
Abstract
Many preclinical gene therapy studies use a reporter gene to evaluate vector design and performance in mouse models of human disease. Unfortunately, most commonly used reporter genes are immunogenic in mice, which confounds accurate evaluation of vector function. In previous studies, we showed that the murine secreted alkaline phosphatase (mSEAP) gene functions well as a simple and sensitive reporter gene in mice. In this study, we have used rational design to enhance mSEAP performance. The majority of native mSEAP remains attached to the outer surface of the cell through glycan phosphatidylinositol linkage; removal of the carboxy-terminal tail of mSEAP resulted in a dramatic enhancement of release of the protein into cell culture medium and into mouse plasma in whole animal experiments. We increased the heat stability of mSEAP through mutation of a key residue in the crown domain of the protein (H451E), thus allowing us to reduce endogenous, background AP activity through heat inactivation for enhanced sensitivity. We show that these alterations in mSEAP result in enhanced performance in tissue culture and mouse studies. Taken together, these data illustrate that mSEAP is a sensitive, nonimmunogenic reporter for preclinical mouse studies.
Introduction
There are a number of reporter genes in common use, and each of the reporter proteins has various advantages and disadvantages regarding ease or sensitivity of quantification, utility for in vivo imaging, and so on. The most popular reporter genes are perhaps β-galactosidase, firefly luciferase, human placental secreted alkaline phosphatase (hSEAP), and green fluorescent protein. However, all of these reporter proteins elicit strong immune responses in immunocompetent mice (Yang et al., 1996; Klavinskis et al., 1999; Parks et al., 1999; Maelandsmo et al., 2005; Finn et al., 2009). Human α1-antitrypsin (AAT) has also been used as a reporter gene in many studies, but it is immunogenic in some mouse strains, such as BALB/c, C3H/J, and CBA/J (Morral et al., 1997; Schowalter et al., 1999), and must be assayed by ELISA, reducing the simplicity of its use. The human α-fetoprotein has been suggested as a good marker of gene transfer but, like AAT, it must be assayed by ELISA and it can be highly immunogenic (O'Neal et al., 2000). The baboon α-fetoprotein has been used successfully as a reporter gene in nonhuman primate studies (Brunetti-Pierri et al., 2006, 2009). Other reporter genes include soluble human carcinoembryonic antigen or the β subunit of human chorionic gonadotropin (Peng et al., 2002a,b). The human sodium iodide symporter is an attractive reporter gene for cancer-directed studies, because it can serve a dual purpose by allowing uptake and concentration of radioisotopes such as iodine-123 for imaging by tomography, or iodine-131 as an anticancer agent (Dingli et al., 2003; Msaouel et al., 2009). All of these reporter genes are useful, but have varying effects in long-term mouse studies in immunocompetent mice. Other factors that influence foreign reporter protein immunogenicity include the vector system and mouse strain used, as well as the route of administration (Barr et al., 1995; Jooss et al., 1998; Brockstedt et al., 1999).
We have shown that murine secreted alkaline phosphatase (mSEAP) is an excellent reporter gene in tissue culture and mouse studies (Muruve et al., 2004; Maelandsmo et al., 2005; Ross et al., 2009; Smith et al., 2009; Poulin et al., 2010). Unfortunately, most of the native mSEAP remains cell-associated due to glycan phosphatidylinositol (GPI) linkage to the cell surface (Engle et al., 1995). Thus, native mSEAP is good for analyzing cell-type expression patterns in cell culture or tissue sections, but has reduced sensitivity in studies involving sequential monitoring of AP activity in media or plasma, because only a fraction of the “secreted” protein is liberated from the cell surface. Furthermore, unlike hSEAP, mSEAP is not heat stable, and therefore transient heating to 65°C cannot be used to inactivate endogenous, background AP activity. In this study, we have modified mSEAP to enhance secretion and heat stability, thereby improving mSEAP function and utility as a reporter gene in mouse studies.
