
Abstract
Before the successful use of lentiviral vectors in clinical trials it is essential that strategies for direct vector delivery into the brain be evaluated in vivo, particularly as these vectors are significantly larger than the brain extracellular space. To date no such studies have been undertaken. In this study, convection-enhanced delivery (CED) was employed in an attempt to achieve widespread lentiviral delivery in the striatum. Infusions of equine infectious anemia virus (EIAV) and HIV vector constructs expressing the reporter gene β-galactosidase (β-Gal) were undertaken into the striatum at a range of flow rates and viral titers. In rats, all EIAV and HIV infusions led to the extensive transduction of cells in perivascular spaces throughout the brain. Although infusions were performed under standardized conditions, the number and volume of distribution of transduced cells were highly variable, with approximately one-third of EIAV infusions leading to no concentrated cell transduction in the striatum. Heparin coinfusion had no effect on EIAV distribution, although coinfusion of nimodipine resulted in a significant reduction in the number and volume of distribution of transduced cells. Intrastriatal EIAV delivery in pigs led to extensive transduction of mainly neurons, which could be effectively visualized in real time by T2-weighted magnetic resonance imaging. No infusions were associated with a significant inflammatory response. Therefore, despite its large size, lentiviral vectors can be administered by CED to the striatum in both small and large animal models. However, the variability in vector distribution under standardized conditions and widespread vector distribution through the perivascular spaces raise serious concerns regarding the practicality of lentivirus-mediated gene therapy in the brain in clinical practice.
Introduction
In contrast to other viruses such as herpes simplex virus (Marchand and Schwab, 1986) and adeno-associated virus (serotype 2; Passini et al., 2005), lentiviral vectors, unless pseudotyped with rabies virus glycoprotein, are not retrogradely transported along axons (Mazarakis et al., 2001). As a consequence lentiviruses are ideal for focal transgene delivery to discrete neuroanatomical structures, particularly if localized transgene expression is required to minimize potential side effects. Indeed, an EIAV vector is currently being employed in a phase 1 clinical trial to deliver genes for dopamine synthesis into the striatum of patients with Parkinson's disease. However, techniques for the safe and effective administration of lentiviral vectors to the brain have yet to be systematically examined in animal models.
Convection-enhanced delivery (CED) is the most rational available technique for the direct intracranial administration of viral vectors into the brain in clinical practice. This technique necessitates the use of an appropriate cannula and infusion flow rate to distribute therapeutic agents by bulk flow directly into the brain extracellular space. In contrast to direct intracranial injection, it is possible to achieve widespread and homogeneous vector distribution by CED, and consequently this technique has been employed extensively for the administration of adeno-associated viral (AAV) vectors to the brain in animal studies and clinical trials (Cunningham et al., 2000, 2008; Nguyen et al., 2001; Chen et al., 2005; Sanftner et al., 2005; Hadaczek et al., 2006, 2009; Kaplitt et al., 2007; Szerlip et al., 2007; Eberling et al., 2008; Fiandaca et al., 2009). However, lentiviruses are significantly larger than AAV vectors, with a diameter between 80 and 120 nm (Segura et al., 2006), and the average diameter of the brain extracellular space has been measured at only 38 to 64 nm (Thorne and Nicholson, 2006). In view of the discrepancy in size between the brain extracellular space and the diameter of lentiviral particles, there are clearly significant doubts regarding the ability of lentiviral vectors to be administered effectively by CED. Indeed, to date no attempt has been made to evaluate the feasibility of lentiviral vector administration by CED.
In this study the feasibility of administering lentiviral vectors by CED is examined objectively under highly standardized conditions, which we previously employed to evaluate the distribution properties of vectors derived from a range of serotypes of AAV (White et al., 2011). Specifically, the relationships between infusion flow rate, viral titer, cell tropism, and volume of distribution of cell transduction are examined in the striatum of rats and pigs with a view to determining whether this vector could be effectively administered by CED in clinical practice.
Materials and Methods
Vectors
Vesicular stomatitis virus glycoprotein (VSV-G)-pseudotyped EIAV vectors expressing a β-galactosidase reporter gene under the control of a cytomegalovirus (CMV) reporter gene were kindly provided by Oxford BioMedica (Oxford, UK). VSV-G-pseudotyped HIV vectors expressing a β-galactosidase reporter gene under the control of a CMV reporter gene were procured from Cambridge Bioscience (Cambridge, UK).
Vector infusions
All procedures were carried out in accordance with U.K. Home Office animal welfare regulations and with appropriate Home Office licenses.
Rat infusion apparatus and procedures
Rat infusions were performed by the same technique employed in an earlier study evaluating the distribution properties of vectors derived from various serotypes of AAV (White et al., 2011). Infusions were performed and analyzed on an intention-to-treat basis. Acute infusion cannulas were constructed from lengths of fused silica with an outer diameter of 220 μm and an inner diameter of 150 μm. These lengths of fused silica were connected to 10-μl Hamilton syringes via a connection device made in-house that served to create a seal between the chamber of the Hamilton syringe and the lumen of the fused silica, and which guided the fused silica cannula from the Hamilton syringe, through the dura and into the brain. The Hamilton syringe, with the cannula attached via this connector device, was then mounted in an infusion pump (World Precision Instruments, Sarasota, FL) attached to a stereotactic frame (Stoelting, Wood Dale, IL) in which rats were immobilized. To insert a cannula, the entire pump/syringe/connector/cannula construct was lowered in the stereotactic frame until the target depth was reached.
Male Wistar rats (B&K Universal, Hull, UK) were group-housed and allowed to acclimatize before experimental procedures. Male rats weighed 225 to 275 g were anesthetized with an intraperitoneal dose of ketamine and xylazine and placed in a stereotactic frame (Stoelting). A linear incision was made between the glabella and the occiput and the skull exposed. Burr holes with a diameter of approximately 2 mm were placed 0.5 mm anterior and 2.75 mm lateral to the bregma and cannulas were inserted to a depth of 5 mm below the dura. All cannulas were preprimed with vector before insertion into the brain. Every attempt was made to ensure that no air bubbles were present in the infusion cannula. Infusions of 4 μl of vector were performed at flow rates of 0.1, 1, 2.5, and 5 μl/min bilaterally into the striatum of rats. EIAV infusions were undertaken at concentrations of 2.8×1010 transducing units (TU)/ml and 5×107 TU/ml. HIV infusions were undertaken at a concentration of 1.5×105 TU/ml. A number of infusions were also performed with coinfused heparin and nimodipine (Bayer HealthCare Pharmaceuticals, Wayne, NJ). Summaries of the infusions performed are outlined in Figs. 1 and 3.
