
Abstract
Recombinant adeno-associated virus 5 (rAAV5) represents a candidate vector with unique advantages for the treatment of hepatic disorders because of its narrow hepatic tropism. Noninvasive in vivo imaging of transgene expression provides an important tool with which to quantify the transduction efficiency, and duration and location, of transgene expression. In this study, we used positron emission tomography (PET) and positron emission tomography–computed tomography (PET-CT) imaging to monitor liver transduction efficacy in rodents and nonhuman primates that received rAAV5 vector encoding herpes simplex virus thymidine kinase (HSV-TK). HSV-TK expression in liver was also measured by immunohistochemistry. Notable differences in liver transduction efficiency were found, dependent on the animal species and sex. Male rodents were better transduced than females, as previously described. Moreover, male nonhuman primates also displayed increased hepatic expression of the rAAV5-delivered transgene, indicating that differences in rAAV-mediated liver transduction can be anticipated in humans. Our results demonstrate the high sensitivity and reproducibility of PET, using HSV-TK and [18F]FHBG, to detect gene expression after rAAV vector administration into living animals, confirming the utility of this technology in the quantification of transgene expression, even at low expression levels. However, we also describe how an immune response against HSV-TK hampered analysis of long-term expression in nonhuman primates.
Introduction
The current, most widely used modality to analyze viral vector efficiency and tropism in vivo is the construction of gene transfer vectors expressing the reporter gene luciferase, the expression of which is monitored by live animal bioluminescence imaging (Pañeda et al., 2009). This technique has high sensitivity and specificity; however, these optical techniques are limited because of limited tissue penetration and detailed spatial resolution (Waerzeggers et al., 2009).
Nuclear medicine techniques, such as positron emission tomography (PET) imaging, enable evaluation of gene expression in vivo on the basis of positron emitting radioisotopes, offering the possibility to monitor the exact location, magnitude, and persistence of reporter gene expression. Among the many reporter genes suitable for PET imaging, the thymidine kinase gene from herpes simplex virus 1 (HSV1-TK) has been extensively exploited for imaging gene expression (Willmann et al., 2008). Imaging of HSV1-TK gene expression relies on the use of acycloguanosines, or uracil, derivatives labeled with positron emitters. These molecular probes can traverse the plasma membrane by means of active transport, and are then trapped within the cell via phosphorylation by the viral TK (Yaghoubi and Gambhir, 2006). Cellular retention of radioactivity is, therefore, an indicator of gene expression of TK and can be detected by PET. Fluorine-18-labeled penciclovir analog 9-(4-[18F]fluoro-3-[hydroxymethyl]butyl)guanine, or [18F]FHBG, has been used as an effective substrate for HSV1-TK and evaluated as superior to other agents in many imaging applications and even used in humans (Gambhir et al., 1999; Peñuelas et al., 2002, 2005a,b; Fontanellas et al., 2009). The development of microPET tomographs with a higher resolution has made this technology accessible for noninvasive and quantitative analysis of gene expression in long-term animal studies (Del Guerra and Belcari, 2002). Integrated PET–computerized tomography (CT) machines combine in a single device PET tomography and CT. Both applications are performed simultaneously in the same individual in a single session. PET-CT can be more informative when identifying an anatomic–metabolic correlation to specific locations (Cherry, 2009).
In this study, we used PET imaging to monitor the expression of HSV1-TK in mice, rats, and monkeys after intravenous injection of an rAAV5 encoding this TK under the control of a chimeric liver-specific promoter (Kramer et al., 2003). In nonhuman primates we have combined the use of microPET with clinical PET-CT to determine the biodistribution of the radiotracer. TK expression in the liver was analyzed by immunohistochemistry. Our data show that rAAV5-EalbAAT-TK delivered and mediated expression of TK in the liver. We also show that microPET is highly sensitive when analyzing TK expression in rodents and nonhuman primates. Furthermore, there is a high correlation between TK expression analysis by PET and by immunohistochemistry. The only limitation in using TK as a long-term expression reporter gene is the induction of immune responses in nonhuman primates.
Materials and Methods
Animal experiments
Male and female C57BL/6 mice (age, 8 weeks; weight, 24–26 g) and Sprague-Dawley rats (age, 8 weeks; weight, 160–170 g) were obtained from Harlan Laboratories (Barcelona, Spain). Recombinant AAV were injected intravenously, via the tail vein, in 200 μl of saline solution for mice and 500 μl for rats.
Four young adult cynomolgus monkeys (Macaca fascicularis) of similar age (3 years), with body weights ranging from 2.5 to 4 kg, were obtained from R.C. Hartelist (Tilburg, The Netherlands). Animals were kept in single cages and fed with food and water ad libitum, in standard facilities with controlled temperature and humidity, with a 12-hr on–off light cycle. Animals were anesthetized by intramuscular injections of a mixture of ketamine (Imalgene, 10 mg/kg) and midazolam (Dormicum, 0.8 mg/kg). rAAV-EalbAAT-TK vector was administered via the saphenous vein, in a final volume of 10 ml, at two different doses: 1 × 1012 and 2.5 × 1012 VG/kg. Three weeks and 2 months after injection two liver biopsies were obtained by guided ultrasound imaging, under general anesthesia. After biopsy, monkeys received an intramuscular injection of ketoprofen (Ketofen, 5 mg/kg) and enrofloxacin antibiotic (10 mg/kg) 6, 24, and 48 hr postsurgery.