Materials and Methods
Cell and virus culture
All cell culture media and reagents were purchased from Invitrogen (Carlsbad, CA) or Sigma-Aldrich (St. Louis, MO). 293 cells (Graham et al., 1977) and A549 cells (human lung carcinoma, CCL 185; American Type Culture Collection [ATCC], Manassas, VA) were grown in minimal essential medium supplemented with 10% fetal calf serum, 2 mM
Reporter gene construction
The E1/E3-deleted Ad vector lacking a transgene has been previously described (AdRP2014; Smith et al., 2009). The E1/E3-deleted Ad viruses expressing native or modified mSEAP or hSEAP carry the reporter gene under regulation by the human ubiquitin C promoter and bovine growth hormone polyadenylation sequence. Construction of the mSEAP genes lacking sequence encoding the hydrophobic, carboxy-terminal tail (mSEAPΔ) and H451E mutation that confers increased heat resistance (mSEAPΔHR) has been previously described (Willemsen, 2009). Briefly, for mSEAPΔ, pBS-mSEAP (Maelandsmo et al., 2005) was used as a template for polymerase chain reaction (PCR) amplification with synthetic oligonucleotides designed to remove the coding region for the terminal 15 amino acid residues from the protein (Fig. 1A). In a similar manner, synthetic oligonucleotides were designed to mutate the mSEAP crown domain, which is associated with heat stability in hSEAP (Millan, 2006), to more closely resemble hSEAP (Fig. 2A). mSEAP genes were generated that contained sequence