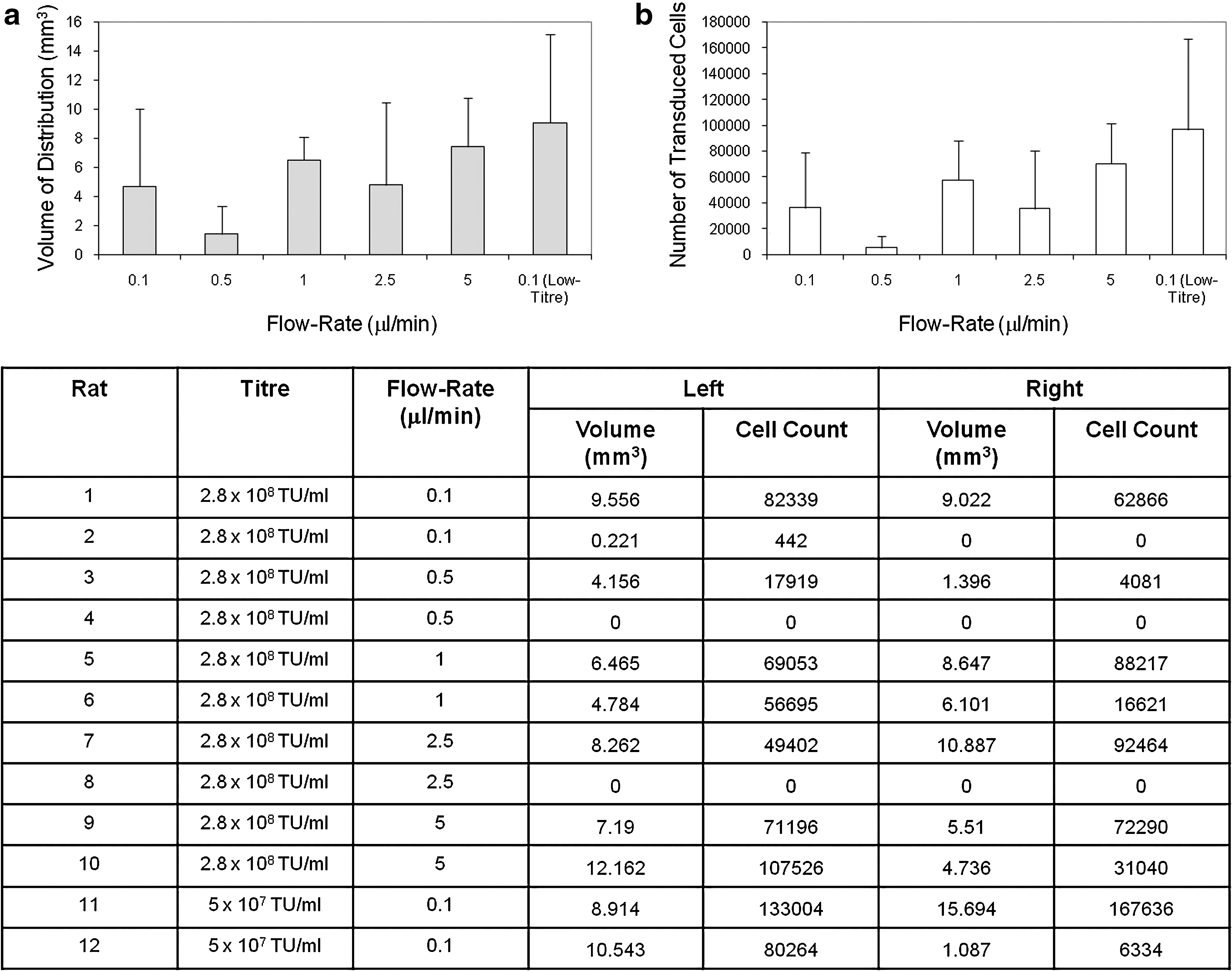
Equine infectious anemia virus (EIAV) distribution properties. Evaluation of the relationship between viral titer, infusion flow rate, and
Animals were killed by transcardial perfusion fixation 21 days after vector administration, and the brains were harvested for histological analysis. After infusion completion, the cannula was left in situ for 5 min before being withdrawn at a rate of 1 mm/min. The wound was then closed with 4/0 Vicryl, a dose of intramuscular buprenorphine was administered (30 μg/kg), and the anesthetic was reversed with a 0.1-mg/kg intraperitoneal dose of atipamezole hydrochloride (Antisedan, 200 μg/kg; Pfizer, Kent, UK). After 28 days, animals were perfusion fixed with 100 ml of phosphate-buffered saline (PBS) followed by 100 ml of 4% paraformaldehyde (PFA, pH 7.4). The brain was then removed from the skull and placed in 4% PFA (pH 7.4) for 48 hr and then cryoprotected in 30% sucrose before sectioning.
Pig infusion apparatus and procedures
The pig was administered an intramuscular dose of ketamine (0.1 mg/kg body weight). General anesthesia was then induced and maintained with isoflurane (2–5%) and the animal was intubated with a cuffed endotracheal tube. Intravenous access was obtained with a cannula placed in an ear vein and normal saline was infused at a rate of 250 ml/hr.
Pig head fixation was achieved with a custom-built fixation device incorporating bilateral magnetic resonance imaging (MRI)-compatible zygomatic screws, a moldable palate tray, and snout strap. All materials were fully MR compatible to prevent imaging artifact. After robust pig head fixation, an arc of fiducials was placed over the animal's head. Flex-L coils were then attached to the lateral aspects of the head and the animal was transferred to a 1.5T MRI scanner (Intera; Philips, Best, The Netherlands). Stereotactic surgical planning and procedures were undertaken with a PathFinder (Prosurgics, High Wycombe, UK) stereotactic robotic arm and associated software. Briefly, this stereotactic arm functioned as follows. The pig was imaged with an array of fiducial balls placed in a fixed location over the animal's head. In theater, the fiducial balls were replaced with optical reflector balls placed into precisely the same locations. The locations of the reflector balls were visualized with a camera in the underside of the robotic arm. The optical reflector balls and MRI fiducials were coregistered automatically by the planning software. The software allowed visualization of MR images only from a single plane. As a consequence, as coronal images facilitated the best views of the planned cannula trajectory, they were used for all surgical planning. To conduct the surgical procedure, a range of end-effectors designed to accommodate the burr hole generation and cannula delivery tooling were placed onto the robotic arm.
Vector infusions were undertaken in a male Large White/Landrace pig weighing 40 kg, using a catheter delivery system developed in-house. Specifically, infusions of 120 μl containing 2.8×108 TU/ml were undertaken bilaterally into the putamen, using the following ramped infusion regimen: 0.5 μl/min for 5 min, 1 μl/min for 5 min, 2.5 μl/min for 5 min, and then 5 μl/min for 20 min. After infusion completion, the catheter was left in place for 10 min before being withdrawn slowly by hand. The burr holes were then filled with bone dust and the wound was then closed with interrupted 4/0 Prolene sutures (Ethicon, Somerville, NJ). The animal was then transferred back to the MRI scanner and T2-weighted imaging was immediately performed at a slice thickness of 2 mm. As a consequence imaging was undertaken approximately 30 min after infusion cessation. The animal was allowed to recover for a period of 28 days. The animal was then killed by perfusion fixation under terminal anesthesia and the brain was harvested for histological analysis.
Histology
Rat brains were cut into 35-μm-thick coronal sections with a Leica CM1850 cryostat (Leica Microsystems, Wetzlar, Germany). Pig brains were cut into 100-μm coronal sections with a Leica SM2500 microtome.
5-Bromo-4-chloro-3-indolyl-β-
Immunohistochemistry was performed on sections adjacent to X-Gal-stained sections. Briefly, all solutions for immunohistohemistry were made in PBS. Free-floating PFA-fixed sections were washed three times for 15 min in PBS and incubated in 3% hydrogen peroxide to remove endogenous peroxidase activity. Sections were then washed three times for 15 min in PBS, before being blocked for 1 hr in blocking solution (10% normal goat or donkey serum) at room temperature. Sections were then transferred directly from blocking solution into primary antibody, appropriately diluted in blocking solution, and incubated overnight. The following primary antibodies were used: mouse anti-neuronal nuclei (NeuN) (diluted 1:300; Chemicon, Chanders Ford, Hampshire, UK), rabbit anti-glial fibrillary acidic protein (GFAP) (diluted 1:200; Chemicon), mouse anti-ED1 (diluted 1:100; Serotec, Oxford, UK), mouse anti-CD4 (OX38) (diluted 1:300; Serotec), mouse anti-CD8 (OX8) (diluted 1:300; Serotec), mouse anti-desmin (diluted 1:200; Abcam, Cambridge, UK), and rabbit anti-β-Gal (diluted 1:200; Chemicon). After three PBS washes, sections were then incubated with secondary antibody for at least 2 hr at room temperature. For fluorescence immunohistochemistry, species-specific secondary antibodies (Cy3) were used (diluted 1:200; Jackson Immunoresearch Laboratories, West Grove, PA). After PBS washes, sections were mounted in Vectashield (Vector Laboratories) on gelatin-coated slides and coverslipped, before fluorescence imaging.
