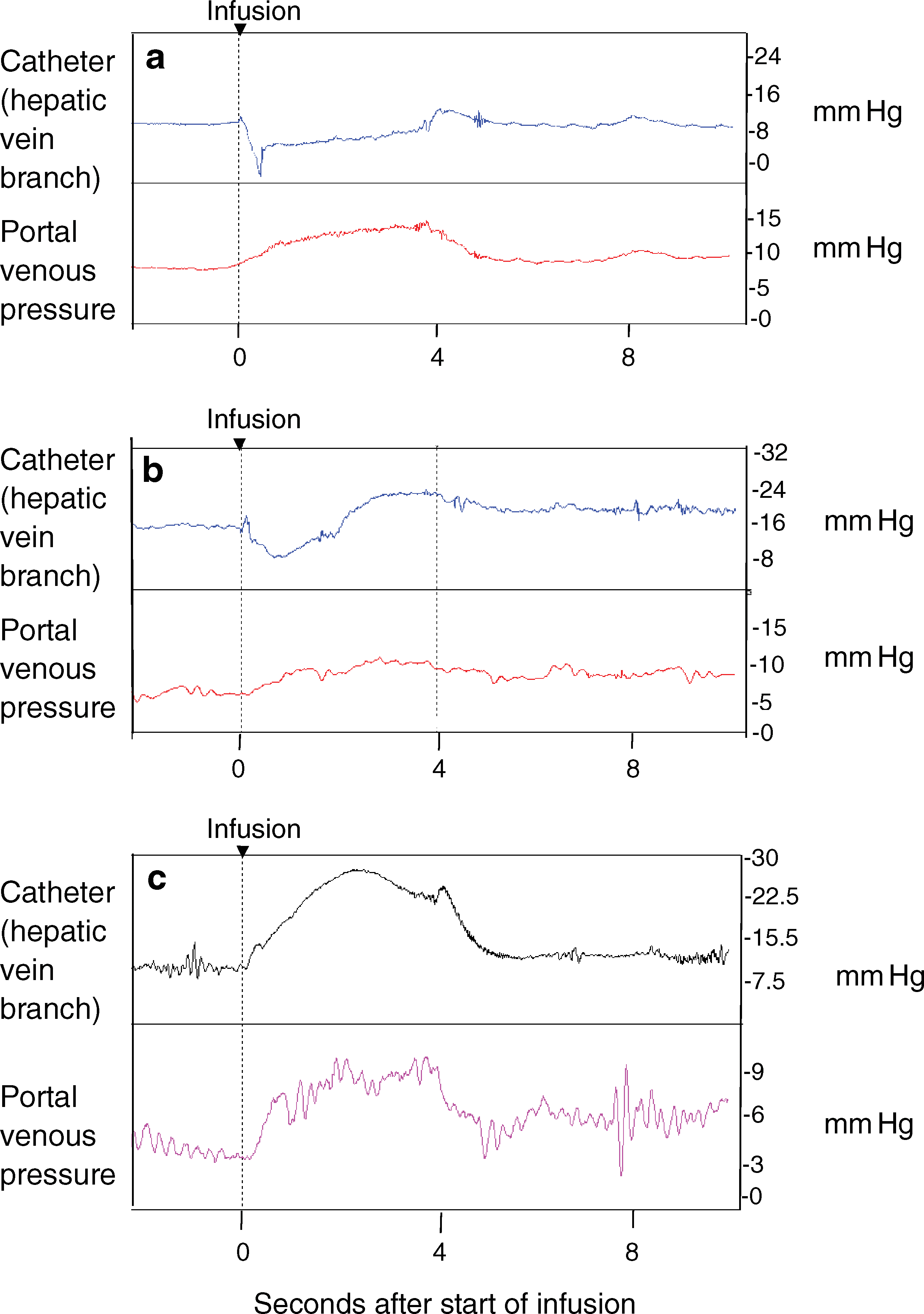
Vascular pressures during hydrodynamic delivery into a hepatic vein branch without outflow obstruction (“minimally invasive” technique). Pigs weighing 28–38 kg were given 200 ml at 50 ml/sec (3 liters/min) into a branch of the hepatic vein, using a catheter introduced via the right internal jugular vein. Targeted liver segments were ∼200 g, representing ∼20% of the liver mass (see Table 1 for details). The pressures shown are (
Pig 45 is the only pig in this group that received DNA. Highest expression from five biopsy specimens of the perfused segment was 2 × 104 relative light units/mg (Table 1), which is ∼1% of the levels obtained in the rat (Sawyer et al., 2007, 2008) and approximately those seen in the pig using the IVC segment approach (Fabre et al., 2008).