encoding either the H451E mutation or the combination of R450D and H451E; however, the later molecule had heat stability identical to that of the single H451E mutant (data not shown), and was not used in subsequent studies. Viruses carrying the various mSEAP constructs were constructed by standard techniques (Ross and Parks, 2009).
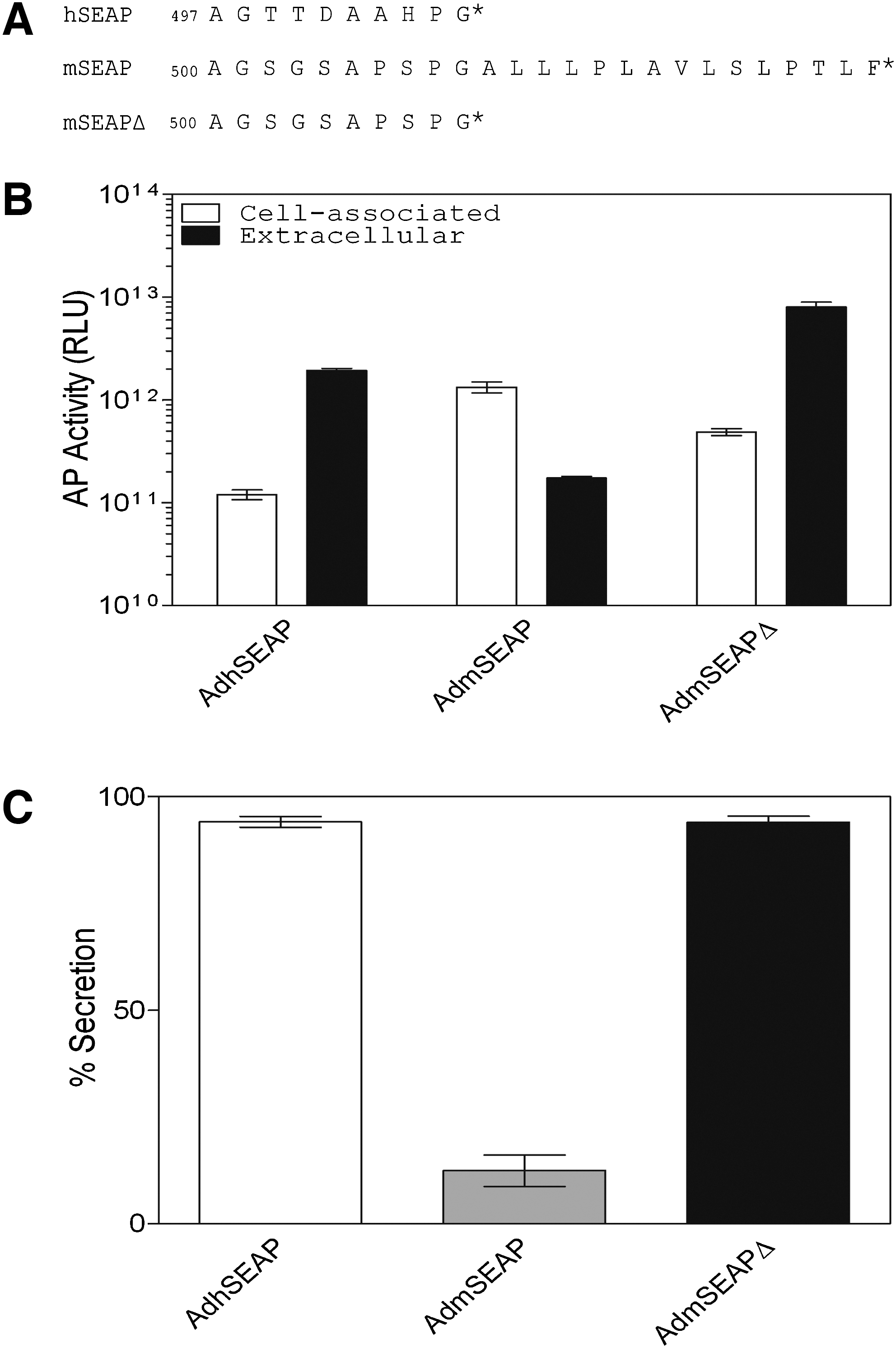
Removal of the murine secreted alkaline phosphatase (mSEAP) carboxy-terminal tail enhances protein secretion. (
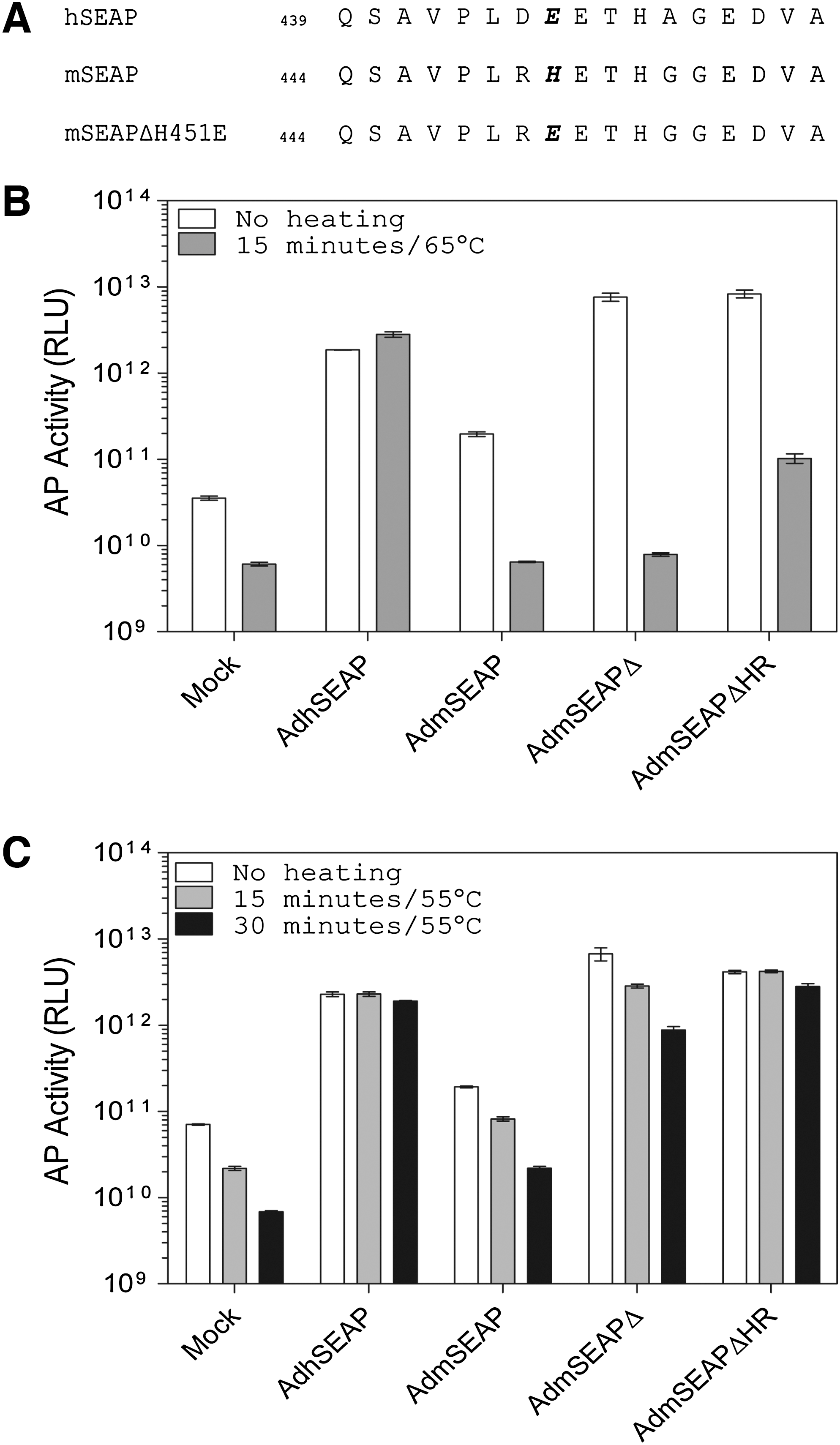
A single point mutation in mSEAP (H451E) enhances protein resistance to heating. (
Assays for SEAP activity in vitro
To monitor SEAP expression in vitro, A549 cells in 35-mm dishes were infected for 1 hr at a multiplicity of infection (MOI) of 20 with AdhSEAP, AdmSEAP, AdmSEAPΔ, or AdmSEAPΔHR. Twenty-four hours later, the medium and cells were collected separately, and the cells were lysed in reporter lysis buffer (Promega, Madison, WI). Aliquots of each were assayed for alkaline phosphatase (AP) activity with a commercially available chemiluminescence kit (Roche, Indianapolis, IN), using purified hSEAP as standard to ensure the readings were within the linear range. This kit is based on detection of alkaline phosphatase activity. Unlike hSEAP, mSEAP and its derivatives are sensitive to the AP inhibitors present in the inactivation buffer in this kit. Thus, the inactivation buffer was not included in the assays for AP activity.
