
Abstract
Oncolytic viral therapy represents a promising strategy for the treatment of head and neck squamous cell carcinoma (HNSCC), with dl1520 (ONYX-015) the most widely used oncolytic adenovirus in clinical trials. This study aimed to determine the effectiveness of the Lister vaccine strain of vaccinia virus as well as a vaccinia virus armed with the endostatin–angiostatin fusion gene (VVhEA) as a novel therapy for HNSCC and to compare them with dl1520. The potency and replication of the Lister strain and VVhEA and the expression and function of the fusion protein were determined in human HNSCC cells in vitro and in vivo. Finally, the efficacy of VVhEA was compared with dl1520 in vivo in a human HNSCC model. The Lister vaccine strain of vaccinia virus was more effective than the adenovirus against all HNSCC cell lines tested in vitro. Although the potency of VVhEA was attenuated in vitro, the expression and function of the endostatin–angiostatin fusion protein was confirmed in HNSCC models both in vitro and in vivo. This novel vaccinia virus (VVhEA) demonstrated superior antitumor potency in vivo compared with both dl1520 and the control vaccinia virus. This study suggests that the Lister strain vaccinia virus armed with an endostatin–angiostatin fusion gene may be a potential therapeutic agent for HNSCC.
Introduction
HNSCC is an ideal model for cancer gene therapy because it generally progresses in a predictable manner from the primary site to local metastases in cervical lymph nodes, with distant metastatic disease only in the most advanced cases. HNSCC tumors are generally easily accessible and are therefore amenable to repeated biopsy and intratumoral injection.
Viruses have emerged as one of the most promising classes of vector for delivery of gene therapy to HNSCC (Harrington et al., 2005), and it was for the treatment of advanced HNSCC that China approved the world's first virus for cancer gene therapy in October 2003, a nonreplicating, E1-deleted serotype 5 adenovirus (Ad5) expressing p53 (Peng, 2005). Approval for the first replication-competent oncolytic virus also came from China in November 2005 for H101. This Ad5, in common with dl1520 (ONYX-015), has an E1B-55kD deletion, although H101 has an additional partial E3 deletion (Garber, 2006). dl1520 had previously been developed in the United States, with encouraging results when combined with conventional therapies in phase II trials (Nemunaitis et al., 2001). However, the clinical outcome of monotherapy with this virus has been disappointing.
Fundamental limitations to the efficacy of oncolytic adenoviruses have been identified, such as poor spread through tumors (Shayakhmetov et al., 2002) and rapid hepatic clearance from the circulation after intravenous delivery (Nunes et al., 1999). Adenovirus enters cells using the Coxsackie-adenovirus receptor, the expression of which is down-regulated in HNSCC (Jee et al., 2002). This led us to consider alternative viral vectors that might not be limited in the same way.
Oncolytic vaccinia virus relies mostly on its own encoded proteins to carry out replication and transcription in the cytoplasm, with few, if any, host proteins required. It is also inherently tumor selective due to the low production of interferons by tumor cells in response to vaccinia infection; in normal cells, interferons inhibit vaccinia replication (Luker et al., 2005). Vaccinia virus has a large cloning capacity and, following use in Europe as the vaccine to eradicate smallpox, a good safety profile in humans (Thorne and Kirn, 2004; Thorne et al., 2005).
We recently demonstrated that the Lister vaccine strain of vaccinia virus (VVLister) represents an attractive alternative oncolytic virus in pancreatic cancer models due to an inherently high selectivity for cancer cells, which spares normal cells both in vitro and in vivo (Tysome et al., 2009a). We sought to determine the potential of this virus for the treatment of HNSCC, since the tumor selectivity of vaccinia virus is, in part, due to the overexpression of epidermal growth factor receptor (EGFR). Vaccinia viruses express vaccinia growth factor early during replication. This is homologous to cellular EGF and, therefore, binds to the ErbB family of receptors. This results in thymidine kinase–induced nucleotide production (Stein and Staros, 2000), thereby increasing capacity for vaccinia replication (Tzahar et al., 1998). Most HNSCC tumors are known to overexpress EGFR, resulting in the recent targeting of this receptor for monoclonal antibody and tyrosine kinase inhibitor therapy (Astsaturov et al., 2008). Therefore, HNSCC represents a particularly promising model for targeted oncolytic vaccinia therapy.