Imaging
Imaging was undertaken with a Leica DM5500 microscope (Leica Microsystems) and digital camera (MicroBrightField, Williston, VT). Stereological counts were undertaken on immunostained sections with commercially available software (Stereo Investigator; MicroBrightField). Briefly, population estimates were undertaken on representative tissue sections to determine the counting frame size, counting frame number, and number and separation of tissue sections necessary to achieve an accurate cell count with a Gundersen (m=1) coefficient of error less than 0.1. Using these parameters, cell counts were then undertaken on serial sections a uniform distance apart, using an optical fractionator probe. The volume of distribution of transduced cells was calculated by tracing contours around the outer margins of the enhanced green fluorescent protein (EGFP)-expressing cells on each section. Transduced cells outside the striatum were excluded from these contours to ensure that only the intrastriatal volume of distribution of transduced cells was calculated. Infusions that were associated with obvious leakage of vector into the ventricular system were excluded from further analysis. Determination of the vector cell tropism and the density of activated microglia and CD4- and CD8-positive T lymphocytes in the volume of viral distribution for each infusion was performed on selected tissue sections close to the needle track, using the fractionator probe.
Coronal MR images were correlated with histological sections by measuring the distance of both MR images and histological sections from the posterior commissure. Although MR images had a thickness of 2 mm, they were correlated with histological sections in the middle of this volume.
In vitro assay of neuronal transduction
To assess whether addition of the calcium antagonist nimodipine (Bayer HealthCare Pharmaceuticals) had an effect on EIAV-mediated cell transduction, primary neuronal cultures were generated from fetal rat hippocampi (Brewer et al., 1993). After 5 days in culture, cells were transduced with EIAV vectors expressing β-Gal in the presence or absence of nimodipine at a concentration of 2.5×10–8 M. Cells were then maintained for a period of 7 days before being fixed with 2% PFA. β-Galactosidase expression was assessed by performing immunohistochemistry. After overnight incubation in mouse anti-β-galactosidase primary antibody (diluted 1:300; Chemicon), cells were washed in PBS and then incubated for 1 hr in rabbit anti-mouse secondary antibody (Cy2; Jackson Immunoresearch Laboratories). Cell transduction was determined by fluorescence microscopy. In vitro assays were repeated 10 times and the mean value for the percentage of neurons transduced from these assays was calculated.
Statistical analysis
Kruskal–Wallis one-way analysis of variance was performed to determine whether there was a significant difference in vector distribution and cell transduction associated with various infusion parameters.
Results
Convection-enhanced delivery of EIAV into the striatum of rats
Highly variable transduction and distribution were observed when EIAV was infused into the striatum of rats. Accurate cannula implantation into the striatum was observed in all cases as determined by a visible cannula track terminating in the striatum. There was no evidence of infusate reflux with any infusion. There was no statistically significant relationship between the infusion flow rate or viral titer infused and the resulting number or volume of distribution of transduced cells. There was also significant variability in both the number and volume of distribution of transduced cells between infusions performed at the same flow rate and viral titer (Fig. 1a and b). Some infusions were associated with no concentrated transduction of cells within the striatum. There was no obvious relationship between this finding and the flow rate or viral titer employed.
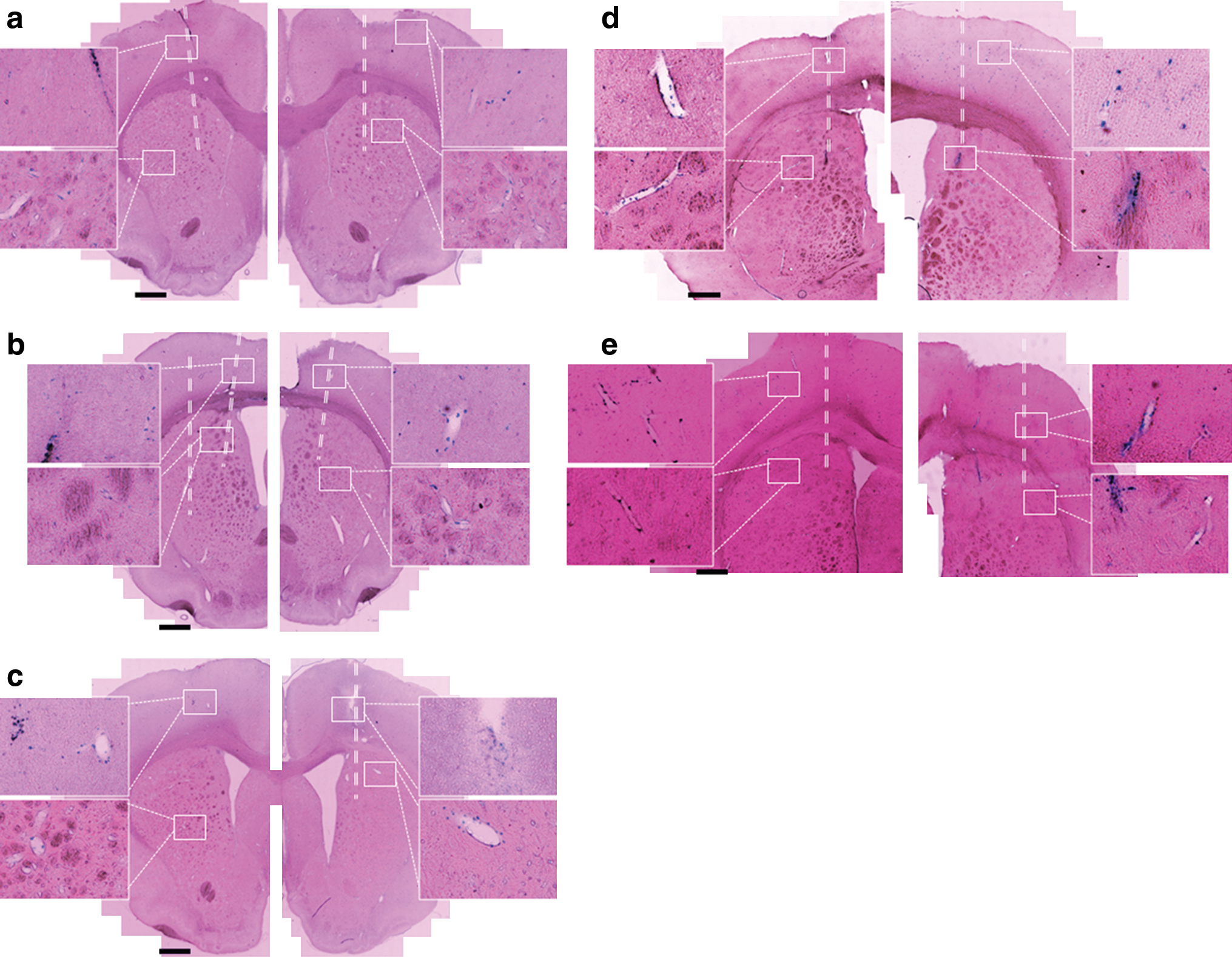
Detailed examination of X-Gal-stained tissue sections demonstrated two patterns of cell transduction (Fig. 2). The majority of infusions (19 of 24) led to concentrated cell transduction within the striatum (Fig. 2a). The remaining infusions (5 of 24) did not result in concentrated cell transduction in the striatum, although closer examination demonstrated widespread cell transduction throughout the brain. In these cases cell transduction appeared to be located in the perivascular spaces. For these infusions, the cannula track could be identified histologically and the cannula tip appeared to have been inserted immediately adjacent to a large vessel (Fig. 2b–d).

Patterns of EIAV distribution. Representative X-Gal-stained tissue sections show the two principal patterns of distribution of transduced cells in rat brain. The majority of infusions (19 of 24) led to concentrated cell transduction in the striatum
Convection-enhanced delivery of HIV into the striatum of rats
To determine whether the widespread perivascular distribution of EIAV is unique amongst lentiviral vectors, HIV infusions were undertaken into the striatum of rats. Infusions of 1.0×105 TU/ml were performed at infusion rates of 0.1, 0.5, 1.0, 2.5, and 5.0 μl/min. All infusions led to minimal concentrated cell transduction in the striatum, but as with EIAV administration, widespread cell transduction was observed in perivascular spaces throughout the brain (Fig. 3).
