
Abstract
The use of stem cells for tissue regeneration and repair is advancing both at the bench and bedside. Stem cells isolated from bone marrow are currently being tested for their therapeutic potential in a variety of clinical conditions including cardiovascular injury, kidney failure, cancer, and neurological and bone disorders. Despite the advantages, stem cell therapy is still limited by low survival, engraftment, and homing to damage area as well as inefficiencies in differentiating into fully functional tissues. Genetic engineering of mesenchymal stem cells is being explored as a means to circumvent some of these problems. This review presents the current understanding of the use of genetically engineered mesenchymal stem cells in human disease therapy with emphasis on genetic modifications aimed to improve survival, homing, angiogenesis, and heart function after myocardial infarction. Advancements in other disease areas are also discussed.
Introduction
MSCs offer several benefits that make them ideal therapeutic agents for regenerative medicine (Pittenger et al., 1999; Boyle et al., 2010; Griffin et al., 2010; Salem and Thiemermann, 2010). An overview is shown in Fig. 1. MSCs can give rise to a variety of cell types, such as bone cells (osteocytes), cartilage cells (chondrocytes), fat cells (adipocytes), as well as blood, brain, and nerve cells and cardiac and skeletal muscle cells. In addition, MSCs are easily isolated, and can be greatly expanded ex vivo without loss of phenotype or differentiative capacity. Moreover, they are easily transfectable and amenable to genetic modification in vitro. Once transplanted, they are endogenously recruited and home to sites of inflammation and injury. Importantly, MSCs are also immunologically inert and, therefore, transplantation into an allogeneic host may not require immunosuppression (Rasmusson et al., 2003; DelaRosa and Lombardo, 2010). Furthermore, MSCs are easily obtained from a variety of tissue sources. Although most studies have been performed with bone marrow-derived MSCs, these cells have also been identified in cord blood, adipose tissue, muscle, cartilage, and skin (Rebelatto et al., 2008). It has been postulated that MSCs from these different sources may have diverse therapeutic properties (Rebelatto et al., 2008; van der Bogt et al., 2009). Last, as MSCs can be extracted from adult tissue, they do not pose the same ethical concerns as embryonic stem cells.
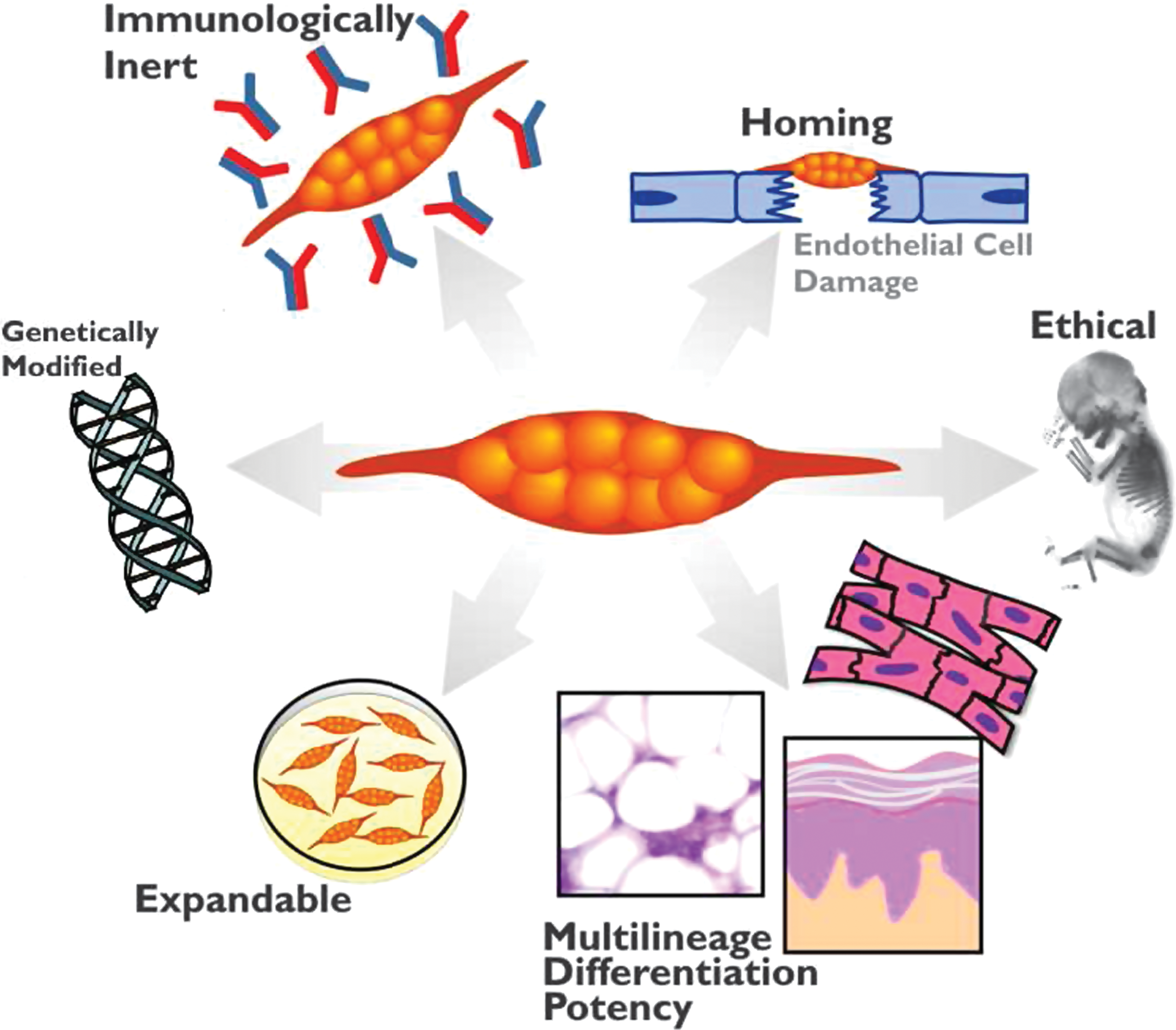
Advantages of mesenchymal stem cell (MSC) therapy. The cellular characteristics of MSCs are important for their therapeutic uses. These characteristics include being easily isolated and manipulated ex vivo, being immunologically inert, homing into the injury area, and having multilineage differentiation capacity. Genetic engineering of MSCs is aimed at improving their survival and engraftment as well as enhancing their repair mechanisms.