Recent interest in angiogenesis inhibition as a treatment strategy for HNSCC has arisen because numerous angiogenesis promoters are released from these tumors (Saba et al., 2007) and high vascular density correlates with poor prognosis (Lopez-Graniel et al., 2001). Gene therapy using replication-competent viral vectors offers the possibility of targeted angiogenesis inhibitor delivery to tumors, providing local expression of these proteins on a continuous basis to achieve maximum efficacy (Tysome et al., 2009b). The use of angiogenesis inhibitors in combination is possible because many have a broad spectrum of activity, with low toxicity and resistance. Endostatin and angiostatin are two potent endogenous angiogenesis inhibitors that have been delivered by gene therapy for HNSCC (Li et al., 2006; Matsumoto et al., 2002). They were found to act synergistically when used in combination. This led to the development of an endostatin–angiostatin fusion gene (Scappaticci et al., 2001), which has been delivered using other viral vectors (Scappaticci et al., 2001; Raikwar et al., 2005; L. Li et al., 2006; X. Li et al., 2008). We recently demonstrated significant antitumor potency against a human pancreatic cancer xenograft model in vivo using a novel vaccinia virus expressing the endostatin–angiostatin fusion gene (Tysome et al., 2009a). Here we highlight the potential of the Lister strain of vaccinia virus for the treatment of HNSCC by demonstrating the superior efficacy of oncolytic vaccinia virus armed with the human endostatin–angiostatin fusion gene over dl1520 in a human HNSCC model.
Material and Methods
Cell lines
The HNSCC cell lines SCC4 (tongue) and SCC25 (tongue) were obtained from Cancer Research UK Central Cell services (CR-UK CCS) and were maintained in a 1:1 mix of Dulbecco's modified Eagle's medium (DMEM) and Ham's F12 supplemented with 10% heat-inactivated fetal calf serum (FCS) and 400 ng/ml hydrocortisone (Sigma-Aldrich). The HNSCC cell lines TR126 (tongue) and TR138 (larynx) were obtained from CR-UK CCS and maintained in DMEM and Ham's F12 respectively, each with 10% FCS. FaDu was obtained from the American Type Culture Collection (ATCC) and maintained in Earle's minimal essential medium with 10% FCS.
Human umbilical vein epithelial cells (HUVEC) were obtained from Cambrex and maintained in EGM-2. The African Green Monkey normal kidney cell line, CV1, was obtained from ATCC and cultured in DMEM supplemented with 10% FCS. JH293, human kidney epithelial cells transformed with the Ad5 E1A gene, were obtained from CR-UK CSS and maintained in DMEM with 10% FCS.
Viruses
The highly attenuated Lister vaccine strain (VVLister) of vaccinia virus was used as a parental virus for the construction of recombinant viruses (Denes et al., 2006), which was performed as previously described (Timiryasova et al., 2001). The nonessential Not1 restriction site, located between F14L and F15L of the VVLister (Timiryasova et al., 2001), was used for insertion of the fused human endostatin and angiostatin genes (VVhEA) or the Escherichia coli lacZ gene (VVlacZ), to serve as a control for VVhEA. Recombinant viruses were verified by Southern blot analysis and DNA sequencing. Wild-type adenovirus Ad5 was described previously (Kaufman et al., 2005) and dl1520 was originally obtained from Arnold J. Berk, UCLA, Los Angeles, California (Barker and Berk, 1987).
Viral anti-tumor potency in vitro
Cells were seeded in 96-well plates at 1 × 103 and 5 × 103 cells/well, depending on growth rates, and infected with viruses 16–18 hr later. Cell survival was determined by MTS [3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium] assay according to the manufacturer's protocol (Promega) 6 days following infection. All assays were performed at least three times, and viral dose killing 50% of tumor cells (EC50 values) were calculated as previously described (Wang et al., 2003). The ratios of EC50 values of adenovirus and vaccinia virus were determined as a measure of sensitivity to virally induced cytopathic effects, with higher ratios indicating greater sensitivity to vaccinia virus.
Viral replication in vitro
Cells were seeded in three wells of six-well plates in media with 10% FCS at 2 × 105 to 4 × 105 cells/well, depending on growth rates, and infected with 1 plaque-forming unit (PFU)/cell of vaccinia viruses or adenoviruses in medium with 2% FCS 16–18 hr later. Samples were harvested in triplicate at 24-hr intervals up to 144 hr, freeze-thawed three times, and titrated on CV1 or JH293 cells to determine the 50% tissue culture infective dose (TCID50) of vaccinia viruses and adenoviruses respectively as previously described (Wang et al., 2003).
Western blotting for detection of endostatin–angiostatin fusion gene expression
2 × 105 FaDu cells were seeded in 60-mm dishes in conditional media and infected with 1 PFU/cell VVhEA or VVlacZ in DMEM with 2% FCS after 16–18 hr. Cells and supernatant were harvested separately 24, 48, and 72 hr following infection, and supernatants were concentrated using Centricon 10 kDa columns (Millipore). Thirty micrograms of proteins were separated on precast NuPage Novex 4–12% Bis-Tris gels (Invitrogen), electrotransferred onto nitrocellulose membranes, and probed with a polyclonal rabbit antibody to human endostatin (Abcam). Immunocomplexes were detected as previously described (Wang et al., 2003).