HIV distribution properties. Coronal tissue sections obtained along the cannula track demonstrate the distribution of cell transduction after infusions of (1×105) of HIV into the striatum of rats at flow rates of 0.1
Effect of heparin and nimodipine coinfusion on EIAV distribution
In an attempt to improve the distribution of EIAV and to minimize variability, infusions were repeated at a low titer (5×107 TU/ml) with heparin to minimize nonspecific viral binding or with nimodipine to dilate the cerebral vasculature and minimize vector flow through the perivascular spaces. Of four infusions with heparin, only one infusion led to concentrated cell transduction in the striatum. Interestingly, that infusion led to the greatest volume of distribution of transduced cells seen with any infusion. The remaining three infusions led to cell transduction throughout the brain, although cell transduction was confined to perivascular spaces. Overall, there was no statistically significant difference in the volume or number of transduced cells with heparin coinfusion compared with control infusions (p=0.18 and 0.14, respectively). Coinfusion with nimodipine led to small volumes of cell transduction within the striatum (Fig. 4), although nimodipine had no effect on the ability of EIAV to transduce neurons in a primary neuronal rat hippocampal culture. Specifically, application of EIAV in a solution of nimodipine resulted in the transduction of 78% (±2%) of neurons compared with EIAV instillation with vehicle, which resulted in the transduction of 79% (±6%) of neurons in vitro. Overall, the reduction in the number and volume of transduced cells with nimodipine coinfusion compared with control infusions was statistically significant (p=0.005 and 0.007, respectively). With all of these infusions, cell transduction was also visualized within perivascular spaces throughout the brain.
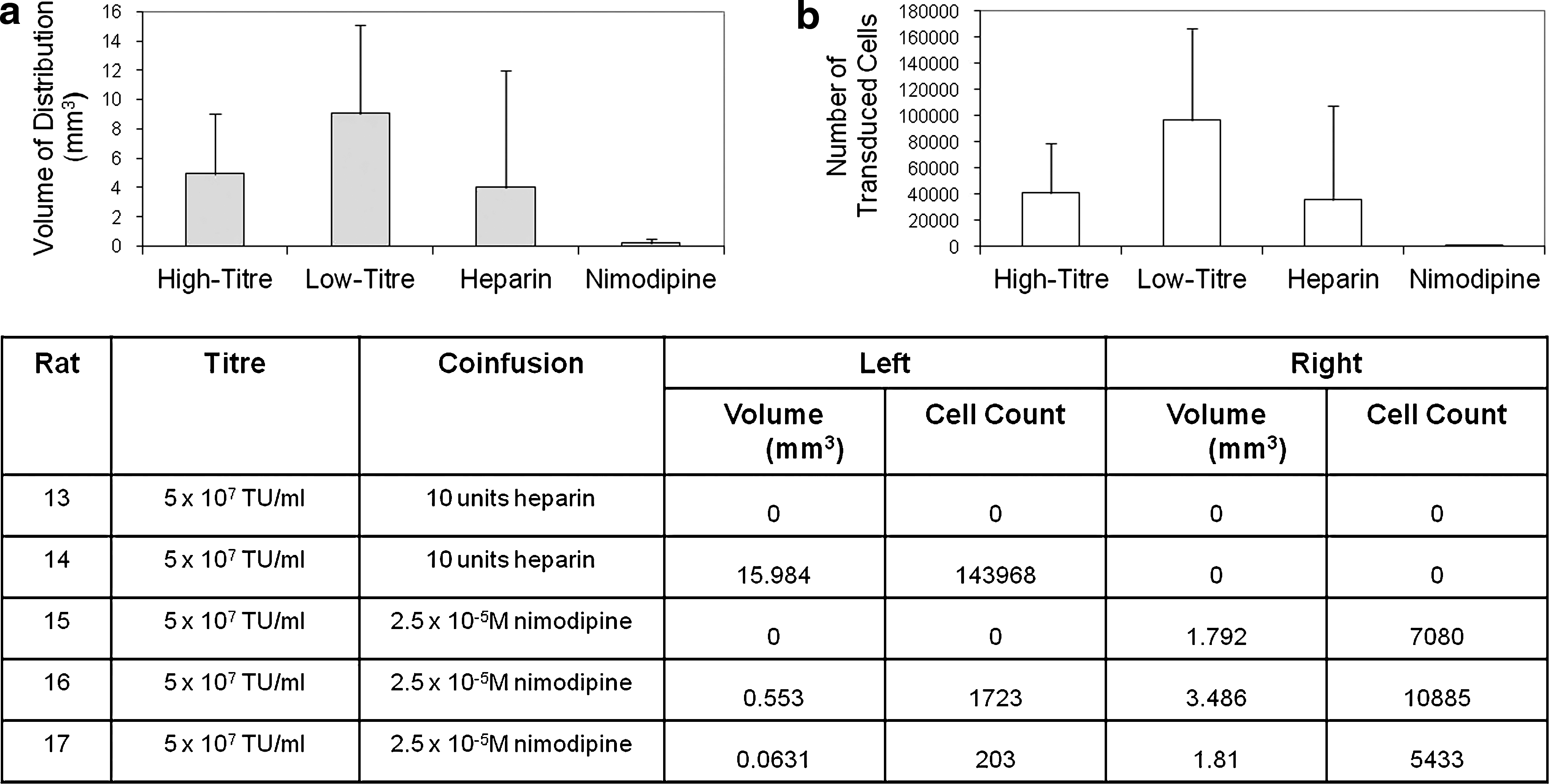
Effect of heparin/nimodipine coinfusion on EIAV distribution. Shown is the evaluation of the effects of heparin and nimodipine coinfusion on the volume of distribution of transduced cells
Transductional tropism of EIAV in rat brain
Double immunostaining of tissue sections for β-galactosidase and the neuronal marker NeuN demonstrated that the majority of transduced cells in areas of concentrated cell transduction were neuronal (all infusions, 85.8±4.7%; Fig. 5a–c). There was no significant difference in the proportion of transduced cells that were neuronal at different viral titers (2.8×108 TU/ml, 85.1±5.6%; 5×107 TU/ml, 85.8±2.9%) or with coinfusion of nimodipine (86.9±4.5%) or heparin (89.3%; only a single infusion led to concentrated cell transduction).

EIAV tropism in rat striatum. Representative image shows extensive β-galactosidase expression in the striatum
Immune cell infiltration after CED of EIAV into the rat striatum
Double immunostaining for β-galactosidase expression and the presence of the inflammatory cell markers ED1 (activated microglia), CD4 (helper T cells), and CD8 (cytotoxic T cells) demonstrated minimal tissue infiltration of inflammatory cells except along the course of the needle track, where there was a focal infiltration of both ED1-positive activated microglia and CD8-positive T cells. Negligible inflammatory cell infiltration into the brain parenchyma was observed regardless of the infused viral titer, infusion flow rate, use of coinfusates, and the presence of concentrated cell transduction in the striatum or diffuse cell transduction in the perivascular spaces throughout the brain (Fig. 6).