Hydrodynamic delivery via a hepatic vein branch, with total obstruction of venous outflow from the liver
Eastman et al. (2002) attempted to introduce an element of outflow obstruction to the minimally invasive technique by total obstruction of venous outflow from the liver with a balloon in the IVC. However, this was technically difficult in rabbits, as mentioned above. In the pig, we took advantage of the fact that the left and middle hepatic veins drain into the IVC cranial to the right hepatic vein, separated by a distance of ∼1–1.5 cm in ∼30-kg pigs (Fig. 3a). The technique is illustrated in Fig. 3b. An occlusion balloon catheter is introduced via the right internal jugular vein and positioned in the IVC opposite the point of entry of the left/middle hepatic veins. A perfusion catheter is then introduced into the infrarenal IVC and advanced so that the tip is opposite the point of entry of the right hepatic vein. A tie is placed around the IVC (cranial to the renal and suprarenal veins). The balloon is inflated 1 min before hydrodynamic delivery. This blocks all venous outflow from the liver and permits gene delivery via the right hepatic vein. Fluoroscopy (Fig. 3c) illustrates the effectiveness of the procedure, with the occluding balloon protruding into the terminal parts of the left and middle hepatic veins and perfusion exclusively via the right hepatic vein. An alternative approach is to place forceps across the suprahepatic IVC, occluding the left and middle hepatic veins, as illustrated in Fig. 3d.
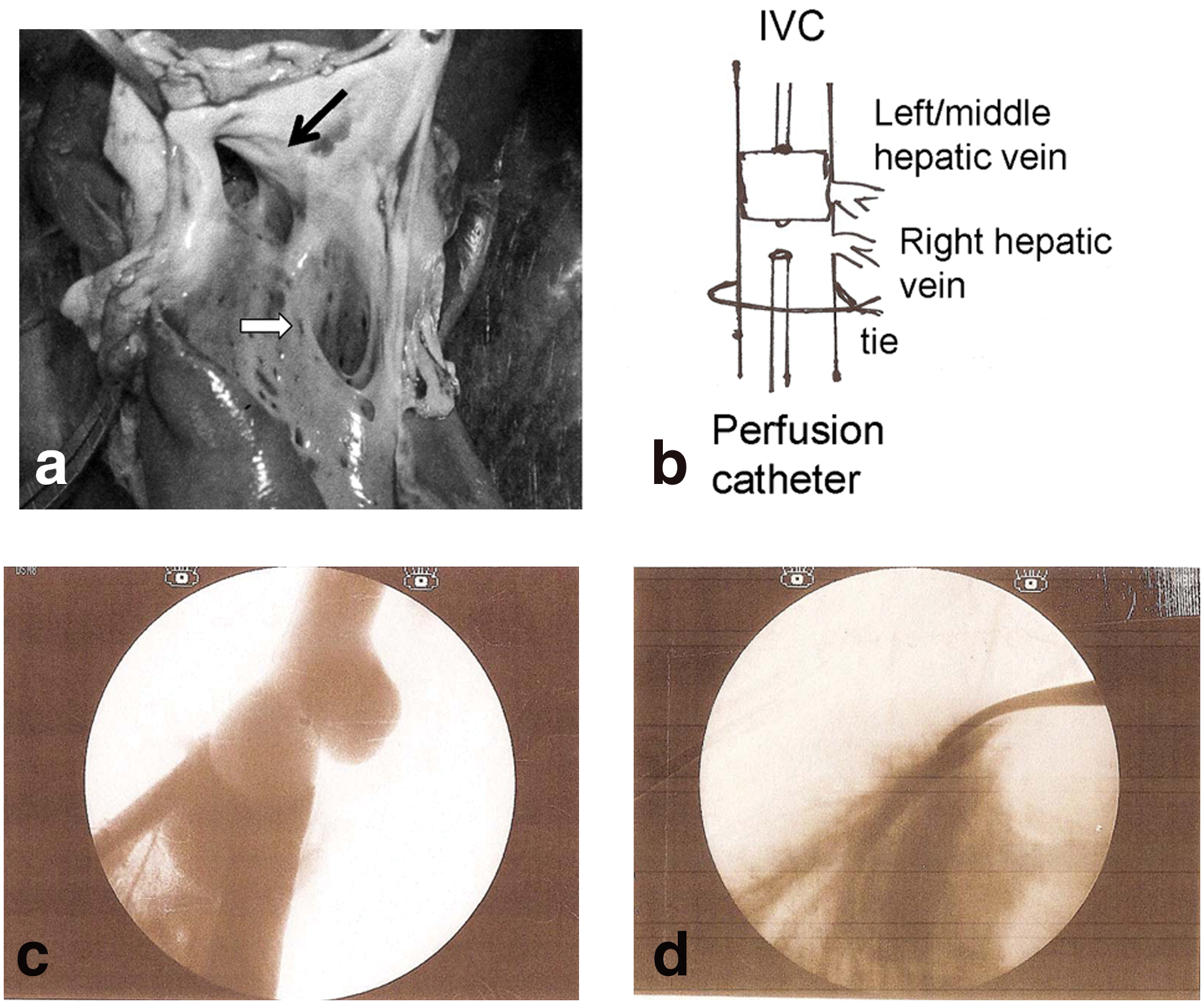
Anatomy of left/middle and right hepatic veins permits selective obstruction and selective retrograde hydrodynamic gene delivery. (
The right hepatic vein generally drains the right lateral lobe, part of the right median lobe, and the caudate lobes, which amount to ∼30–40% of the liver mass. We delivered volumes corresponding to 0.5% of the body weight at 25 ml/sec (Supplementary Table S1; Supplementary Data are available online at