HUVEC proliferation and tube formation
HUVEC were seeded in 96-well plates at 1 × 104 cells/well in EGM-2 and treated 16–18 hr later with 10 μg/ml of supernatant concentrated from SCC4, SCC25, or FaDu cells that had been previously infected with VVlacZ or VVhEA for 72 hr. After another 96 hr, HUVEC proliferation was determined using cell viability measured by MTS assay.
The inhibition of HUVEC tube formation was observed using the BD Biocoat angiogenesis system (BD Biosciences) according to the manufacturer's instructions. Tube formation of HUVEC treated with supernatant was compared to mock-infected HUVEC alone by counting the number of completely formed tubes per well by fluorescence microscopy.
Histopathology and immunohistochemistry
Thirty-six 4- to 5-week-old female BALB/c nude mice received 1 × 106 FaDu cells by subcutaneous implantation into their right flank. When tumors reached 4–5 mm in diameter, mice were stratified into groups of 12 with similar tumor sizes. Groups received one 100-μl intratumoral (IT) injection of 1.0 × 108 PFU of VVhEA, VVlacZ, or PBS. Groups of three mice in each cohort were sacrificed 1, 3, 10, and 17 days later, and tumors were removed, snap-frozen in isopentane, and stored at −80°C. Blood was collected into a heparinized tube and plasma stored at −80°C. All tissues were processed for histopathology and immunohistochemistry analysis for viral coat protein (1:2000 rabbit anti-vaccinia virus coat protein polyclonal antibody [MorphoSys UK Ltd.]), endostatin expression (1:250 rabbit anti-human endostatin polyclonal antibody [Abcam]), and CD31 (1:200 rat anti-murine CD31 antibody [BD Biosciences]) to determine microvessel density as described previously (Kaufman et al., 2005).
Antitumor efficacy in vivo
Thirty-five 4- to 5-week-old female BALB/c nude mice received 1 × 106 FaDu cells by subcutaneous implantation into their right flank. When tumors reached 0.4–0.5 cm in diameter, mice were regrouped (n = 7 per group) by tumor size and received 100-μl IT injections of VVlacZ (1.0 × 108 PFU), VVhEA (1.0 × 108 PFU), dl1520 (5 × 108 PFU), or PBS on days 0, 3, 6, 9, 12, and 15. Tumor volumes were estimated [volume =(width × length2 × π)/6] twice weekly until mice were sacrificed when sick or tumor volume reached 1 cm3.
Results
VVLister demonstrates better cell killing than adenovirus and replicates in human HNSCC cell lines
To demonstrate the potential of the VVLister as a therapeutic agent for human HNSCC, we compared the cytolytic potential of VVLister and Ad5 in a panel of human HNSCC cell lines in vitro (Table 1). VVLister displayed a greater potency than Ad5 in all cell lines tested, effective even against those cancer cells insensitive to adenovirus, such as FaDu. To determine whether vaccinia virus–induced cell killing was indicative of replication induction in cancer cells, replication of VVLister was confirmed in SCC4 and FaDu, the cell lines that were the most and the least sensitive to VVLister, respectively (Fig. 1). The level of viral replication was consistent with the relative cytotoxicity of the viruses.