Animal experiments
Eight- to 10-week-old female BALB/c mice (Charles River, Sherbrooke, QC, Canada) were injected via the tail vein with 5 × 108 plaque-forming units (PFU) (volume, 100 μl) of AdhSEAP, AdmSEAP, AdmSEAPΔ, AdmSEAPΔHR, empty vector (AdRP2014), or phosphate-buffered saline (PBS). At various times postinjection, blood samples (∼100 μl) were collected into tubes containing lithium–heparin, and then subjected to centrifugation at 5000 × g. The resulting plasma samples were stored at −80°C until assayed for AP activity. All animal experiments were approved by and performed according to the guidelines set by the Animal Research Ethics Board at the University of Ottawa (Ottawa, ON, Canada).
Detection of antibodies to SEAP and adenovirus in immunized mice
Antibodies to SEAP were detected by a combination of supershift assay and an immobilized SEAP activity assay, as previously described (Maelandsmo et al., 2005). In this assay, SEAP protein is mixed with plasma from immunized animals, the mixture is separated by nondenaturing polyacrylamide gel electrophoresis (PAGE), and the resulting gel is stained for AP activity; the presence of SEAP-reacting antibodies is detected as a shift in the migration of the SEAP protein to a more slowly migrating form. Medium samples from A549 cells infected with AdhSEAP or AdmSEAPΔHR were used as a source of AP protein. Five microliters of pooled plasma (n = 5, 1 μl each) from mice immunized with the various Ad constructs was mixed with 50 μl of medium from the infected cells, and each mixture was incubated 1 hr at 37°C. The protein–plasma mixture was mixed with an equal volume of loading buffer (62.5 mM Tris-HCl [pH 6.8], 30% glycerol, 0.5% bromophenol blue), separated by nondenaturing 10% PAGE according to standard techniques (Bio-Rad, Hercules, CA), and transferred to a polyvinylidene difluoride (PVDF) membrane (Immobilon; Millipore, Bedford, MA), as recommended by the manufacturer. The membrane was washed twice in alkaline phosphatase buffer (100 mM Tris-HCl [pH 9.5], 100 mM NaCl, 50 mM MgCl2) and incubated with 10 ml of a solution containing nitroblue tetrazolium (NBT)/5-bromo-4-chloro-3-indolyl phosphate (BCIP) (NBT/BCIP ready-to-use tablets; Roche). After sufficient color had developed, the membrane was washed twice in termination buffer (20 mM Tris-HCl [pH 7.5], 5 mM EDTA). The polyacrylamide gel was also stained to detect residual AP activity.
We also examined whether antibodies to hSEAP neutralized AP activity. Serial dilutions of purified hSEAP (protein standard from the SEAP reporter gene assay kit; Roche) were mixed with 5 μl of pooled plasma (n = 5) isolated 28 days postimmunization, and incubated at 37°C for 1 hr. The samples were then assayed for AP activity, using a chemiluminescence assay.