Gene delivery was poor (Table S1), ∼100-fold less than the IVC segment approach (Fabre et al., 2008).
Hydrodynamic delivery via a portal vein branch, with selective outflow obstruction from the targeted segment (our “optimal” technique)
We prefer to use clamps rather than balloons wherever possible, because of the uncertainty regarding integrity of the seal provided by balloons, especially when the vasculature is pressurized and dilates (Eastman et al., 2002; Yoshino et al., 2006). In humans, after mobilization of the liver, the hepatic veins can be directly clamped, making outflow obstruction on the hepatic venous side a clinically attractive option. Thus our preferred option is delivery of the DNA solution using a balloon perfusion catheter on the portal side and direct clamping of the draining hepatic vein.
Our approach in the pig model is to target the right part of the median lobe (∼25% of liver mass). The perfusion catheter (13 French in ∼30-kg pigs) is introduced via the splenic vein, thereby avoiding the need to clamp the portal vein (Fig. 4a). Selective obstruction of the right hepatic vein is achieved by inserting a balloon catheter into the infrahepatic IVC and positioning the balloon at the point of drainage of the right hepatic vein (Fig. 4b; also refer to Fig. 3a). With this technique, portal venous flow occurs through the left side of the liver (see the direction of the arrows in Fig. 4), and gut congestion and ischemia are avoided. Moreover, although venous return from the kidneys and lower body is obstructed by the balloon in the IVC, venous return from the gut, spleen, and pancreas is maintained. This maintains reasonable venous return to the heart and minimizes cardiovascular effects. Note that in humans the right hepatic vein is amenable to direct clamping, thereby avoiding the need for an occlusion balloon in the IVC.