Despite tremendous advancements, major unresolved issues about their therapeutic application still exist. Injected MSCs suffer from poor survival and engraftment into the host tissue; in addition, in vitro culturing conditions can affect MSC pluripotency as well as expression of homing receptors (Wagner and Ho, 2007). In some cases, such as when transplanted into cardiac tissue, MSCs do not show high efficiency of transdifferentiation into functional cardiomyocytes (Noiseux et al., 2006). More importantly, there is limited knowledge about the optimal time and mode of MSC administration and there are concerns that MSC treatment has only marginal and transient effects. Meyer and colleagues found, in patients who underwent successful percutaneous coronary intervention for acute myocardial infarction (MI), an improvement in left ventricular function 6 months after intracoronary transfer of autologous bone marrow cells (Wollert et al., 2004). However, follow-up studies showed that there was no sustainable improvement at the long-term mark (Meyer et al., 2009; Schaefer et al., 2010).
It is evident therefore that these issues need to be addressed if MSCs are to be used in the clinical setting. Moreover, the precise mechanisms underlying mesenchymal stem cell “plasticity,” or pluripotency, as well as the full spectrum of the molecular processes involved in their therapeutic capacity, are not yet fully understood. For instance, after transplantation, MSCs can participate in tissue repair not only by direct transdifferentiation but also by reducing cell damage and activating endogenous mechanisms of tissue regeneration. An overview of these mechanisms is shown in Fig. 2. The ability therefore to understand and/or regulate these processes could be important to the replacement and repair of diseased tissue.
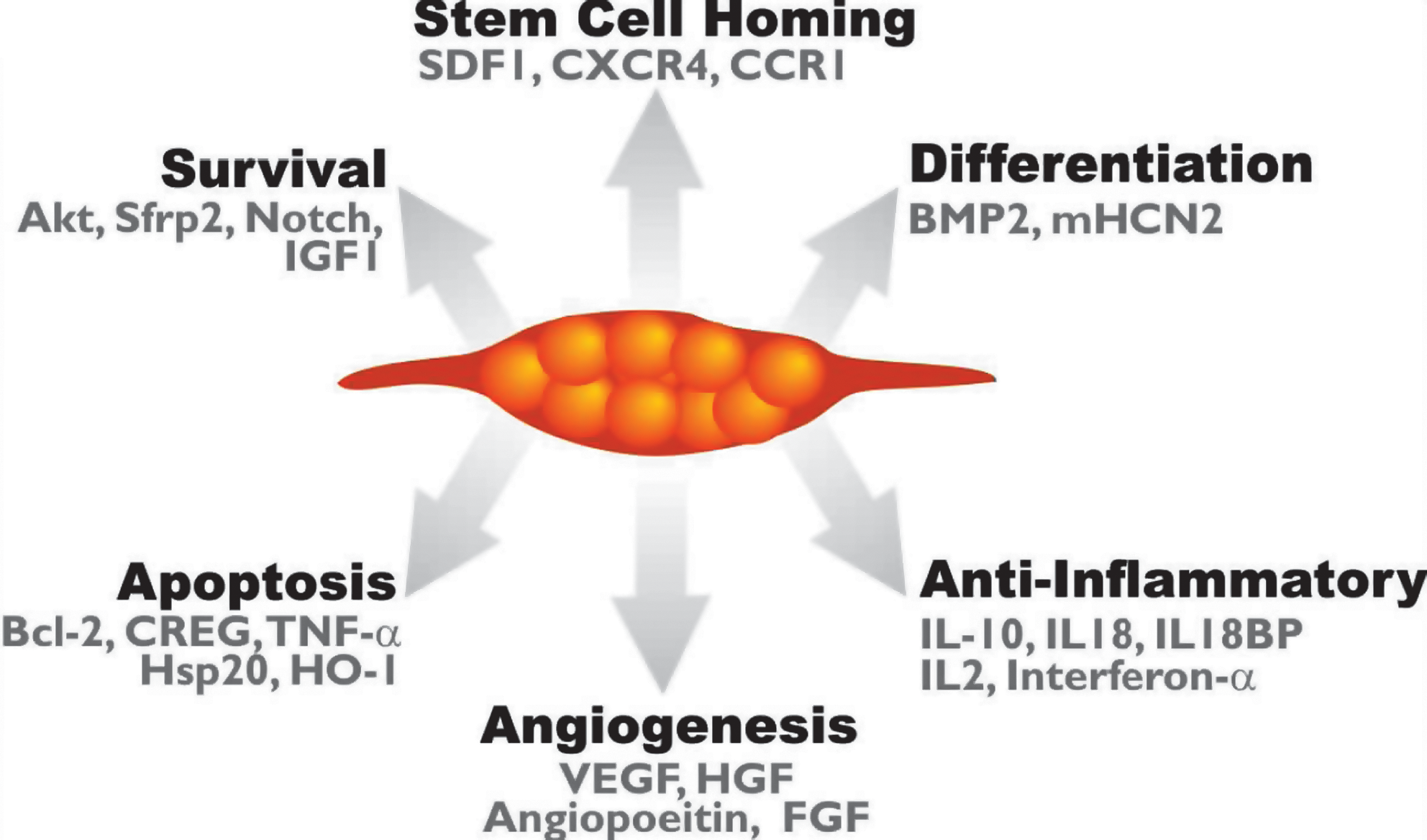
Genetic modification of MSCs enhances their therapeutic potential. Genetic modification of MSCs is aimed toward enhancing different cellular process such as MSC survival after transplantation, homing, differentiation, angiogenesis, and antiinflammatory effects. Some of the genetic targets used for this purpose are highlighted.