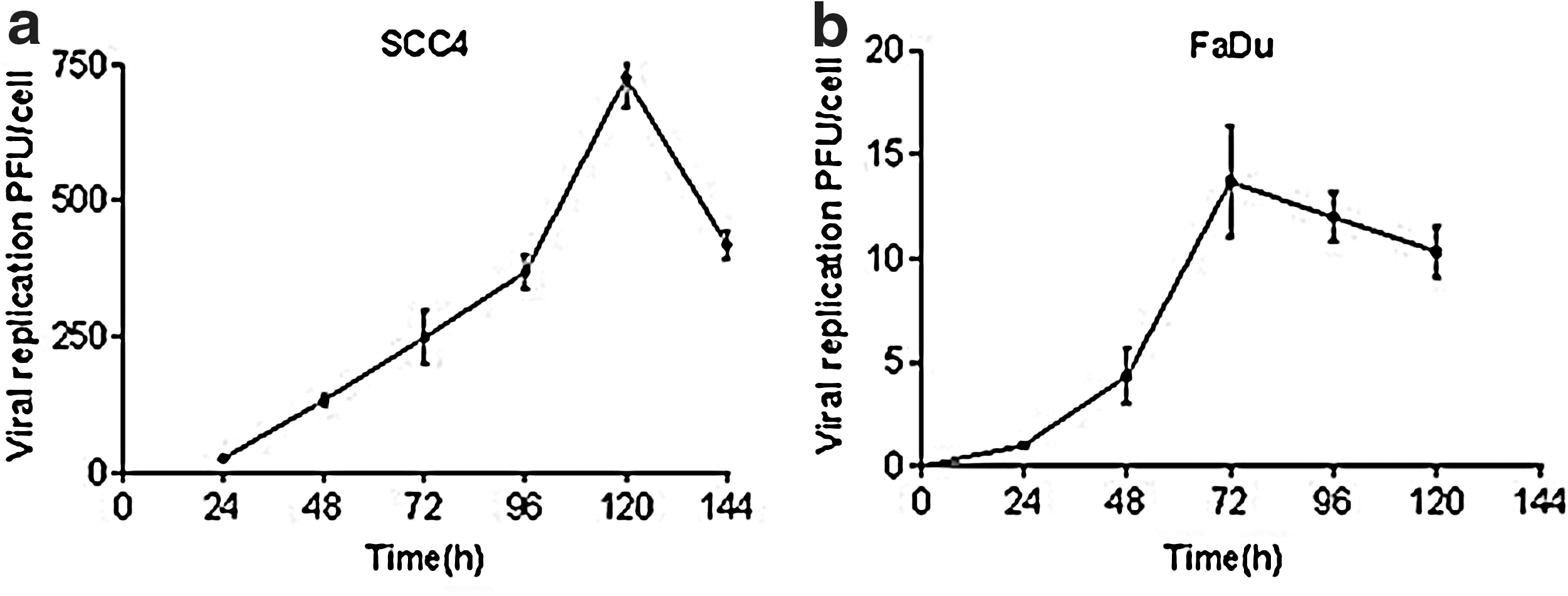
Replication of Lister vaccine strain of vaccinia virus (VVLister) in head and neck squamous cell carcinoma (HNSCC) cell lines. Cells were infected with 1 plaque-forming unit (PFU)/cell VVLister, and cell lysates were harvested at 24-hr intervals up to 144 hr. Mean viral replication ± SEM in human HNSCC cell lines.
EC50 values were derived by MTS assay 144 hr after infection with a serial of dose of viruses in various human HNSCC cell lines.
Vaccinia virus expressing human endostatin–angiostatin fusion protein
In order to enhance the efficacy of oncolytic vaccinia virus, the endostatin–angiostatin fusion gene was previously inserted at the Not1 restriction site of the VVLister (VVhEA) (Tysome et al., 2009a). Another recombinant virus (VVlacZ), constructed by insertion of lacZ at the same site, served as a control. The potency and replication of both recombinant viruses in vitro were consistently less than those of the parental VVLister (Fig. 2a, b). The potency of VVhEA was also less than that of VVlacZ in all cell lines tested. Peak replication of the recombinant viruses was lower than that of VVLister, although peak replication was significantly greater with VVlacZ than with VVhEA in SCC4 cells (Fig. 2b).