Antibodies to Ad were detected using plasma samples collected 42 days postinfection. Ad-infected cells were lysed in 4% sodium dodecyl sulfate (SDS) sample buffer and resolved by 9% PAGE and transferred to a PVDF membrane. After blocking for 1 hr with 5% skim milk in Tris-buffered saline (TBS-T; 150 mM NaCl, 10 mM Tris-HCl [pH 7.5], 0.1% Tween 20), the membrane was placed in a Miniblotter 28 apparatus (Immunetics, Boston, MA). Mouse plasma samples were diluted in 5% milk in TBS-T at a dilution of 1:100 and applied to the membrane for 1 hr at room temperature. As a positive control, we used a commercially available anti-Ad antibody (ab6982, diluted 1:1000; Abcam, Cambridge, UK). After three 5-min washes with TBS-T, the membrane was incubated with a horseradish peroxidase (HRP)-conjugated secondary antibody for 45 min at room temperature. After three 5-min washes in TBS-T, the membrane was incubated with a chemiluminescent substrate (Pierce/Thermo Fisher Scientific, Rockford, IL) for 5 min and exposed to X-ray film.
Results and Discussion
Removal of the C terminus of mSEAP enhances release from the cell
Most mammalian AP remains cell associated due to anchoring of the protein to the outer cell membrane through a glycan phosphatidylinositol (GPI) linkage (Engle et al., 1995), which reduces the accuracy and sensitivity of SEAP determination from medium or plasma samples. Low levels of SEAP activity in the milieu may lead to a requirement for direct measurement of cell-associated activity, thereby eliminating the possibility of sequentially assaying the same cells or animal. hSEAP was engineered for more efficient release from the cell by removal of the C-terminal 24 amino acids of the parental PLAP (placental alkaline phosphatase) protein (Berger et al., 1988). Although this truncation does not remove the aspartic acid residue involved in the GPI linkage (Micanovic et al., 1988), it sufficiently affects protein maturation and results in release of >90% of the protein into the surrounding milieu. Sequence alignment of PLAP and mSEAP did not yield significant similarity in the carboxy termini of the two proteins; thus, we chose to truncate mSEAP after a conserved proline–glycine motif that is also the site of truncation for hSEAP, and we designated the molecule mSEAPΔ (Fig. 1A). To test whether deletion of the carboxy-terminal tail of mSEAP resulted in enhanced release from cells, we infected A549 cells with AdhSEAP, AdmSEAP, or AdmSEAPΔ and examined total AP activity in cells and medium 24 hr later. As shown in Fig. 1B and C, approximately 98% of hSEAP protein is found in the medium, whereas only 15% of native mSEAP is released. Removal of the carboxy-terminal region of mSEAP results in an ∼6-fold increase in release of mSEAP into the medium, which reaches the level of secretion of hSEAP. Thus, removal of the carboxy-terminal tail allows for more efficient release of mSEAPΔ from the cell and results in higher levels of AP activity in the surrounding milieu.
Specific mutation of the mSEAP crown domain enhances heat resistance
All cells possess a certain level of endogenous AP activity. Most endogenous APs are heat sensitive, and “background” AP activity can be reduced through simple heat treatment of samples or tissues. Our preliminary studies showed that, unlike hSEAP, mSEAP and mSEAPΔ were not heat stable when heated at 65°C (Fig. 2B). Previous studies have shown that at least part of the heat stability observed for PLAP (the precursor of hSEAP) is conferred by the specific amino acid sequence of the crown domain, a flexible loop of approximately 60 amino acid residues located near the carboxy terminus of the protein (Bossi et al., 1993). Although the crown domain displays significant variability among APs, alignment of mSEAP and hSEAP showed a relatively well-conserved region of 17 amino acids (Fig. 2A). In this region, there were three amino acid differences between mSEAP and hSEAP at amino acid positions 450, 451, and 455 (using mSEAP numbering). We reasoned that the G-to-A substitution at position 455 would have minimal effect on protein stability, and thus concentrated on the other two residues. These two residues of PLAP have been studied in great detail as they contribute significantly to the function and stability of the protein (reviewed in Millan, 2006). We therefore designed two mSEAPΔ variants that contained either an H451E mutation (ultimately designated mSEAPΔHR—heat resistant—in this study) or a combination of R450D/H451E (Fig. 2A). Inclusion of the double mutation did not provide any significant advantage over H451E alone (data not shown), and thus was not pursued further.