In the light of this, various researchers have sought to improve MSC therapy. One emergent technology is the genetic engineering of MSCs to express proteins that improve their ability to be therapeutic agents; this avenue has generated much interest because of the positive results arising from these studies (Kode et al., 2009; Griffin et al., 2010). Engineering of MSCs has improved their ability to home to sites of disease, and to promote their survival and engraftment as well as their differentiation. Moreover, as MSCs often exert their beneficial effects through the release of paracrine factors, genetic modification can also contribute significantly to the MSC-mediated paracrine effects (Gnecchi et al., 2005, 2008).
This review aims to outline the current status of our knowledge about the genetic modifications being explored as a means to improve the MSC therapy of human diseases, focusing on cardiovascular diseases. Summaries for other disease areas are also presented.
Cardiovascular Disease
Cardiovascular disease is one of the major causes of death in the world. This is due to the series of unwanted changes such as cell death, mechanical and electrical dysfunction, changes in heart structure, and scarring that follow a heart attack (Yellon and Hausenloy, 2007; Frangogiannis, 2008). MSCs have been reported to contribute to cardiac repair by promoting neovascularization, protecting the myocardium from ischemic cell death, promoting reparative processes, and enhancing cardiomyocyte regeneration (Gnecchi et al., 2008; Nesselmann et al., 2008). At present, the challenges for MSC cardiac therapy lie in improving their survival and engraftment into the injured myocardium as well as enhancing their capacity to promote cardiac regeneration (Balsam et al., 2004; Caplice and Deb, 2004; Murry et al., 2004; Dimmeler et al., 2005). These objectives have formed the basis for the majority of published material on the genetic engineering of MSCs.
Survival
It is evident that the majority of injected MSCs die within several hours of delivery (Hofmann et al., 2005; Hou et al., 2005; Freyman et al., 2006). Therefore methods to improve their survival will be important therapeutically.
In that context, multiple genetic engineering strategies have been applied to stem cells to increase their cell viability both in vitro and in vivo (Tang et al., 2005). For instance, overexpression of fibroblast growth factor (FGF)-2 improved the viability of MSCs implanted into injured myocardium, increasing expression of cardiac markers such as cardiac troponin and the calcium channel CaV2.1 as well as promoting angiogenesis (Song et al., 2005).