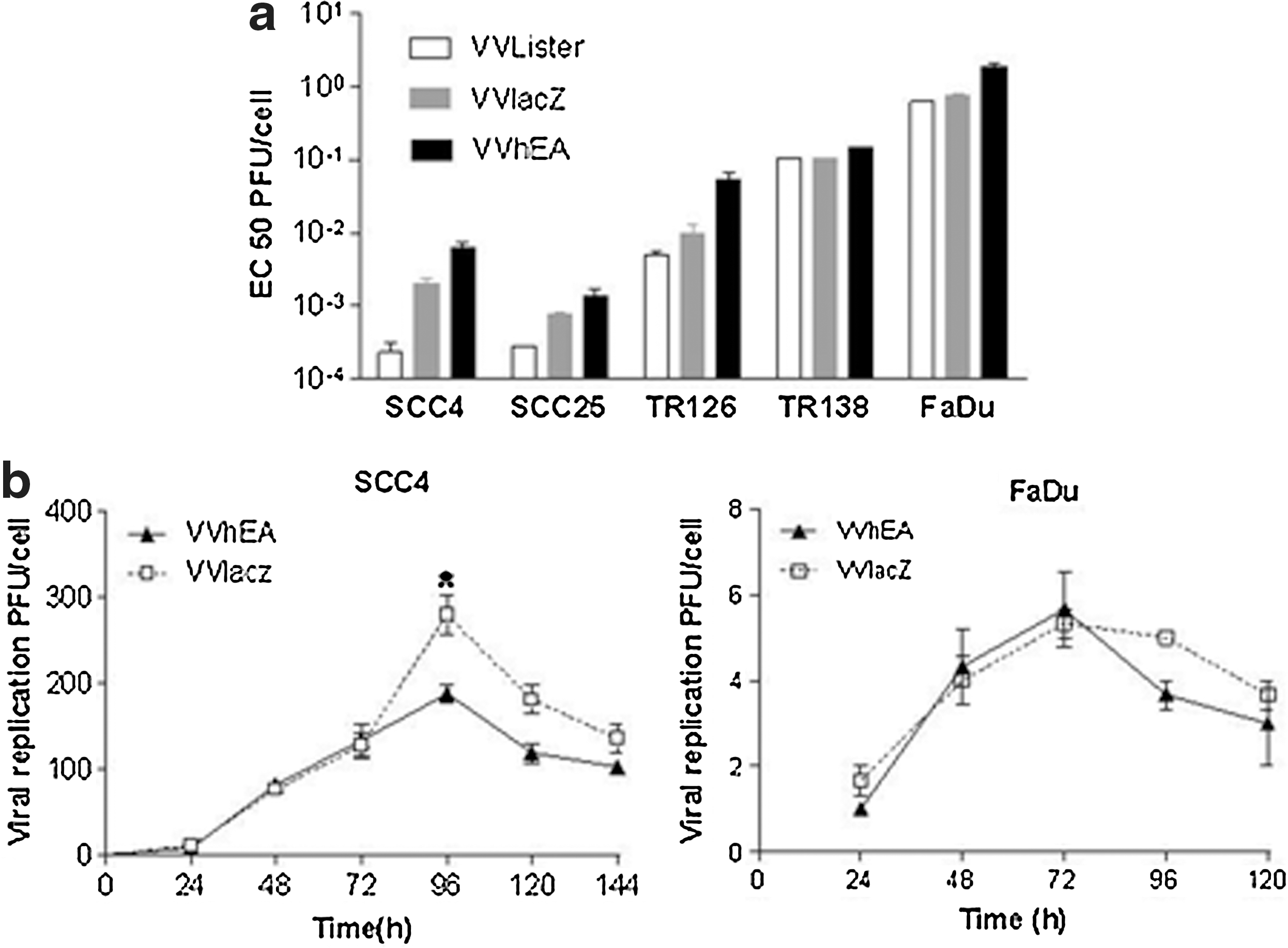
Potency and replication of recombinant vaccinia viruses.
Expression and function of the endostatin–angiostatin fusion protein in HNSCC in vitro
The expression of the human endostatin–angiostatin fusion gene in FaDu cells infected with VVhEA was confirmed by Western blotting (Fig. 3a). Although intracellular transgene expression was found in the cells by 24 hr following infection, endostatin–angiostatin fusion protein was detected in the supernatant only after 48 hr.
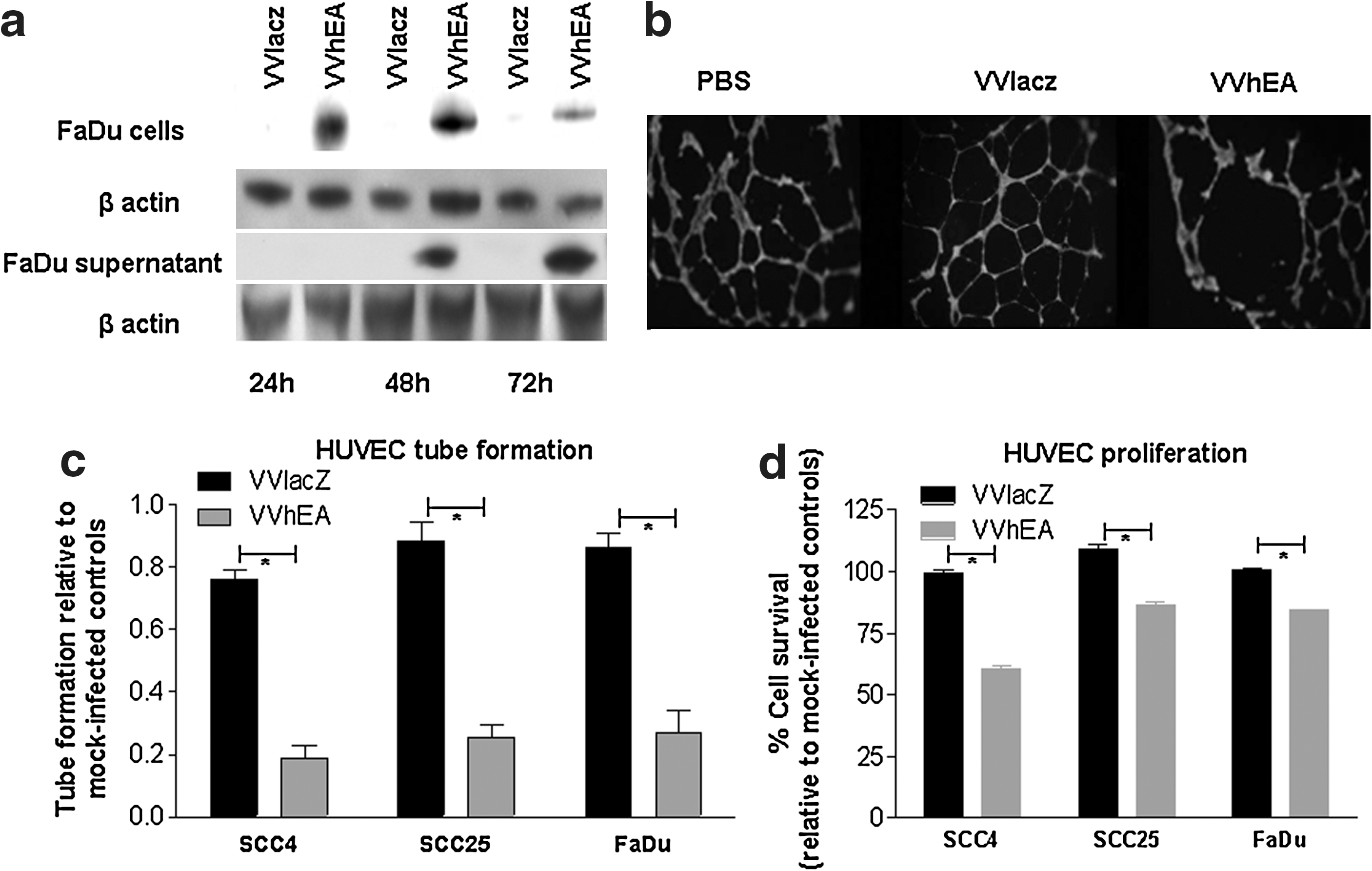
Expression and angiogenesis inhibition by the human endostatin-angiostatin fusion protein in HNSCC cells infected with VvhEA.
The antiangiogenic function of the endostatin–angiostatin fusion protein was confirmed by the inhibition of HUVEC tube formation and proliferation (Fig. 3b–d). HUVEC tube formation was significantly inhibited by supernatant from all HNSCC cells infected with VVhEA when compared with VVlacZ (p < 0.0001) or mock-infected controls (Fig. 3c). Although mean HUVEC tube formation following VVlacZ supernatant treatment was lower than that of the mock-infected controls, it did not reach significance. The inhibition of HUVEC proliferation was also significantly greater when treated with VVhEA supernatant than with VVlacZ (p < 0.001) or mock-infected controls (Fig. 3d).
Expression and function of the endostatin–angiostatin fusion protein in HNSCC in vivo
The expression of the human endostatin-angiostatin fusion gene in FaDu cells infected with VVhEA was confirmed by immunohistochemistry, observed from 3 to 17 days after one IT injection of 1 × 108 PFU VVhEA (Fig. 4a). Plasma levels of human endostatin continued to rise in the same animals up to 17 days after treatment (Fig. 4b). The antiangiogenic function of the endostatin–angiostatin fusion protein was confirmed in vivo by the significantly lower tumor microvessel density seen after the treatment of FaDu xenografts with VVhEA when compared with VVlacZ (Fig. 4c). However, even in VVhEA-treated tumors, microvessel density continued to rise up to 17 days after one treatment.