To examine the heat stability of mSEAPΔH451E, Ad vectors expressing this protein, or control vectors, were used to infect A549 cells, the medium was collected 24 hr later, and an aliquot was heated at 65°C for 15 min before assay for AP activity (Fig. 2B). Uninfected cells showed a high basal level of AP activity that is significantly reduced by the heat treatment (∼6.7 × 1010 vs. 6.8 × 109 RLU/ml of medium, respectively). Over the same 15-min heating protocol, there was no decrease in hSEAP activity, clearly illustrating the heat stability of this protein. Both native mSEAP and mSEAPΔ showed a dramatic loss of activity when heated at 65°C for 15 min. Although mSEAPΔHR also showed a significant loss of activity after 15 min of heating, it was still clearly detectable relative to untreated cells, and retained approximately 10-fold greater activity above parental mSEAPΔ. These data suggest that the H451E mutation does impart some heat resistance, but does not reach the stability of hSEAP. We therefore examined AP activity after a heating protocol using a lower temperature of 55°C (Fig. 2C). Mock-infected cells showed an ∼10-fold decrease in AP activity after heating at 55°C for 30 min. Once again, we observed a significant loss of AP activity for both mSEAP- or mSEAPΔ-containing protein extracts (∼10-fold for each over the duration of the time course). Both hSEAP and mSEAPΔHR were stable with heating at 55°C for 30 min. Thus, the H451E mutation improves the heat stability of mSEAPΔ, and we have identified a heating protocol that significantly reduces background AP activity while leaving the mSEAPΔHR reporter protein unaffected.
Expression of mSEAPΔHR in vivo
The main advantage of mSEAP (and its derivatives) as a reporter protein is that, after vector delivery to mice, the level and duration of reporter gene AP activity can be monitored in plasma samples taken sequentially from the same mice over an extended period of time. Therefore, we next performed a series of in vivo experiments to determine whether (1) deletion of the carboxy-terminal tail from mSEAP leads to greater release of protein into plasma; (2) inclusion of the H451E mutation in mSEAP allowed us to heat-treat plasma samples to decrease endogenous, background AP activity, thereby enhancing reporter protein sensitivity; and (3) inclusion of these two modifications altered the immunogenicity of this otherwise “self” reporter protein.
BALB/c mice were injected via the tail vein with 5 × 108 PFU of AdRP2014 (empty vector), AdhSEAP, AdmSEAP, AdmSEAPΔ, or AdmSEAPΔHR (or PBS) and blood samples were removed 4 days later. The plasma was then examined for the quantity of AP activity in both unheated and heat-treated samples (Fig. 3A). Mice injected with PBS had a baseline level of AP activity of 2.6 × 108 RLU/μl of plasma, which dropped ∼3-fold after heat treatment (55°C for 30 min). Animals receiving the AdRP2014 empty vector control had an identical level of AP activity as PBS-treated animals, indicating that treatment with this dose of vector did not adversely affect liver function, as gauged by the lack of elevated levels of liver-derived AP activity in the plasma. For AdhSEAP-treated animals, there was a high level of AP activity in plasma, which was unaffected by heat treatment. We observed a ∼2-fold higher level of AP activity in the plasma of AdmSEAPΔ- versus AdmSEAP-treated animals and, as expected, the level of AP activity in the plasma declined by 3- to 4-fold on heat treatment, once again clearly illustrating that native mSEAP is not heat stable. Finally, in mice treated with AdmSEAPΔHR, there was a higher quantity of AP activity in the plasma relative to AdmSEAP-treated animals (1.3 × 109 vs. 7.5 × 108 RLU/μl plasma, respectively), and this activity was only marginally reduced by heat treatment. For comparison, the fold increase above background for AdmSEAPΔHR for nonheated and heat-treated samples was ∼5- and 12-fold, respectively, showing that the added heat stability of AdmSEAPΔHR allows for processing of the samples to enhance the dynamic range of reporter protein detection above background.

Expression of mSEAPΔ and mSEAPΔHR in vivo. (
We next examined expression of our various mSEAP constructs over a 60-day time course in mice, to determine whether our protein modifications to mSEAP affected the stability or duration of expression. Once again, BALB/c mice were injected via the tail vein with 5 × 108 PFU of AdRP2014 (empty vector), AdhSEAP, AdmSEAP, AdmSEAPΔ, or AdmSEAPΔHR (or PBS) and blood samples were removed at various times (Fig. 3B). In this experiment, all plasma samples were heat-treated for 30 min at 55°C before AP activity determination. Both PBS- and AdRP2014-treated showed an identical and consistent level of AP activity over the duration of the time course. Mice treated with AdhSEAP showed a high level of AP activity in plasma for approximately 1 week; however, there was a dramatic, ∼30-fold decline in activity to background levels by 2 weeks. Mice receiving AdmSEAP showed a 12-fold lower level of peak AP activity in plasma compared with AdhSEAP (2.5 × 108 vs. 3.1 × 109 RLU/μl plasma, respectively), and AP activity was undetectable above background by 2 weeks. Both AdmSEAPΔ and AdmSEAPΔHR showed similar kinetics of expression over the duration of the experiment, although the level of AP activity in plasma was slightly higher for AdmSEAPΔHR, likely because of its enhanced heat resistance relative to mSEAPΔ. For these two viruses, peak levels of AP activity were observed 4 days after vector delivery, and the quantity of AP was approximately 4-fold above that observed for parental AdmSEAP. After 4 days, there was a gradual decline in AP activity but the quantity of AP remained 5-fold above background at the end of the experiment (60 days after vector delivery). The dramatic difference in expression kinetics between hSEAP and mSEAPΔHR clearly illustrates that the nature of the reporter protein (self vs. non-self) can greatly affect the duration of gene expression from gene therapy vectors, which in this study were E1-deleted Ad.