Inflammatory cell infiltration into EIAV-transduced striatum. Representative images show ED1-positive microglia
Confocal microscopic examination of cell transduction
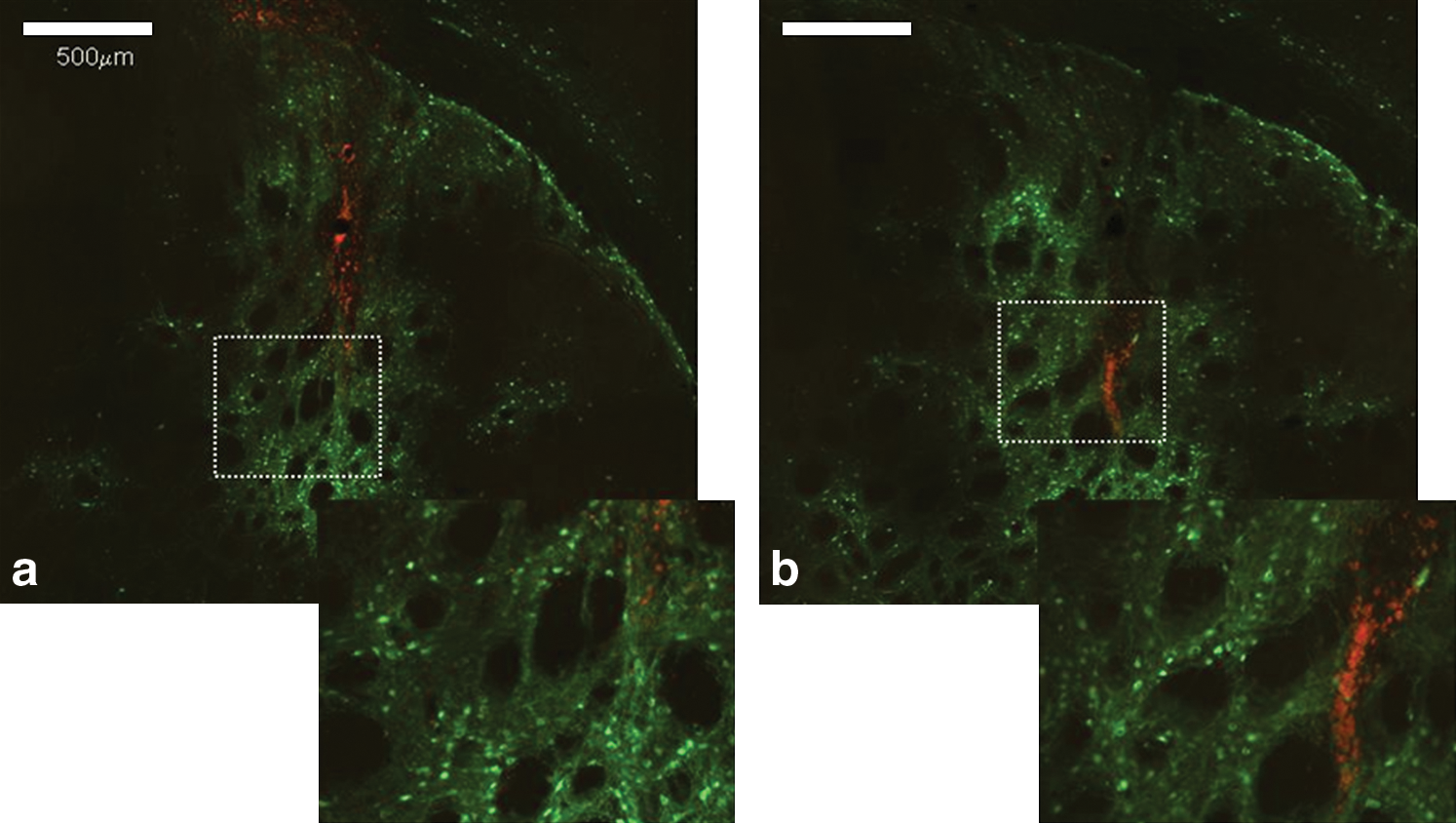
In view of the extensive transduction of cells in perivascular spaces throughout the brain after infusions of EIAV into the rat striatum, it was hypothesized that if vector distribution occurs primarily through the perivascular spaces, in areas of concentrated cell transduction these transduced cells should lie in immediate proximity to vessels. Consequently, the microstructural location of transduced cells within concentrated areas of cell transduction in the rat striatum was examined by confocal microscopy. Examination of representative sections demonstrated that the majority of transduced cell bodies were not in immediate proximity to obvious vessels suggesting EIAV vector penetration into the neuropil (Fig. 7).
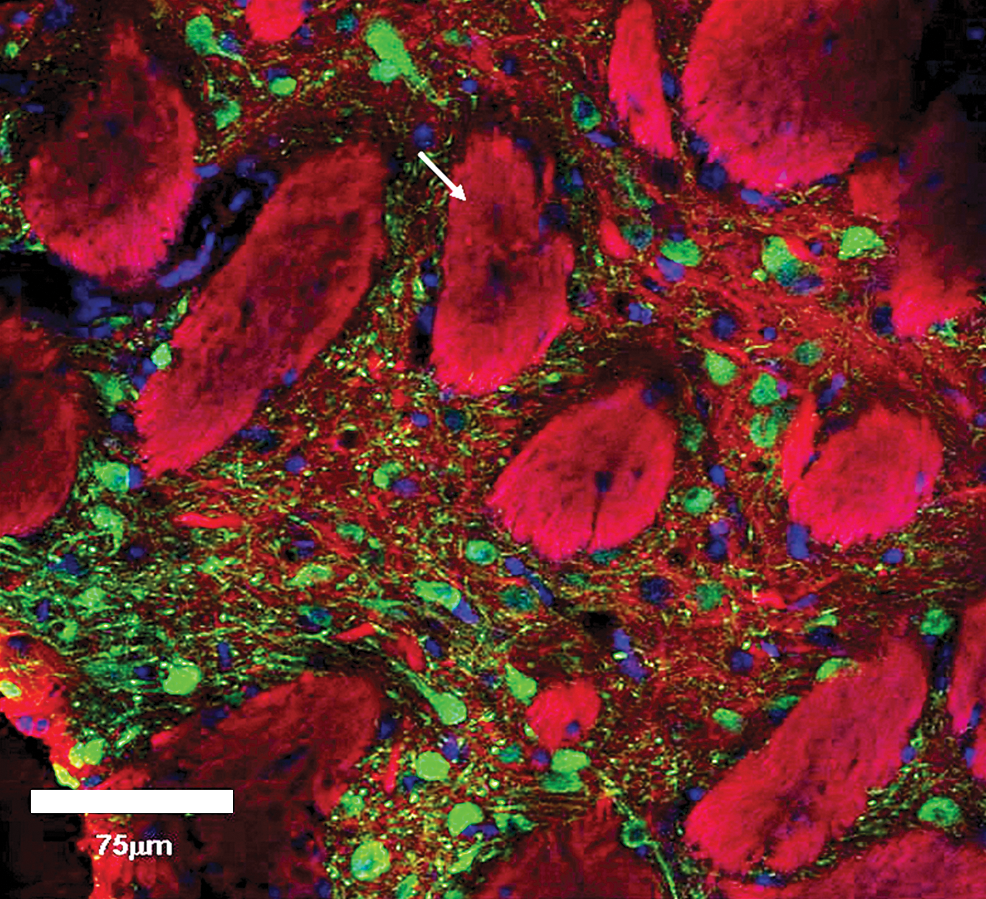
Confocal microscopy examination of EIAV-transduced striatum. Shown is a confocal image of concentrated β-galactosidase expression (green) in the striatum, DAPI staining of cell nuclei (blue), and Dil staining of lipid membranes (red). The large Dil-stained structures are striosomes (arrowed). The majority of transduced cells are not clearly located in immediate proximity to perivascular spaces, suggesting vector distribution into the neuropil. Color images available online at
Convection-enhanced delivery of EIAV into the striatum of a pig
Infusions of 120 μl of EIAV vector were performed bilaterally into the striata of a pig brain. In the left hemisphere, highly concentrated X-Gal staining was present in a pattern that closely resembled areas of high signal on T2-weighted MR images obtained immediately after vector infusion (Fig. 8). In the right hemisphere, immediate postoperative MR imaging revealed an area of high signal in the striatum surrounding a small area of low signal. This finding was compatible with a small intraparenchymal hemorrhage surrounded by infusion-related edema. As in the left hemisphere, histologically there was widespread X-Gal staining in the striatum that accurately correlated with areas of high signal on T2-weighted MR images. In addition, histology confirmed the presence of a small intraparenchymal hemorrhage at the cannula tip, although the animal had no clinically detectable neurological deficit postoperatively (Fig. 9).