MSCs exposed to hypoxic conditions before transplantation also show a higher survival rate. Multiple lines of evidence suggest that this is due in part to upregulation of survival proteins such as Akt (Hu et al., 2008; Rosova et al., 2008; Y.L. Tang et al., 2009). Work in our laboratory has explored the use of MSCs genetically engineered to overexpress Akt, using a viral approach as a means to improve MSC survival in the injured myocardium. Injection of these modified MSCs (Akt-MSCs) into the heart after MI showed that they had a higher survival rate in the ischemic heart and resulted in major improvements in cardiac function (Mangi et al., 2003; Gnecchi et al., 2005). Further research revealed that Akt-MSCs promoted their therapeutic effects by releasing paracrine signals that support the survival of organ cells, as well as by initiating angiogenesis (Mangi et al., 2003; Gnecchi et al., 2005). Further validation of this paracrine model came from in vivo experiments using a rat coronary occlusion model, where administration of concentrated conditional medium from Akt-MSCs had an effect similar to that following administration of Akt-MSC cells, reducing infarct size and cardiac cell apoptosis (Gnecchi et al., 2005). Additional work from our laboratory identified several genes that were overexpressed by Akt-MSCs, leading potentially to the secretion of vascular endothelial growth factor (VEGF), basic fibroblast growth factor (bFGF), hepatocyte growth factor (HGF), insulin-like growth factor (IGF)-1, and secreted frizzled-related protein-2 (Sfrp2) (Mirotsou et al., 2007). In this study, we identified Sfrp2 as one of the key paracrine factors released by Akt-MSCs and playing a critical role in the survival of ischemic cardiac myocytes (Mirotsou et al., 2007). The protective effects of Sfrp2 involved the canonical Wnt3a pathway (Zhang et al., 2009). Importantly, Alfaro and colleagues have shown that intramyocardial injection of MSCs overexpressing SFRP2 acts to reduce infarct size by promoting MSC engraftment, and improves cardiac function and vascular density (Alfaro et al., 2008).
In addition, overexpression of a single heat shock protein, Hsp20, in MSCs, using a viral approach, improved their survival under ischemic conditions. Akt, VEGF, FGF-2, and IGF-1 are substrates of Hsp20, and in Hsp20-MSCs these proteins are refolded, with their survival and angiogenic effects restored after a stress episode (X. Wang et al., 2009). The expression of heat shock proteins is increased during cellular stress events, playing a role in refolding damaged proteins and so restoring their function; as such they are key contributors to cell survival. Conditioned medium from Hsp20-MSCs improved the survival of cardiomyocytes under oxidative stress. As might be expected from these results, injection of Hsp20-MSCs into rat hearts after MI promoted cardiac repair and function. In addition to restoring the function of Akt, Hsp20 overexpression in MSCs increased the ratio of B cell lymphoma (Bcl)-2 to Bax, which acted to inhibit apoptosis of these cells (X. Wang et al., 2009).
Antiapoptosis
Whereas some groups have taken the approach of engineering MSCs to express proliferative proteins such as Akt, others have considered whether modulating the expression of proteins known to play important roles in the cellular pathways governing apoptosis would be beneficial. To date, Bcl-2 (W. Li et al., 2007), cellular repressor of EA1-stimulated genes (CREG) (Deng et al., 2010), heme oxygenase-1 (Zeng et al., 2008a,b), and survivin (Fan et al., 2009) have all been used with the aim of improving cell survival and resistance to apoptosis.
Bcl-2 is a key antiapoptotic protein, well known for its inhibitory role in the oligomerization of proapoptotic proteins Bax and Bak, inhibits mitochondrial outer membrane permeabilization (Tsujimoto et al., 1984; Korsmeyer, 1999; Chipuk and Green, 2008). Overexpression of Bcl-2 in adult rat bone marrow-derived MSCs improved their survival, aiding heart tissue repair and organ recovery after MI (W. Li et al., 2007). Bcl-2-MSCs have increased expression of VEGF, which in itself promotes angiogenesis. Although in vivo therapy using Bcl-2-MSCs has been shown to be beneficial there are questions regarding their safety as Bcl-2 overexpression underpins leukemia development (Tsujimoto et al., 1984).
Cellular repressor of EA1-stimulated genes (CREG) is an inhibitor of apoptosis (Deng et al., 2010), and its overexpression in MSCs improves survival of the stem cells. The beneficial effects appear to be mediated by Akt activation and degradation of p53. Increased Akt activity in these modified MSCs promotes the expression of VEGF, which in turn enhances proliferation and angiogenesis (Deng et al., 2010).
Heme oxygenase (HO-1) is involved in the oxidative cleavage of heme and is known to be protective against apoptosis (Idriss et al., 2008). The protein itself could have therapeutic potential to treat heart disease (Liu et al., 2006, 2007). Interestingly, MSCs engineered to overexpress HO-1 improved organ recovery and function, and decreased ventricular remodeling after transplantation in injured rat hearts. Overexpression of HO-1 improved the survival of the injected MSCs and increased levels of secreted VEGF and bFGF as compared with unmodified cells (Zeng et al., 2008a,b). In addition, decreased expression of the proapoptotic protein Bax and reductions in the levels of proinflammatory molecules tumor necrosis factor (TNF)-α, interleukin (IL)-1β, and IL-6 were also observed (Zeng et al., 2008a,b). Moreover, MSCs in which HO-1 was transiently overexpressed also showed enhanced antiapoptotic and antioxidative properties leading to enhanced repair of the myocardium (Tsubokawa et al., 2010). Additional experiments have taken the idea of HO-1 overexpression further by producing MSCs that overexpress both HO-1 and inducible nitric oxide synthase (iNOS). The authors of this study showed that these modified MSCs improved repair of the heart after MI; furthermore, they suggested that inhibition of either protein completely removed the protective action of MSCs (Chabannes et al., 2007).