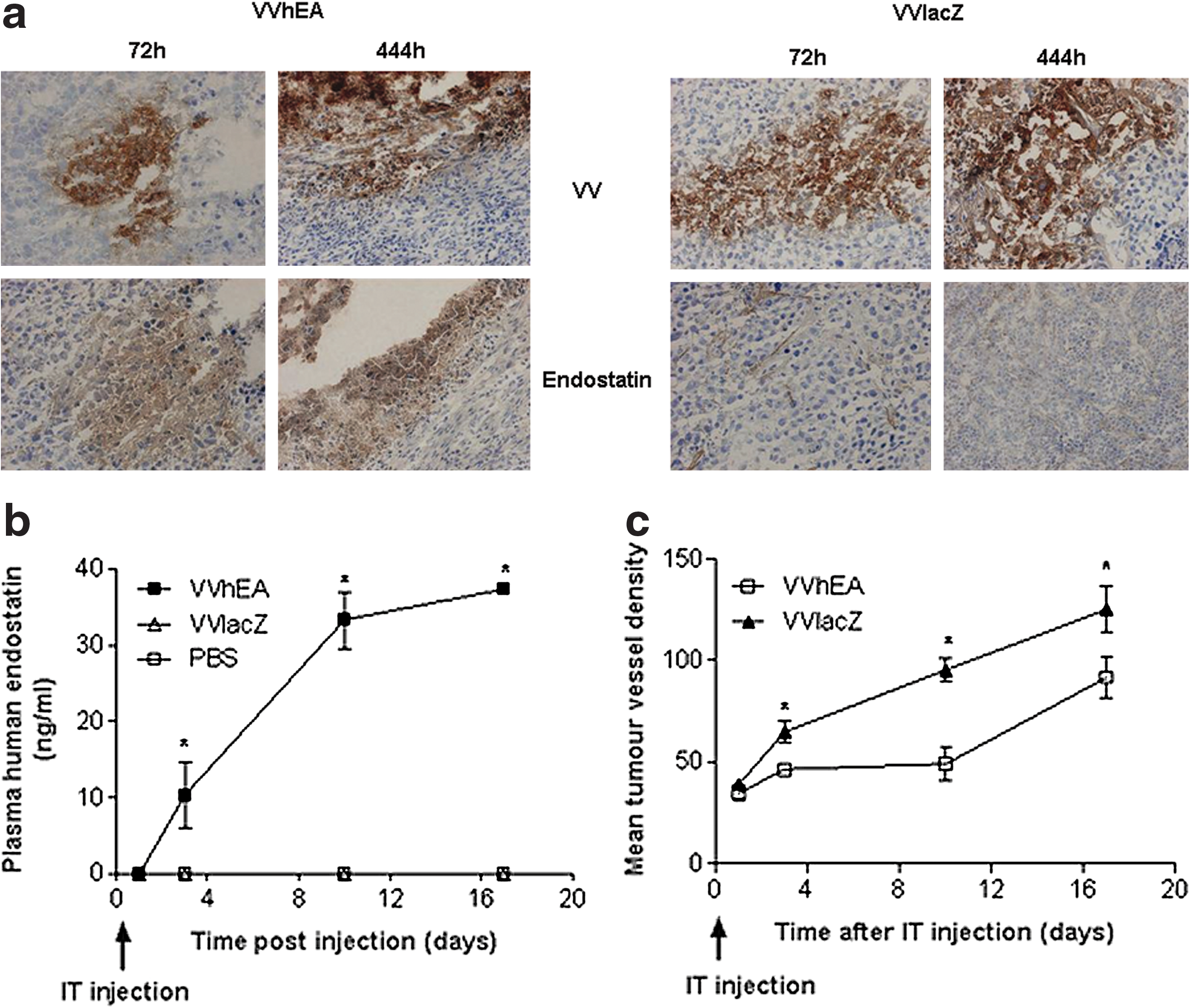
Biodistribution of vaccinia virus and human endostatin expression by immunohistochemistry and expression of human endostatin–angiostatin fusion protein in plasma from nude mice bearing FaDu xenografts treated with intratumoral (IT) vaccinia viruses. Subcutaneous FaDu tumors were grown in the right flank of 36 BALB/c nu/nu mice. When tumors reached 4–5 mm in diameter, 12 mice/group were injected with either IT 1 × 108 PFU VVhEA, VVlacZ, or PBS at time 0, 1, 3, 10, and 17 days later, and three mice per group were killed and tumors harvested. Immunohistochemistry for Lister vaccinia virus coat protein (VV), human endostatin (Endo), and CD31 was performed.
Treatment of a HNSCC model with oncolytic Lister vaccinia virus armed with the endostatin–angiostatin fusion gene improves survival significantly longer than dl1520
Given the poor efficacy of the first generation of oncolytic adenovirus dl1520 for HNSCC, we next sought to determine whether or not our VVhEA oncolytic virus is an alternative to dl1520 for treatment of HNSCC. To this end, the potency and replication of our novel vaccinia viruses were compared with dl1520 in vitro (Fig. 5a). The antitumor potencies of all vaccinia viruses were greater than dl1520, whereas replication after 72 hr was similar for both recombinant vaccinia viruses and dl1520 (Fig. 5b). In order to compare antitumor efficacy in vivo, the FaDu xenograft tumor model of human HNSCC was established in BALB/c nude mice and treated with IT injections of vaccinia viruses and dl1520 (Fig. 5c, d). Intravenous delivery was not used due to the rapid hepatic clearance of systemically delivered adenoviruses (Nunes et al., 1999).

Comparison of antitumor efficacy of VvhEA and dl1520 in human FuDu xenograft tumor model after IT treatment in nude mice.