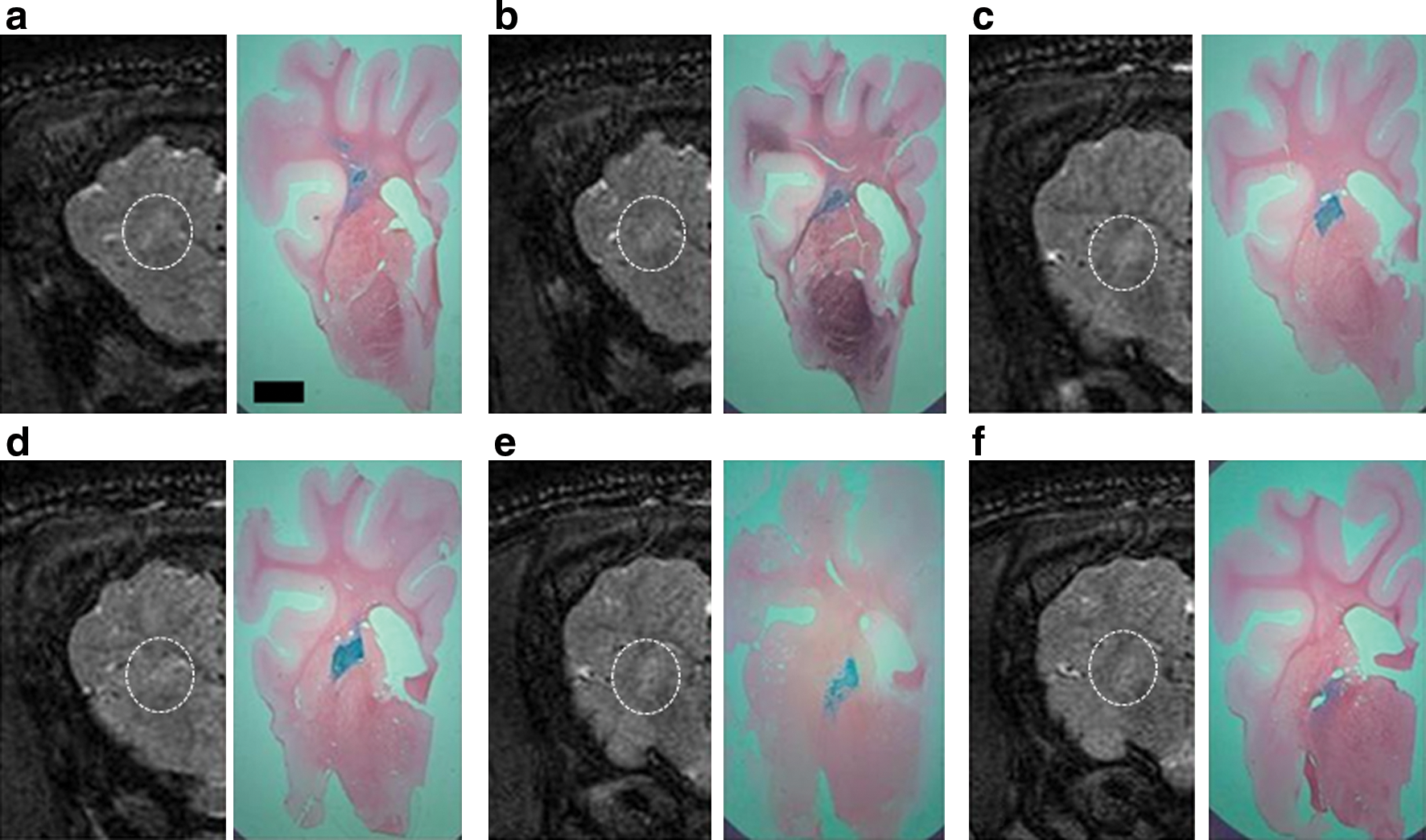
Right pig hemisphere.
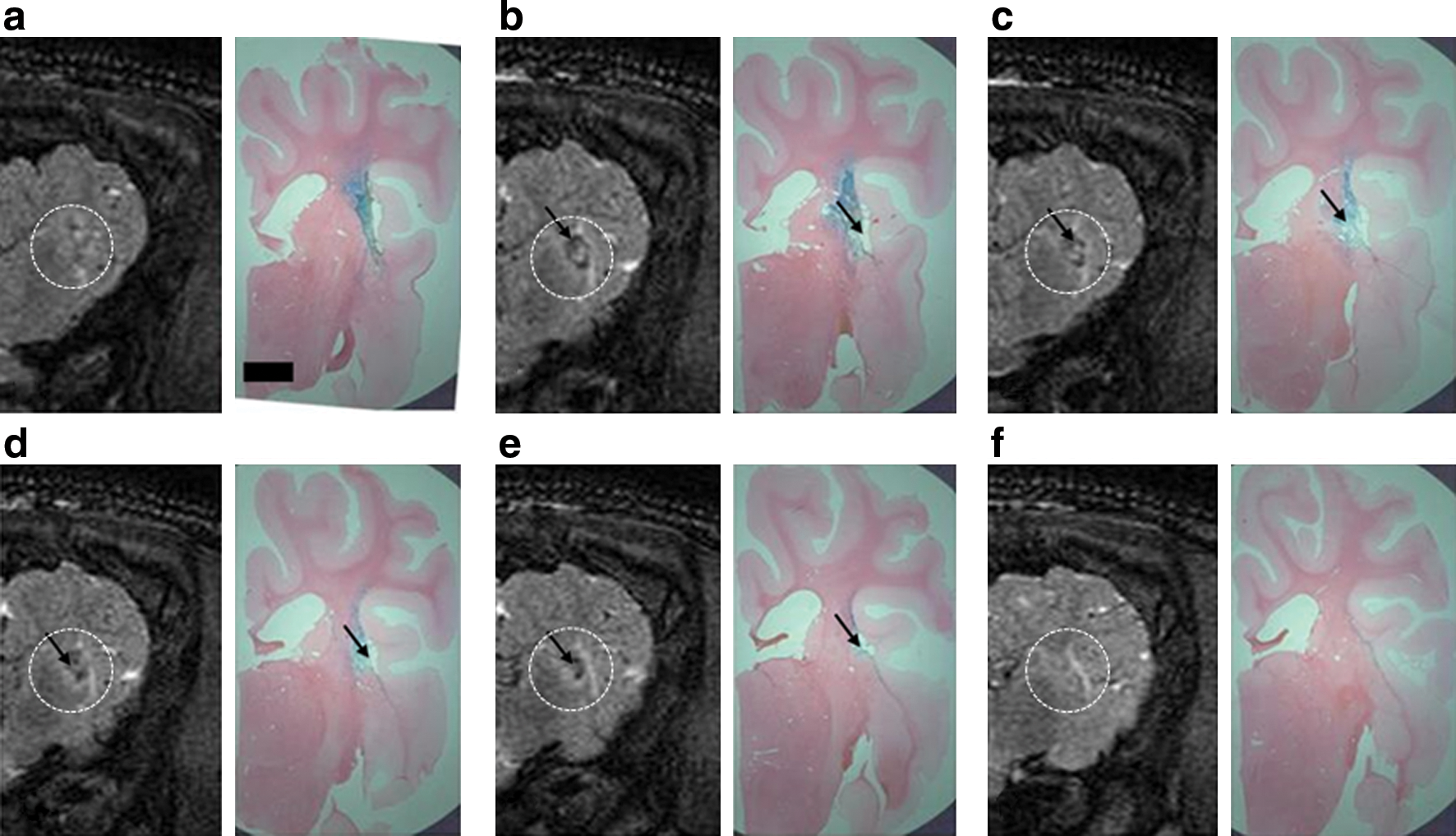
Left pig hemisphere.