TNF-α is a proapoptotic and proinflammatory molecule with an important role in the detrimental effects of heart failure. The TNF-α receptors, TNFR1 and TNFR2, have different roles. TNFR1 promotes apoptosis, decreasing proliferation; on the other hand, TNFR2 promotes proliferation and survival (Zeller et al., 2009). A study has shown that virus-induced overexpression of TNFR:Fc, a fusion of TNFR and immunoglobulin Fc, by MSCs removes the detrimental activity of TNF-α and improves heart recovery and function. This was due in part to reduced expression and release of proinflammatory molecules such as IL-6, TNF-α, and IL-1β (Bao et al., 2008).
Stem cell homing
Chemokines are a group of secreted cytokines that induce chemotaxis in nearby cells (Richmond, 2010). After injury damaged cells secrete chemokines that act as attractants to recruit immune and stem cells to start the process of repair. Therefore another option that can be used to improve MSC therapy is to enhance homing of the MSCs to the injured myocardium (Chavakis et al., 2008).
Stromal cell-derived factor (SDF)-1 and its receptor CXC chemokine receptor-4 (CXCR4) are important mediators of stem cell recruitment after myocardial infarction or ischemia (Zaruba and Franz, 2010). Levels of SDF-1 correlate with rehabilitation of patients after coronary events (Hu et al., 2007; B.C. Lee et al., 2009) and increased engraftment of MSCs overexpressing IGF-1 was found to be dependent on SDF-1 (Haider et al., 2008). Penn's group found that MSCs expressing SDF-1 had beneficial effects on heart function after acute myocardial infarction, in part through cardiac myocyte preservation (Zhang et al., 2007). Although this study could not detect any cardiac regeneration from endogenous stem cells, further work did show the recruitment of small cardiac myosin-expressing cells, which despite being unable to differentiate into mature cardiac myocytes within 5 weeks of myocardial infarction did appear capable of depolarizing (Unzek et al., 2007). SDF-1 also appears to promote the differentiation of MSCs into endothelial cells (J. Tang et al., 2009b), which may involve dipeptidylpeptidase IV (Zaruba et al., 2009; Zaruba and Franz, 2010).
One of the issues associated with using SDF-1 overexpression as therapy is that the expression of its receptor, CXCR4, in progenitor cells is low (Penn, 2009; Y.L. Tang et al., 2009). Indeed, MSCs overexpressing CXCR4 home to the damaged infarct region of myocardium in greater numbers than do unmodified MSCs (Cheng et al., 2008).
C-C chemokine receptor type 1 (CCR1) is a G protein-coupled receptor involved in the recruitment of immune cells to site of inflammation (Kitamura and Taketo, 2007; Zernecke et al., 2008). This receptor is not expressed on MSCs; however, its ligands are significantly upregulated in the injured myocardium (Ip et al., 2007; Shimizu et al., 2009). Our laboratory has demonstrated that when overexpressed in MSCs, CCR1 increased MSC migration induced by chemokines as well as protected the cells against apoptosis in vitro (Huang et al., 2010). In addition, CCR1-modified MSCs injected into the myocardium after coronary ligation were found to accumulate in the damaged area in greater numbers than did unmodified cells. This was also associated with an improvement in cardiac function as shown by reduced infarct size, improved cardiomyocyte survival, and a denser capillary network. (Huang et al., 2010).
Angiogenesis
Neovascularization is the process whereby new blood vessels are formed. This highly regulated process is known to involve more than 20 separate proteins (Asahara et al., 1995) and is an important physiological response after MI that allows the heart to recover. Various groups have shown that MSCs secrete proangiogenic factors (Kinnaird et al., 2004a,b; Gnecchi et al., 2005, 2008). Indeed, both in vitro and in vivo models have shown that MSCs can enhance new blood vessel growth (Kinnaird et al., 2004a). Porcine autologous MSCs, when injected after chronic ischemia, showed enhancement of angiogenesis (Zhou et al., 2009). Increasing the secretion of angiogenic factors by MSCs by genetic engineering has been linked to improvements in cardiac function via enhanced new blood vessel growth.
One of the most extensively characterized angiogenic factors is VEGF. Because transplanted heart cells overexpressing VEGF increased capillary density in the border zone of a myocardial scar (Yau et al., 2001), researchers have attempted to repeat this finding with MSCs. Transplantation of MSCs overexpressing VEGF inhibited progression of left ventricular hypertrophy induced by chronic pressure overload in swine. This was associated with significant angiogenesis and improved heart function (Wang et al., 2006). In addition, after injection into mouse ischemic hind limbs, VEGF-modified MSCs promoted angiogenesis and limb retention with concomitant reduced muscle degeneration and tissue fibrosis (Yang et al., 2010). Furthermore, MSCs overexpressing a human form of the VEGF gene increased angiogenesis in the infarct region after acute MI (Gao et al., 2007; Matsumoto et al., 2005). Additional beneficial effects on infarct size and heart function, such as ejection fraction and E wave/A wave ratio, were also observed (Matsumoto et al., 2005; Gao et al., 2007). The most important member of the VEGF family is VEGF-A, of which 16 splice variants are currently known. A paper by Lin and colleagues raises the possibility that the therapeutic potential of VEGF-expressing MSCs can be modulated by the splice variant employed (Lin et al., 2008). The authors of this study expressed three VEGF-A splice variants, VEGF120, VEGF164, and VEGF188, in MSCs and measured the effects on proliferation, differentiation, and survival. Whereas all three variants strongly promoted MSC proliferation there were also clear differences. VEGF120 and VEGF188 were strongly associated with amplification of growth factor and cytokine expression, whereas VEGF164 was linked to genes involved in endothelial differentiation. In addition, VEGF188 preferentially facilitated MSC osteogenesis and enhanced cell death arising from serum starvation (Lin et al., 2008). In addition to the important role that VEGF plays in promoting angiogenesis it appears that this growth factor can also promote homing of stem cells. The protein acts to recruit cKit+ CD31+ progenitor cells to damaged myocardium in a process involving phosphatidylinositol-3-kinase (PI3K) and Akt (Wragg et al., 2008; J. Tang et al., 2009a; Zisa et al., 2009).