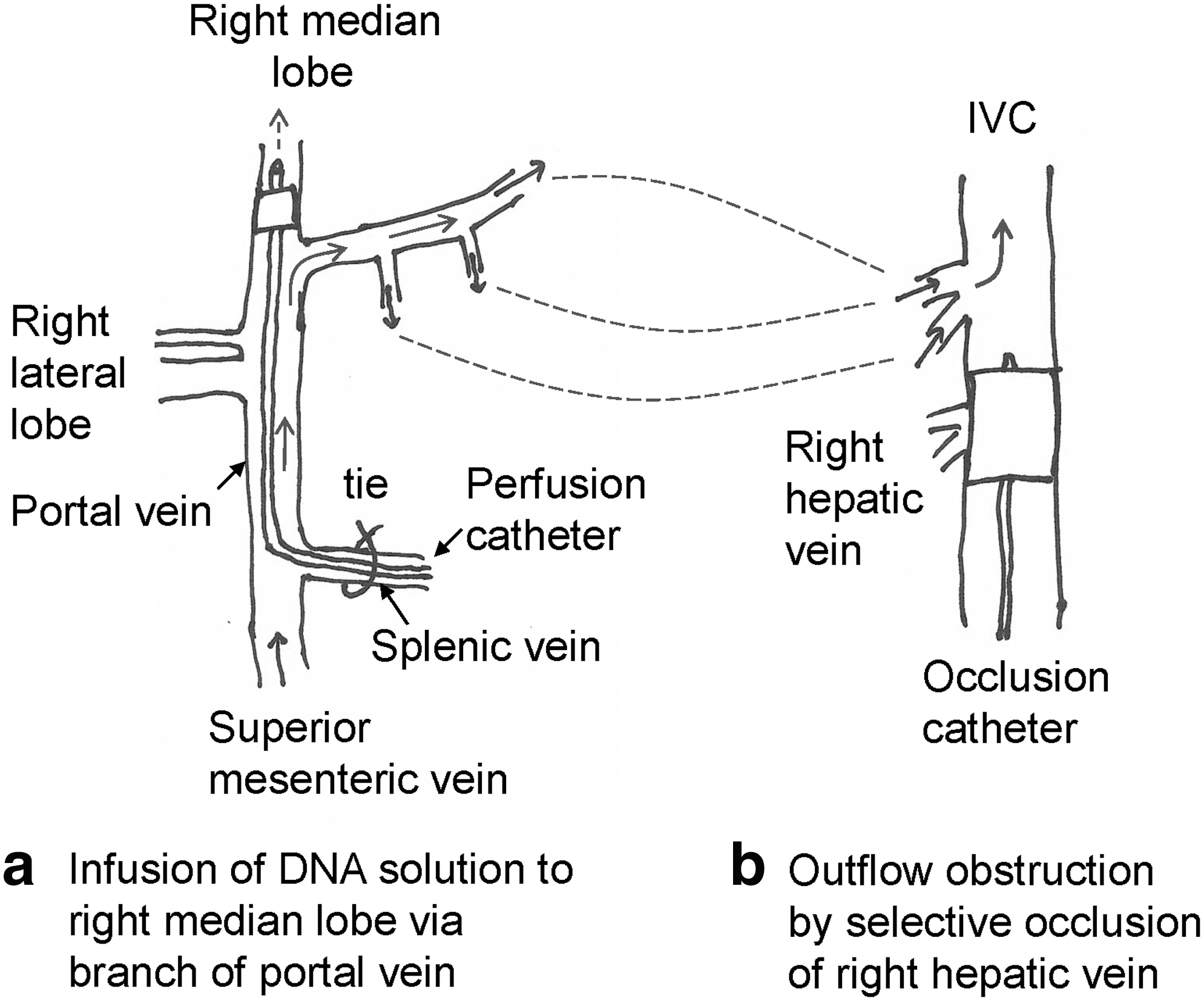
Diagrammatic illustration of the optimal technique for pressurization of a target liver segment for hydrodynamic gene delivery. (
Two hundred milliliters of DNA solution was delivered at 50 ml/sec in three pigs, and 200 ml of saline alone was delivered to a fourth pig (for pressure measurements only) (Table 2). Unlike the previous groups, the perfusion pressure rose very sharply (Fig. 5) and reached levels not seen in the absence of outflow obstruction. This demonstrates, for the first time, that selective pressurization of individual liver segments is possible. This is an important finding because there is no anatomical correspondence between inflow and outflow of blood for the various anatomical segments of the liver. The portal vein and hepatic artery branch together to supply the same segments of liver, but the terminal branches of the hepatic veins drain adjacent segments (Williams, 1995) (Fig. S1). Pressure conduits will therefore exist at the periphery of the isolated perfused segment (Sawyer et al., 2009).