In both hemispheres, there was evidence of extensive X-Gal staining in perivascular spaces. This was most evident in vessels adjacent to the infusion site, although cell transduction in perivascular spaces was also evident up to 4 cm away. The majority of cell transduction in perivascular spaces distant from the infusion site was found in the distribution of the lenticulostriate arteries, although perivascular spaces in the frontal lobe cortex and thalamus were also affected (Fig. 10).
Perivascular transport of EIAV. Representative X-Gal-stained histological sections show extensive X-Gal staining in the striatum
Transductional tropism of EIAV in the pig brain
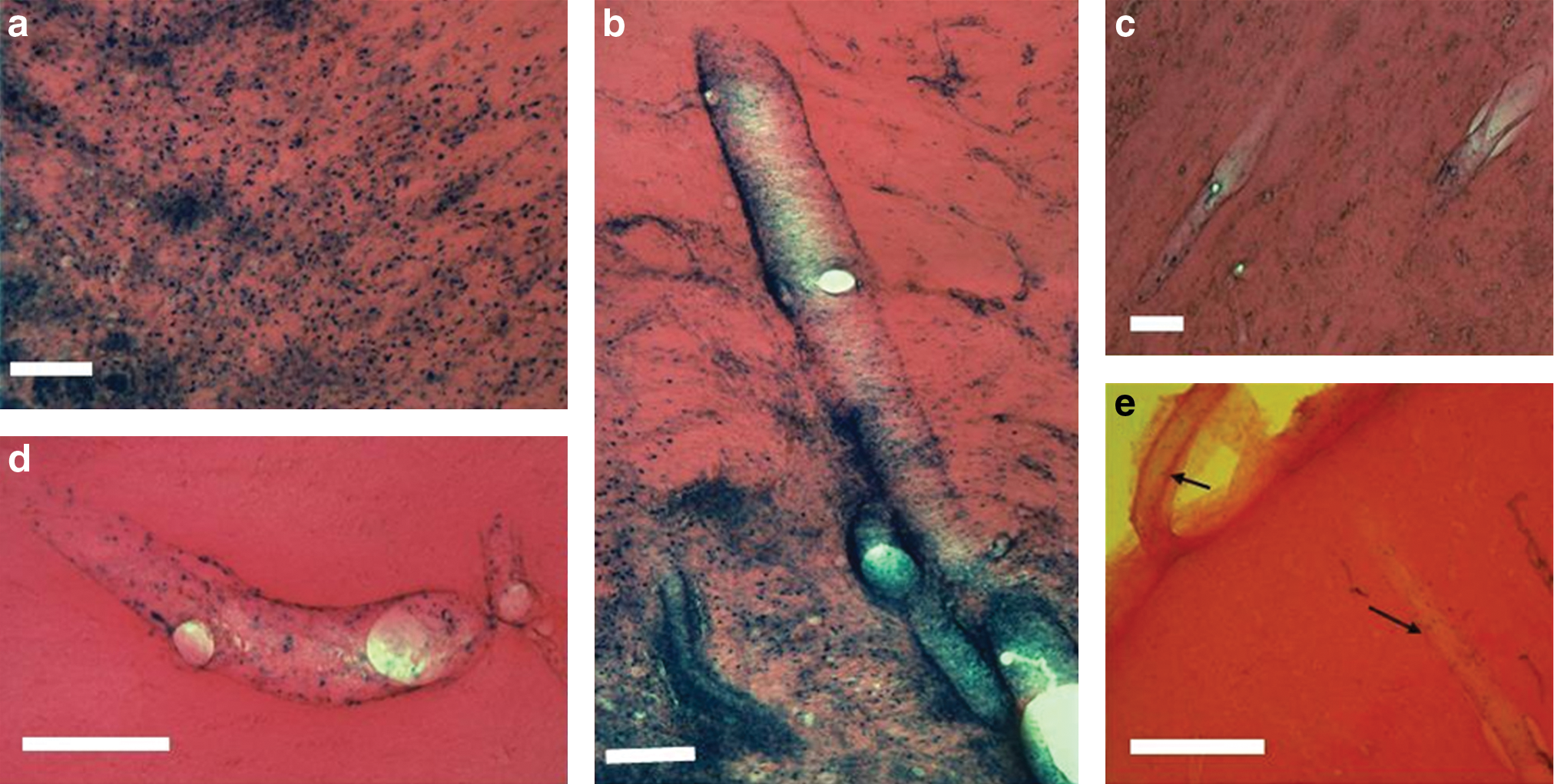
Double immunostaining of tissue sections for β-galactosidase and the neuronal marker NeuN demonstrated that almost all of the transduced cells in the areas of concentrated cell transduction in pig striatum were neuronal (Fig. 11a–c). It was not possible to identify with certainty the type of cells transduced in the perivascular spaces, as the antibody against β-galactosidase used in this study was polyclonal, leading to high levels of background staining. However, whereas there were no identifiable cells within these perivascular spaces that were positive for NeuN, GFAP, ED1, CD8, or the pericyte marker desmin, detailed examination of adjacent sections demonstrated large numbers of HLA-DR-positive cells in a distribution similar to that of β-galactosidase-expressing cells. This finding suggests that transduced cells in the perivascular spaces may be HLA-DR-positive macrophages (Fig. 11d and e). Pig brain sections were examined for evidence of infiltration of ED1-postive microglia and CD8-positive T cells in the vicinity of cell transduction. There was no detectable infiltration of either cell type.

EIAV tropism in pig striatum. Representative images showing extensive β-galactosidase expression in the striatum
Discussion
In this study, convection-enhanced delivery has been evaluated as a strategy to achieve widespread distribution of a prototypical lentiviral vector in the striatum of rat and pig brain. The most striking finding from this study was the massive variability in distribution between infusions despite all infusions being conducted according to a technique identical to that used in a previous study evaluating the distribution properties of vectors derived from a range of serotypes of AAV (White et al., 2011). Whereas we previously demonstrated consistent and reproducible rAAV distribution, in this study EIAV and HIV distribution was extremely variable, implying that the technique itself was not responsible for this inconsistency, but was instead due to intrinsic properties of the lentiviral vectors. A possible explanation could be that lentiviral vectors are significantly larger than rAAV vectors and perhaps tend to distribute through the perivascular spaces, which may be larger than the extracellular space of the neuropil. Nevertheless, the occurrence of widespread distribution of transduced cells in perivascular spaces throughout the brain, in the absence of concentrated cell transduction in the striatum, was concerning and raises major concerns regarding the feasibility of administering lentiviral vectors to the brain.
In view of the enormous variability in vector distribution observed in this study despite the use of a highly standardized procedure, it would have been inappropriate to undertake further infusions. All infusions were performed, using an identical cannula and stereotactic coordinates, four to six times (with animals receiving bilateral infusions) and no trend of consistent vector distribution in each group or between groups was seen
Indeed, approximately one-third of the EIAV infusions and all of the HIV infusions led to negligible levels of concentrated transgene expression in the striatum. From a clinical perspective, this makes the feasibility of the CED-based administration of lentiviral vectors in clinical trials uncertain at present and further investigations are clearly necessary to try and make lentiviral administration by CED more reproducible and consistent.
At the pre- and postcapillary level, perivascular (Virchow-Robin) spaces exist between the outer vascular basement membrane, which ensheathes pericytes and smooth muscle cells, and the glia limitans, which consists of a basement membrane and astrocytic and microglial endfeet. These astrocytic foot processes have been shown to be thinner and less extensive in the gray matter compared with the white matter (Suzuki et al., 2003). At the capillary level, the endothelial basement membrane fuses with the basement membrane of the glia limitans, closing off the perivascular space. The function of the perivascular spaces is analogous to lymphatic drainage from the brain (Cserr and Patlak, 1992; Weller et al., 1992) in that they facilitate drainage of interstitial fluid. This fluid drainage occurs into the subarachnoid space and interestingly has been shown to flow in the opposite direction to blood flow, possibly by the reverse peristaltic wave that follows each pulse wave (Schley et al., 2006).