Other angiogenic factors have also been successfully employed for MSC genetic engineering. Hepatocyte growth factor-overexpressing MSCs injected into a rat heart after an infarct increased the density of capillaries and reduced the area of damage (Duan et al., 2003). Similarly, MSCs expressing angiopoietin have protective effects in both cerebral (Onda et al., 2008) and myocardial (Sun et al., 2007; Chen et al., 2009) models. In the cerebral artery occlusion model intravenous delivery of angiopoietin-expressing MSCs increased angiogenesis around the border of the lesion but had little or no effect on lesion volume (Onda et al., 2008). By combining overexpression of angiopoietin and Akt, these modified MSCs not only increased angiogenesis but also simultaneously inhibited apoptosis (S. Jiang et al., 2006). Three months after injection a significant number of these cells had survived and differentiated into mature muscle cells, producing fibers that were aligned and electrically coupled to the host muscle (Shujia et al., 2008).
The secretion of paracrine factors is not the only mechanism by which MSCs could promote angiogenesis. Newly formed blood vessels are stabilized by MSCs acting as pericytes. A clear link between MSCs and pericytes has been demonstrated by the finding that perivascular cells from several human organs expressed MSC markers (Caplan, 2008; Crisan et al., 2008). Migration towards pepsin- and papain-digested extracellular matrix (ECM) suggests that these pericytes/MSCs can home to sites of injury (Crisan et al., 2008). These studies open the door for future possibilities in engineering MSCs to promote new blood vessel growth.
Antiinflammatory action
Inflammatory processes arising in the heart after MI underpin the formation of scar tissue, which acts to degrade heart function. Unmodified MSCs themselves appear to activate antiinflammatory pathways. Burchfield and colleagues have shown that the beneficial effects of MSCs are partly dependent on the secretion of IL-10, an antiinflammatory cytokine (Burchfield et al., 2008). Engineering MSCs such that they inhibit post-MI inflammatory mechanisms would therefore have clear benefits.
One of the proinflammatory cytokines released from damaged myocardium is IL-18 (Venkatachalam et al., 2009). This cytokine promotes cell death in MSCs and limits their secretion of VEGF. IL-18 may therefore partly explain why the effects of MSCs are limited in vivo. IL-18-binding protein (IL-18BP), the naturally occurring inhibitor of IL-18 activity, decreases the severity of inflammation in response to injury; MSCs overexpressing IL-18BP were protected from cell death. Intramyocardial injection of IL-18BP-expressing MSCs improved various parameters of heart function and decreased infarct size in a coronary ligation model when compared with normal MSCs (M. Wang et al., 2009).
Cardiac pacemakers
Cardiac pacemaker cells initiate the beating cycle of the heart. It is important that implanted MSCs that differentiate into cardiomyocytes develop pacemaker potential and synchronize with preexisting tissue. In that context, human mesenchymal stem cells (hMSCs) transfected with murine hyperpolarization
Neurological Disease
Unmodified MSCs have been used as a potential therapy for various neurological disorders including Parkinson's (Studer et al., 1998), Huntington's (Armstrong et al., 2000), stroke (Modo et al., 2002; Zhao et al., 2002; Chen et al., 2003; Kurozumi et al., 2004), and multiple sclerosis (Akiyama et al., 2002). Studies using genetically modified MSCs for the treatment of these diseases are currently limited; however, there is plenty of potential for modification, especially by increasing differentiation and paracrine effects.