Analysis of immune responses to reporter proteins and vector
Previously, we showed that expression of hSEAP was not limited by the formation of cytotoxic T lymphocytes (CTLs) directed toward cells expressing the protein, but rather was due to the formation of anti-hSEAP antibodies (Maelandsmo et al., 2005). Therefore, we examined plasma samples to determine whether antibodies were generated to our hSEAP or modified mSEAP proteins in this study. We used an immobilized SEAP activity assay for detection of anti-hSEAP or anti-mSEAP antibodies, as we have previously described (Maelandsmo et al., 2005). In this assay, SEAP protein is mixed with plasma from immunized animals, the mixture is separated by nondenaturing PAGE, and the resulting gel (or nylon membrane after transfer) is stained for AP activity. The presence of SEAP-reacting antibodies is detected as a shift in the migration of the SEAP protein to a more slowly migrating form. As expected, mice treated with AdhSEAP developed antibodies to the protein, as indicated by a “supershift” of the protein, and these antibodies did not cross-react with mSEAP protein or its derivatives (Fig. 4A). We did not detect any antibodies to mSEAP, mSEAPΔ, or mSEAPΔHR, indicating that these relatively minor modifications to the protein sequence that dramatically enhance protein secretion and stability do not affect the immunogenicity of the protein. We next examined whether the antibody generated to hSEAP neutralized protein activity in vitro: a constant amount of plasma from animals immunized with AdhSEAP was mixed with serial dilutions of purified hSEAP protein for 1 hr at 37°C, and then assayed for AP activity (Fig. 4B). We did not observe a difference in AP activity between samples containing plasma or PBS controls, suggesting that, although antibodies were formed to hSEAP, these antibodies did not specifically neutralize its activity. Thus, loss of hSEAP expression in animals was due either to antibody binding hSEAP in the blood and removal of this complex from the circulation, or antibody-mediated cytolysis of infected, hSEAP-expressing cells.
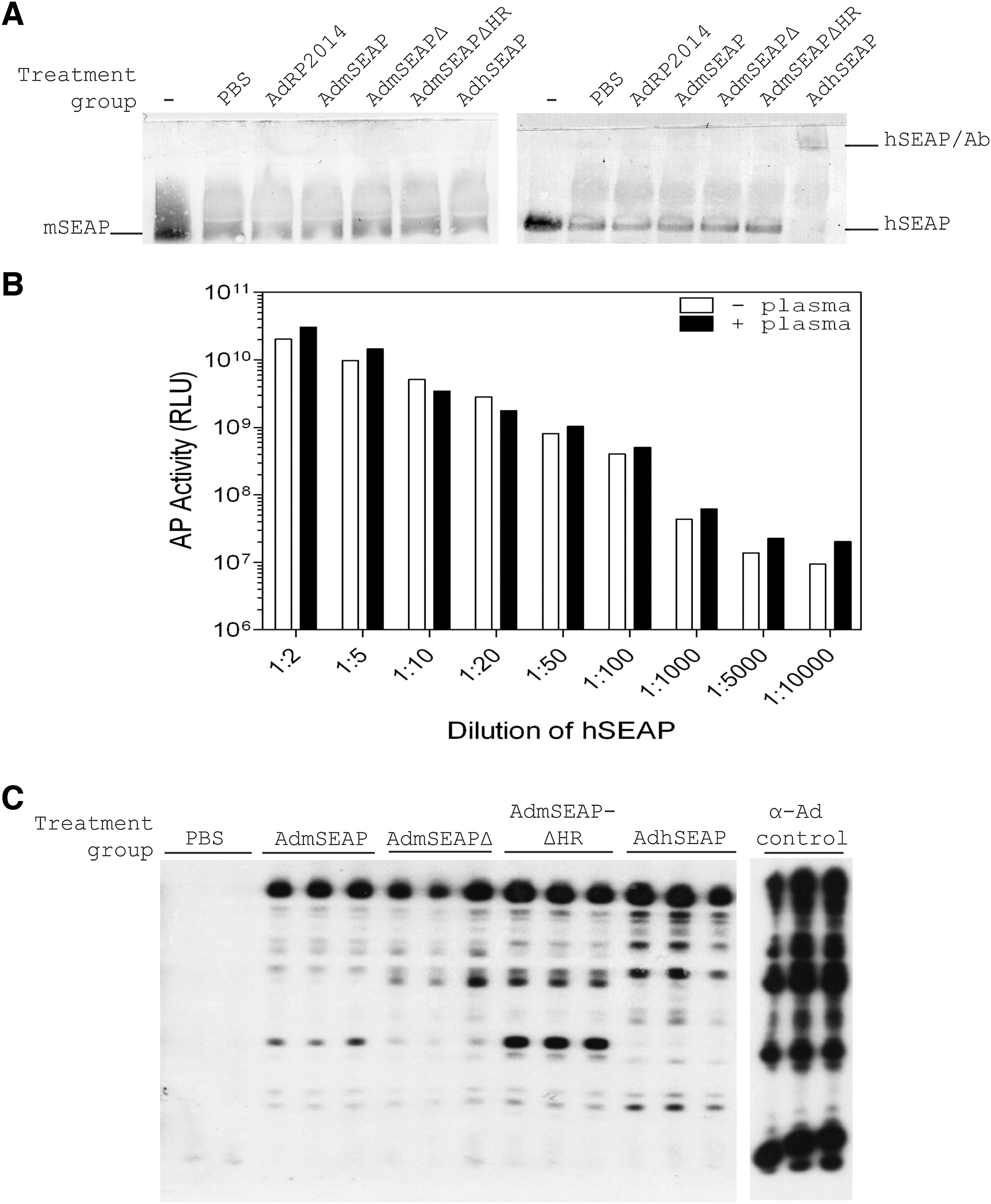
Analysis of immunological response to Ad-mediated delivery of hSEAP and mSEAP and its derivatives. (