Hadaczek and colleagues previously identified flow in the perivascular space as being critical in mediating the distribution of infusates, including AAV2 and liposomes, after CED into the rat striatum (Hadaczek et al., 2006). By modulating the blood pressure and pulse of animals they found it possible to vary the distribution of these infusates significantly, such that at high blood pressure and rapid pulse rate distribution was significantly increased. However, in this study, although the blood pressure and pulse of rats were not modulated or indeed monitored during the course of vector infusions, significant variability in the resultant infusion volume was seen. Interestingly, in the case of infusions in which no concentrated cell transduction was observed in the striatum, the cannula track could frequently be visualized terminating on a vessel, suggesting that cannula tip location relative to significant vessels in the brain could be a critical factor in determining vector distribution and consequently the resulting location of transduced cells.
In an attempt to minimize the extent of vector distribution through the perivascular spaces and to reduce the variability of vector distribution, infusions were repeated with heparin or nimodipine. As VSV-G-pseudotyped lentiviral vectors have been shown to bind to heparan sulfate proteoglycans (Guibinga et al., 2002), it was hypothesized that coinfusing vector with a low concentration of heparin would facilitate bulk flow of vector into the brain parenchyma. Indeed, heparin coinfusion has been shown to significantly improve the distribution of AAV2-mediated reporter gene transfer (Nguyen et al., 2001). However, in this study heparin coinfusion was associated with distribution of vector exclusively through the perivascular space for the majority of infusions, although a single infusion did lead to the greatest area of concentrated cell transduction of any infusion of EIAV into rat brain. It is most likely that this discrepancy in distribution resulted from intrinsic variability of cannula tip position relative to vessels within the striatum. This hypothesis is supported by the observation that although one animal had extensive cell transduction unilaterally in the striatum after vector coinfusion with heparin, in the contralateral hemisphere cell transduction was limited exclusively to the perivascular spaces.
By coinfusing the calcium antagonist nimodipine with vector, it was hoped that dilation of vascular smooth muscle would act to partially close the perivascular space, forcing vector into the brain parenchyma. However, when nimodipine was coinfused at a concentration that has been shown to produce significant vasodilation of pial vessels on topical administration (Auer and Mokry, 1986), the volume of distribution of transduced cells fell significantly, providing further evidence that the principal mechanism for vector distribution in the brain is dependent on flow of infusate through the perivascular space.
Interestingly, although this study provides consistent and reproducible evidence of extensive vector distribution through the perivascular spaces, confocal examination of areas of concentrated cell transduction in the striatum suggested that the majority of transduced cell bodies were neuronal. There are a number of potential explanations for this finding. Specifically, it is possible that vector distribution occurred through the extracellular space and not just the perivascular spaces in these animals, perhaps because of the cannula tip being fortuitously placed away from large vessels. Alternatively, had vector passed preferentially through the perivascular spaces it could have transduced axonal terminals innervating cells within the perivascular space and the vector could then have been retrogradely transported to distant neuronal cell bodies. However, the lack of evidence for retrograde or indeed anterograde axonal transport of VSV-G-pseudotyped lentiviral vectors in this and other studies does not support this hypothesis. Alternatively, overwhelming the capacity of the brain to adequately clear vector from the perivascular spaces into the subarachnoid space may result in vector leakage into the parenchymal extracellular space and subsequent transduction of neurons. This hypothesis is supported by observations drawn from infusions of EIAV into the striatum of pigs. Specifically, with these infusions there was an area of concentrated neuronal transduction bilaterally in the putamen. However, there was also extensive cell transduction of cells in the perivascular spaces of vessels spreading radially away from the infusion site, particularly in vessels extending into the frontal cortex, thalamus, and particularly along vessels in the distribution of the lenticulostriate arteries.
The flow of vector through the perivascular space is likely to bring vector into immediate contact with cells of the immune system. Indeed, the perivascular space is populated by macrophages that are regularly replaced by mononuclear cells derived from the bloodstream (Bechmann et al., 2007). Furthermore, perivascular fluid passes retrogradely into the subarachnoid space, from where cerebrospinal fluid may drain to the cervical lymph nodes (Boulton et al., 1999), and along arteries to join the circle of Willis, from where it has the potential to drain to the cervical lymph nodes (Szentistvanyi et al., 1984). As such, in view of the extensive cell transduction in the perivascular space, it was reassuring that EIAV vector delivery into the brain resulted in low levels of immune cell infiltration into the brain, even when vector-mediated cell transduction was restricted entirely to the perivascular spaces. However, the effect of extensive transduction of cells in the perivascular space merits further examination, particularly as these cells may not be terminally differentiated and some may have a number of putative functions that include acting as neural stem cells (Dore-Duffy et al., 2006).
The ability to accurately visualize vector distribution radiologically is likely to be critical in the successful application of gene therapy to the brain. Although it is difficult to accurately compare histological sections that are inevitably distorted by brain removal from the skull and perioperative MR images, there was a detectable but subtle correlation between areas of concentrated cell transduction histologically and areas of high signal on T2-weighted MR images, even though the animal had been allowed to recover for 1 month after the infusion. This offers a practical means of assessing the adequacy of vector distribution within the target structure in clinical practice, without necessitating coinfusion of a radiological tracer such as liposomal gadolinium or iron oxide particles (Szerlip et al., 2007; Fiandaca et al., 2009). Indeed, in contrast to other viral gene therapy vectors such as herpes simplex virus (Marchand and Schwab, 1986) and AAV2 (Passini et al., 2005), VSV-G-pseudotyped lentiviruses are not known to undergo retrograde axonal transport, making it potentially easier to visualize the ultimate distribution of transduced cells.
Previous studies have demonstrated that intraoperative MRI is a useful strategy for real-time visualization of infusate distribution (Mardor et al., 2005; Krauze et al., 2006; Lonser et al., 2007). Furthermore, the distribution of rAAV2 can be estimated intraoperatively through the coinfusion of an MRI contrast agent such as liposomal gadolinium (Fiandaca et al., 2008, 2009) or iron oxide particles (Szerlip et al., 2007). However, in these studies vector infusions led to concentrated cell transduction around the cannula tip, whereas in this study, in many cases cell transduction was restricted to perivascular spaces throughout the brain. In the case of concentrated cell transduction in the striatum, intraoperative MRI is clearly a useful technique. However, when cell transduction is limited to perivascular spaces throughout the brain, which typically occurred around vessels with a diameter significantly less than 100 μm (and therefore impossible to visualize by computed tomography or MR), these techniques are unhelpful, although the use of coinfused contrast agents merits further investigation. Furthermore, it would not be possible to stereotactically insert a cannula in an optimal location relative to such small vessels using existing imaging modalities.
In conclusion, although CED is an effective technique for achieving widespread distribution of many therapeutic agents in the brain, this study suggests that lentiviral vector administration by this technique is highly variable, perhaps because of preferential vector flow through the perivascular spaces. This variability in vector distribution, despite the use of highly standardized conditions, raises significant concerns regarding the feasibility of achieving controlled vector distribution in clinical practice. Considerable further research may be necessary before reproducible lentiviral vector delivery to the brain can be achieved in a clinical setting.
Footnotes
Acknowledgments
The authors acknowledge the financial assistance of the MRC, the Cure Parkinson's Trust, and the Friends of the Bristol Oncology and Haematology Centre. The authors also kindly acknowledge the support of Roy Harris for engineering expertise. In addition, the authors also thank the Neurological Products Division of Renishaw (UK).
Author Disclosure Statement
Edward White, Alison Bienemann, Lisa Megraw, Chotirote Bunnun, Marcella Wyatt, Hannah Taylor, and Steven Gill have read the manuscript and have approved the work to be published in Human Gene Therapy. Steven Gill can confirm that the authors have no conflict of interest and that no competing financial interests exist.