MSCs possess neuron markers such as glial fibrillary acidic protein (GFAP), neuron-specific enolase (Mareschi et al., 2006), nestin, and Tuj-1 (Tondreau et al., 2004). Multiple reports have indicated that MSCs can develop neuronal morphology and expression patterns, using simple treatments such as cyclic AMP (cAMP) (Deng et al., 2001; Kim et al., 2006), epidermal growth factor (EGF)/bFGF (Kim et al., 2006), 2-mercaptoethanol (Mareschi et al., 2006), or specialized media (Woodbury et al., 2000; Tondreau et al., 2004). It has been suggested that more complex regimens that mimic the regulated stepwise differentiation of precursors into adult neurons are probably required for the generation of neuronal cells from MSCs. In that context, Dezawa and colleagues were able to produce neuronal cells from MSCs by a step-by-step approach (Dezawa et al., 2004). MSCs transfected with the intracellular domain of Notch, a key regulator of the terminal differentiation of neurons and glial cells, displayed neural stem cell markers. Subsequent treatment with glial cell line-derived neurotrophic factor (GDNF) gave rise to cells that could produce dopamine and were tyrosine hydroxylase positive. Importantly, transplantation of these cells into a rat model of Parkinson's gave rise to functional recovery in the animals (Dezawa et al., 2004). In addition, MSCs transfected with the intracellular domain of Notch1 became SB623 neuroprogenitor cells. These SB623 cells derived from Notch1-MSCs, when injected, engrafted and subsequently promoted dense fiber formation with serotonin immunoreactivity in a rodent model of Parkinson's disease (Dezawa et al., 2004). More recently, the same group has shown that these effects were at least partially mediated by enhanced secretion of GDNF (Glavaski-Joksimovic et al., 2009).
Cancer
Factors released by MSCs have antitumor properties reducing the proliferation of glioma, melanoma, lung cancer, hepatoma, and breast cancer cells (Maestroni et al., 1999; Nakamura et al., 2004; Qiao et al., 2008). However, it is the ability of MSCs to migrate to cancer tissue (Nakamizo et al., 2005; Menon et al., 2007; Xin et al., 2007; Loebinger et al., 2009) that has generated the most interest, as this homing ability of MSCs suggests they may be useful as delivery agents to target tumors (Loebinger et al., 2009).
The cancer tissue-homing mechanism employed by MSCs is currently unknown. Cancer cells do secrete SDF-1, which may indicate a role for the SDF-1:CXCR4 axis (Orimo et al., 2005; Dwyer et al., 2007). However, as mentioned earlier, CXCR4 expression is low in MSCs. Considering that there are nearly 20 chemokine receptors that have been reported to be expressed on MSCs (Honczarenko et al., 2006; Ponte et al., 2007; Ringe et al., 2007), elucidating the mechanism is likely to be challenging.
At present, MSCs have been used as delivery agents for a variety of molecules that can inhibit tumor growth. Bone marrow-derived MSCs expressing interferon (IFN)-α reduced proliferation of transformed cells via an increase in apoptosis in a melanoma lung metastasis model (Ren et al., 2008a). IFN-α delivery by MSCs, which leads to accumulation of cells in S phase and increased apoptosis, was found to be beneficial in several cancer models including glioma (D.H. Lee et al., 2009), melanoma (Studeny et al., 2002), and prostate cancer (Ren et al., 2008b). Similarly, IFN-α released from MSCs inhibited leukemia cell proliferation in vitro (Li et al., 2006). Delivery of IL-12 and IL-18 has been adopted in order to recruit T cells and natural killer cells. IL-12-engineered MSCs prevented metastasis into the lymph nodes and other internal organs as well as increased tumor cell apoptosis (Chen et al., 2008). Similarly, MSCs transduced with a vector carrying IL-18 have been shown to be protective against glioma (Xu et al., 2009). Antiglioma effects have also been observed with MSCs expressing IL-2 (Nakamura et al., 2004), which is known to increase T cell proliferation. MSCs engineered to express CX3CL1, a strong T cell chemoattractant, reduced metastasis to the lung after injection of cancer cells (Xin et al., 2007). These results are interesting in light of the strong immunosuppressive effects of MSCs, with T and B cells arrested in G0/G1 (Glennie et al., 2005; Corcione et al., 2006) and regulatory T cells activated (Wolf and Wolf, 2008). TNF-related apoptosis-inducing ligand (TRAIL), a member of the tumor necrosis factor family, is a highly selective protein able to induce apoptosis in cancerous cells but leaving normal cells unaffected (Wiley et al., 1995; Pitti et al., 1996). TRAIL-expressing MSCs induced apoptosis in various cancer cell lines in vitro and cleared lung metastases in about 40% of mice compared with 0% in controls (Loebinger et al., 2009). MSCs have also been employed to deliver adenovirus; replication of the virus within malignant cells destroyed the tumors (Komarova et al., 2006; Hakkarainen et al., 2007; Stoff-Khalili et al., 2007).
However, the use of MSCs in cancer therapy is not without its disadvantages; MSCs have been implicated in promoting the growth of certain cancers. For example, IL-6 and CCL5, proteins produced by MSCs, have been shown to increase the growth and metastasis of breast cancer cells, respectively (Karnoub et al., 2007; Sasser et al., 2007).
Bone Formation
MSCs have also been used in the treatment of bone disease and repair. The incidence of bone fractures failing to heal is dependent on the fracture site and can be as high as 20% (Undale et al., 2009; Chanda et al., 2010). Loading of MSCs onto structural scaffolds has been shown to be useful in the repair of fractures. MSCs loaded onto ceramic cylinders and implanted into rat femora formed bone by 8 weeks (Kadiyala et al., 1997). These hydroxyapatite ceramic cylinders have also successfully generated bone in canine and sheep (Bruder et al., 1998; Kon et al., 2000).