The rate of initial tumor growth in vivo was rapid following treatment with all oncolytic viruses, with no significant differences when compared with PBS controls up until the death of the first animal (Fig. 5c). Treatment with both VVhEA and VVlacZ resulted in significantly longer survival of mice when compared with dl1520 (VVhEA p < 0.001; VVlacZ p < 0.0037) or PBS (VVhEA p < 0.0002; VVlacZ p < 0.0003). Animals treated with VVhEA also survived significantly longer than animals treated with VVlacZ (p < 0.015) (Fig. 5d). In addition, animals treated with VVhEA displayed no side effects, whereas pock lesions were observed from 20 days after initial treatment in the oral cavity or on the paws of mice treated with VVlacZ.
Discussion
We first demonstrated that VVLister was more effective than Ad5 at killing all HNSCC cells tested in vitro, supporting our hypothesis that vaccinia virus represents a good alternative to adenovirus for HNSCC gene therapy. Further experiments demonstrated that the Lister strain vaccinia virus armed with a human endostatin–angiostatin fusion gene (VVhEA) possesses superior antitumor potency compared with dl1520 against the FaDu model and is a potential therapeutic agent for HNSCC.
The Western Reserve (WR) strain of vaccinia virus has been used for the construction of most previous oncolytic vaccinia viruses, based on its superior lytic activity in vitro (Guo and Bartlett, 2004). However, it is a laboratory-based strain, without the advantage of clinical use as a vaccine, raising concerns regarding side effects if it were to be used in translational studies in humans. It is not clear whether the antitumor potency of the WR strain in vivo is better than that of the Lister strain. Both strains induce pock lesions in the paws, tail, and oral cavity of nude mice, independent of the route of delivery, although the side effects depend on the applied viral dose (Ober et al., 2002; Luker et al., 2005). However, the Lister strain, in contrast to the attenuated WR strain (McCart et al., 2001; Luker et al., 2005), has not been detected in the ovaries or brain after infection of mice (Tysome et al., 2009a). The presumed lower potency of the hyperattenuated Lister vaccine strain in vitro may be counterbalanced by a superior safety profile in vivo. The investigation of the oncolytic Lister strain of vaccinia virus enhanced by expression of the endostatin–angiostatin fusion gene was opportune, given the recent expansion in antiangiogenic therapies for HNSCC. We found that the potency and replication in HNSCC cells of both F14.5L-deleted viruses (VVhEA and VVlacZ) were attenuated when compared with the parental VVLister, consistent with our previous findings in pancreatic cancer models (Tysome et al., 2009a). Recombinant vaccinia virus therapy resulted in a significant improvement in survival when compared with dl1520-treated mice in vivo.
The dose of endostatin delivered is crucial, since recombinant endostatin has shown a biphasic U-shaped dose–response curve when administered to tumor-bearing mice in vivo (Celik et al., 2005). This was also observed in a phase I clinical trial in which recombinant endostatin was used to treat a range of metastatic solid tumors including HNSCC, melanoma, and breast cancer (Herbst et al., 2002; Davis et al., 2004). Even one intratumoral injection of 1 × 107 PFU VVhEA generated relatively high plasma endostatin levels. In addition to the therapeutic effect in terms of increased survival observed after treatment with VVhEA, the mice did not suffer from the pock formation observed in the oral cavities and tails of some mice after IT delivery of VVlacZ. We hypothesize that angiogenesis inhibition due to expression of the fusion protein may have prevented vaccinia-induced pock formation by limiting release of the virus into the circulation, although further investigation is clearly required.
We have demonstrated the potential of the Lister strain of vaccinia for treatment of HNSCC. The mechanisms controlling the tumor selectivity of oncolytic viruses appear to primarily involve tumor-specific inactivation of cell protection and immune responses. Although little is known about vaccinia virus–mediated oncolytic mechanisms, it is conceivable that mainly Toll-like receptor, protein kinase R, and interferon response pathways are involved, since vaccinia viruses produce inhibitors of these pathways to avoid the host immune response (Smith et al., 1997; Thorne et al., 2005; Vaha-Koskela et al., 2007). Through inactivation of the F14.5L gene product, we attenuated vaccinia virus, which resulted in a survival advantage over dl1520 in a human HNSCC model. In addition, by arming the Lister strain with the human endostatin–angiostatin fusion gene, we combined the antiangiogenic properties of the fusion protein with the oncolytic properties of the Lister virus, further increasing survival and decreasing virus-induced side effects. This is particularly encouraging in terms of potential for translational therapy, in which the minimization of side effects is paramount. This strain of vaccinia virus shows promise for development as an alternative oncolytic virus to adenovirus for the treatment of HNSCC in the future.
Footnotes
Acknowledgment
This project was supported by Cancer Research UK (C633-A6253/A6251 programme grant), Barts and The London Research Advisory Board, The Royal College of Surgeons of England (JT), and Nature Sciences Foundation of China (30530800).
Author Disclosure Statement
None of the authors have any financial arrangement or involvement with commercial organizations producing competing products.