We also analyzed the plasma samples for the presence of anti-Ad antibodies, using immunoblot analysis (Fig. 4C). As expected, all animals treated with Ad vector developed anti-Ad antibodies. These data suggest that antibodies produced against the E1-deleted Ad vector backbone may have induced some degree of antibody-mediated cytolysis of the transduced cells, contributing to the slow and gradual decline in mSEAPΔ and mSEAPΔHR expression over the course of our in vivo experiment (Fig. 3B). That Ad-infected cells can persist and continue to express transgene protein in the presence of acquired anti-vector immunity has been shown in several studies (Tripathy et al., 1996; Wadsworth et al., 1997). Thus, acquired immunity to the Ad vector was not sufficient to eliminate expression of the “self” mSEAPΔHR reporter protein, clearly showing that the persistence of vector-encoded gene expression is highly dependent on the nature of both the vector and transgene protein.
Reporter genes are ubiquitously used in research, and many have been described. These reporter proteins vary in their sensitivity or ease of use, and the ideal reporter gene for a particular study ultimately depends on the experimental system and research question being addressed. mSEAP has the advantage that it is not immunogenic in mice, allowing for long-term expression studies without the complication of anti-transgene immune responses, and it can be assayed in plasma samples from mice using a commercially available, sensitive chemiluminescence or less sensitive chromogenic assay. Unfortunately, native mSEAP is poorly released from the cell, reducing the quantity of the protein in the plasma and sensitivity of the reporter. In addition the protein is heat sensitive, so transient heating of the plasma samples cannot be used to inactivate endogenous AP. We have now modified mSEAP to increase both secretion and heat stability. The modified reporter protein, mSEAPΔHR, remains nonimmunogenic in mice, and can be used for applications requiring long-term, sequential sampling of treated mice. Thus, the optimized mSEAPΔHR should prove valuable in mouse preclinical studies directed toward optimizing vector design and function.
Footnotes
Acknowledgments
The authors thank members of the Parks laboratory and Dr. Rashmi Kothary for critical evaluation of the manuscript and helpful discussion, and Benoit Goulet, Marisa Rossi, and Kristin Willemsen for technical assistance. This work was supported by grants from the Canadian Institutes of Health Research, Muscular Dystrophy Canada, the Amyotrophic Lateral Sclerosis Society of Canada, the Jesse Davidson Foundation for Gene and Cell Therapy, and the Cancer Research Society (Canada).
Author Disclosure Statement
No competing financial interests exist.