There has been an increase in the use of genetically engineered MSCs to enhance osteoblast lineage commitment, bone formation, fracture repair, and spinal fusion. MSCs expressing bone morphogenetic protein-2 (BMP2), transforming growth factor (TGF)-β1, latent membrane protein (LMP)-1, IGF-1, or growth differentiation factor (GDF)-5 have enhanced cartilage, bone, and tendon repair (Nixon et al., 2007). Specifically, MSCs overexpressing IGF-1 improved tendon healing in a collagenase model of tendinitis (Schnabel et al., 2009). Ultimate tensile strength was not directly increased by MSC injection; however, tendon architecture was improved in the early period after injury, and appropriate choice of integrating nonimmunogenic vectors should further improve growth factor residence time in the tendon.
In addition, a number of studies have also demonstrated that genetic modification of MSCs by enforced expression of either telomerase reverse transcriptase (TERT) (Shi et al., 2002; Gronthos et al., 2003; Abdallah et al., 2005), Wnt4 (Chang et al., 2007), or BMP (Jiang et al., 2005; X. Jiang et al., 2006) enhanced their proliferation capacity and improved bone formation and repair of bone defects, with no adverse effects on the endogenous tissue in response to viral transfer or tumor development (Arthur et al., 2009).
Furthermore, MSCs genetically modified to express BMP2, with a further modification to express α4-integrin to enhance homing of the cells, increased bone mineral density and mineral content. This was due in part to enhanced recruitment of endogenous progenitors involved in bone formation (Kumar et al., 2010). Similar results have been obtained in other studies (H. Li et al., 2007; Tai et al., 2008). An in-depth study of the properties of BMP2-MSC-induced bone formation indicated that this new bone is structurally similar to that of native adult bone (Tai et al., 2008).
More recently, in vitro studies have shown that MSCs genetically engineered to overexpress the longer, VEGF188 isoform facilitated BMP7-mediated MSC osteogenesis. Whether these cells will also show enhanced osteogenic properties in vivo remains to be tested (Lin et al., 2008).
Renal Failure
Many kidney disorders involve both ischemic/inflammatory and immunologic injury. Therefore cell-based therapies such as those using MSCs that function through multiple mechanisms and have the potential to target the inflammatory and immunologic pathways have been considered a clinically relevant solution, in contrast to pharmacologic agents that target only a single event or pathway in the pathophysiology of a given disease. Experimental evidence suggests that administering exogenous mesenchymal stem cells during acute and chronic kidney injury may improve functional and structural recovery of the tubular, glomerular, and interstitial kidney compartments (Asanuma et al., 2010). Eliopoulos and colleagues have generated murine MSCs genetically modified to secrete erythropoietin (EPO) and they were tested to determine whether they can improve anemia in mice with mild to moderate chronic renal failure (Eliopoulos et al., 2006). Their data showed that these cells led to an increase in hematocrit when admixed in a bovine collagen matrix and implanted by subcutaneous injection in mice. More recently, the same group has demonstrated that delivery of MSCs overexpressing a combination of EPO and IGF-1 resulted in enhanced hematocrit elevation, as well as improved cardiac function, compared with animals receiving MSCs expressing EPO alone (Kucic et al., 2008).
Future Directions
MSC therapy has been shown to be beneficial in the treatment of a diverse range of diseases. However, problems with poor survival, engraftment, and differentiation have hampered the routine use of MSCs in the clinic. Genetic engineering of MSCs has the potential to overcome these challenges. Research on engineered MSCs revealed that many of the effects attributable to MSC therapy appear to be mediated by secreted proteins. This opens the door to the possibility of directly administering these proteins in a clinical setting without the need for injecting cells. Nevertheless, genetically engineered MSCs and their applicability as therapeutic agents remains an emerging and developing field. Additional research into the streamlining of protocols for the optimal isolation and expansion of those cells in vitro, as well as for their administration in vivo, is currently in progress, and has been enhanced by the use of novel biomaterials (Kobayashi et al., 2004; Mouw et al., 2007; Zhao et al., 2007; Potier et al., 2010). Importantly, the genetic modification of MSCs, such as the use of a new generation of viral vectors or the use of RNA silencing, alone or in combination with traditional gene modification protocols, is currently under investigation (Nixon et al., 2007). As more becomes understood about MSC biology the engineering of MSCs will become more refined and useful in the treatment of a greater number of diseases. Progress in these areas might make the prospect of “off-the-shelf” MSC biotherapeutic products achievable.
Footnotes
Acknowledgments
The authors express appreciation to Steven Conlon (Department of Pathology, Duke University) for his artistic contribution to the illustrations in this review. Research conducted by our group and mentioned in this review was supported by National Heart, Lung, and Blood Institute grants RO1 HL35610, HL81744, HL72010, and HL73219 (to V.J.D.); the Edna Mandel Foundation (to V.J.D. and M.M.); and the Leducq Foundation (to V.J.D.). M.M. is also supported by an American Heart Association National Scientist Development Award (10SDG4280011).
Author Disclosure Statement
No disclosures.