Perfusion pressures during hydrodynamic gene delivery to the right median lobe with selective outflow obstruction of the right hepatic vein. Pigs weighing 30–35 kg were given 200 ml at 50 ml/sec to the right median lobe via a perfusion catheter inserted via the splenic vein into the portal vein branch supplying the right median lobe. Outflow of the DNA solution was obstructed by an occlusion balloon catheter placed in the IVC at the point of drainage of the right hepatic vein (see Fig. 4). The perfusion pressures were measured from a pressure line at the tip of the perfusion catheter. (
Measured at day 1, at time of harvest.
Assuming right median lobe represents 25% of the liver mass.
The recording apparatus did not record pressures above these values.
The liver was not weighed in this pig.
Saline without DNA was administered, to record pressure measurements only.
RLU, relative light units.
In the pig illustrated in Fig. 5a, the portal pressure rose from a baseline of 13.3 mm Hg to 76.2 mm Hg. A second pig showed a very similar picture, with the portal pressure rising from a baseline of 21.1 mm Hg to a plateau of 97.3 mm Hg. These pressure plateaus were a consequence of the apparatus settings not recording pressures above those shown. Thus they represent minimum pressures during the infusion. In the third pig (pig 44) (Fig. 5b), the pressure rose sharply from a baseline of 12.2 mm Hg to 37.7 mm Hg in 0.1 sec and then rose slowly to 103.9 mm Hg by the end of the infusion. This abrupt change in the rate of increase in pressure could reflect the elastic properties of the liver segment, possibly in association with venting of the DNA solution via the peripheral conduits discussed above, or partial loss of seal on the inflow or outflow balloon once the blood vessels began to swell from the raised vascular pressure. The pressures in the fourth pig (pig 37) (Fig. 5c) were unexpectedly complex. It is difficult to explain this pattern, but it could represent loss of the balloon seal (at either the perfusion or occlusion catheter) at 0.2 sec, as the pressure rose and the liver vasculature swelled, with re-establishment of the occlusion at 2.7 sec as the intrahepatic vascular pressure fell and the vasculature became less swollen.
The cardiovascular response to the procedure was marginal but was consistent with access of the DNA solution to the general circulation prior to release of clamps, presumably via the peripheral conduits mentioned above (Fig. S2).
Hydrodynamic delivery to the right median lobe via a branch of the portal vein, with total vascular isolation of the liver
We wished to evaluate the effects of relatively small volumes of DNA solution in a totally closed system. Kamimura et al. (2009) have reported a similar experiment, but using a branch of the hepatic vein for DNA delivery. However, as total vascular isolation of the liver has major cardiovascular complications and is unlikely to be used for gene delivery in the clinic, we present these data as supplementary information (Supplementary Text, Table S2, and Figs. S3 and S4).
Discussion
In this study we demonstrate two critical issues for regional hydrodynamic gene delivery to individual segments of the liver. First, without outflow obstruction, intrahepatic vascular pressure is either marginally raised or negative as a consequence of the rapid flow of the DNA solution through the liver without resistance (Bernoulli's principle). This is in spite of using large volumes per unit weight of liver (∼1 ml/g of targeted liver tissue) and rapid flow rates (>10 ml/min/g of targeted liver tissue). Consequently, minimally invasive techniques (e.g., Khorsandi et al., 2008), which involve DNA infusion into a hepatic vein branch without outflow obstruction, are unlikely to be effective for hydrodynamic gene delivery. For clinical application, hydrodynamic gene delivery to a segment of the liver will require surgical access to the abdomen, either open abdominal surgery or laporoscopic surgery, which represents a significant surgical intervention for the patient.
Second, we demonstrate for the first time that it is in fact possible to pressurize an individual segment of liver by selective obstruction of outflow from that segment. This is a critical point because of the lack of concordance between inflow and outflow of blood in the liver, as mentioned above (Fig. S1). It is anatomically impossible to completely block hepatic venous outflow from a targeted segment of liver because of pressure conduits at the periphery of the targeted segment. However, from our studies it is clear that the resistance offered by these peripheral vessels is sufficient to permit pressurization.
Whether or not pressurization is sufficient for hydrodynamic gene delivery in humans will depend on other factors (e.g., liver compliance [Fabre et al., 2008]) and whether or not liver size per se influences effectiveness of the hydrodynamic approach is not known (Sawyer et al., 2009). However, this can be determined only empirically. A particularly important practical issue is the efficacy of the balloon seal during the rapid infusion and buildup of pressure, with consequent increases in vessel diameter. This has been reported to be a problem by other groups in the pig model (e.g., Yoshino et al., 2006). As we state above, vascular clamps give an absolutely secure seal and can be used clinically on the hepatic veins for outflow obstruction. For delivery of the DNA solution, the perfusion catheter will be too far advanced into the liver to permit clamping or ligating, so a balloon seal will be required. Optimal location of the balloon using fluoroscopic techniques will be essential.
Kamimura et al. (2009) have reported much higher levels of reporter gene expression in the pig liver, in comparison with our studies. However, they harvested livers 4 hr after gene delivery, as opposed to the more usual 24 hr, and it is known that maximum gene expression following hydrodynamic delivery of plasmids to the liver occurs in the first few hours after delivery. For example, Liu et al. (1999) showed in their original study that luciferase levels fall 2,400-fold between 8 hr and 3 days after delivery, with most of the decline occurring in the first 24 hr. Kamimura et al. (2009) also used a different expression plasmid, which can make a major difference. The crucial thing is that in their hands reporter gene expression in liver is ∼100-fold lower in pigs when compared with mice, which is precisely the same as we have found (Fabre et al., 2008). Kamimura et al. (2009) used total vascular isolation of the liver to raise intrahepatic vascular pressure. However, this has major cardiovascular consequences and causes congestion and ischemia of the gut. Interestingly, in their studies, whether or not the liver was pressurized made no difference to the levels of reporter gene expression. Similarly, in our study, reporter gene expression seems unrelated to the intrahepatic vascular pressures generated. However, it is crucial to remember that the levels of reporter gene expression in the pig livers were very low by rodent standards. From the earliest studies (Liu et al., 1999; Zhang et al., 1999) it has been known that the efficacy of hydrodynamic gene delivery in rodents falls sharply with marginal reductions in volumes and/or flow rates (i.e., the relationship between reporter gene expression and hydrodynamic parameters is not linear). It is possible that the very low levels of reporter gene expression observed in the pig involve mechanisms of cellular uptake unrelated to the hydrodynamic effect.
A critical aspect of our optimized technique is that we insert the perfusion catheter into the splenic vein and then via the portal vein into the portal vein branch to the right part of the median lobe. A second critical point is that we selectively occluded the right hepatic vein. There is thus no obstruction of the portal circulation (which flows freely through the left side of the liver) and consequently no gut ischaemia and minimal reduction in venous return to the heart. In the pig model, obstruction of the right hepatic vein was obtained by the judicious placement of a balloon in the IVC. In humans, the hepatic veins can be directly clamped, removing the need for balloon catheters in the IVC. Finally, excellent pressurization with this technique was obtained with 200 ml of DNA solution in 30–35-kg pigs (0.6–0.7% of body weight), thus avoiding the cardiovascular problems of acute volume overload (Sawyer et al., 2007). We believe that this technique is therefore applicable in the clinical setting.
Footnotes
Acknowledgments
This work was funded by the U.K. Department of Health, the Welton Foundation, the Rosetrees Trust, and Dr. K. Ravindranath (Global Hospitals, India). We would like to thank Mr. Peter Matthews of Medrad for generously providing the syringe pumps and Mr. Steve Clarke of Kimal for advice and provision of catheters and high pressure tubing.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.