All animal studies were performed in accordance with guidelines from the institutional ethics commission. The experimental design employing nonhuman primates was approved by the Ethics Committee for Animal Testing of the University of Navarra (Pamplona, Spain) and by the health department of the government of Navarra (ref. no. NA-UNAV-02-08).
Viral construction, production, and purification
Recombinant AAV vectors were constructed with a transgene cassette encoding the reporter gene human herpesvirus 1 strain thymidine kinase (HSV1-TK; GenBank accession no. AF243488) or the reporter gene luciferase (GenBank accession no. M15077) under the regulation of a chimeric liver-specific promoter composed of the human α1-antitrypsin promoter (AAT) with regulatory sequences from the albumin enhancer (Ealb) (Kramer et al., 2003). The porphobilinogen deaminase (PBGD) 3′ untranslated region (UTR) was downstream of the TK or luciferase gene (kindly provided by A. Fontanellas, University of Navarra, Pamplona, Spain). The transgene cassette was flanked by AAV2 wild-type inverted terminal repeats and the entire cassette was subcloned into the baculoviral transfer plasmid to generate recombinant baculovirus. Pseudotyped AAV2/5 vectors were produced with a baculovirus expression vector system (Protein Sciences, Meriden, CT) as described (Urabe et al., 2006). Baculoviral stocks expressing the rAAV genome, VP capsid proteins, and Rep proteins were used for AAV production by triple infection of spodoptera frugiperda (Sf+) insect cells, at a ratio of 1:1:5.
AAV purification was performed by affinity chromatography, using AAV-specific llama heavy chain variable domain (VHH) antibodies (Hermens et al., 1999; Zolotukhin et al., 1999). Viral titers in terms of genome copies per milliliter (GC/ml) were determined by quantitative polymerase chain reaction (qPCR) (Pañeda et al., 2009). Capsid protein composition and purity were analyzed by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE) and endotoxin test. Ad-CMV-TK was constructed as described by Peñuelas and colleagues (2005b).
Analysis of TK expression by microPET and PET-CT
PET imaging was performed in a dedicated small animal tomograph (MOSAIC HP; Philips, Cleveland, OH) with 2-mm resolution. The technique is based on the injection of a reporter probe, [18F]FHBG, prepared as previously described (Peñuelas et al., 2005b). For rodent studies, general anesthesia was induced with 2% isoflurane in 100% O2 gas. Subsequently, radiotracer was injected via the tail vein (mice, 14.1 ± 7.4 MBq in 100 μl; rats, 37 ± 15 MBq in 200 μl). Animals were immediately awakened for radiotracer uptake and placed back in a cage for 60 min. Afterward, animals were placed prone on the PET scanner bed to perform a static acquisition for 15 min under continuous inhalational anesthesia. In the case of rats, a transmission scan was performed after image acquisition. After PET studies, mice and rats were killed for liver collection and immunohistochemistry (IHC) for TK.
For monkey microPET studies, animals were fasted overnight before PET. On the day of the scan, anesthesia was initially induced with ketamine (10 mg/kg, intramuscular) and maintained throughout the procedure with ketamine and midazolam (10 and 0.8 mg/kg, respectively). After positioning the animal prone in the scanner, a transmission study was carried out for 10 min. Radiotracer with an activity of 75 ± 17 MBq was then injected into the saphenous vein and after 60 min, static acquisition was carried out for 15 min. In some cases, after microPET studies and in order to obtain additional information about the anatomical distribution of [18F]FHBG, monkeys were moved to a hybrid PET-CT tomograph (Biograph Duo; CTI Molecular Imaging/Siemens, Knoxville, TN). CT was performed 1 min before PET scan, with the animal in the same position. CT parameters for a dual-detector helical CT were 130 keV, 50 mA, 0.8 sec per CT rotation, 5-mm slice thickness, and a pitch of 1.5. PET images were acquired in three-dimensional mode with 3 min per bed position. CT images were reconstructed with 2.4-mm increments and PET emission data were corrected for random events, dead time, and attenuation correction and were reconstructed with an OSEM (ordered-subsets expectation maximization) algorithm.
All the microPET images, both for rodents and monkeys, were reconstructed using a three-dimensional row-action maximum likelihood algorithm (RAMLA) with a 1-mm voxel size, applying dead time, decay, random and scattering corrections, and also attenuation correction for rats and monkeys.
Image quantitative data for the PET are expressed as maximal standard uptake value (SUVmax). To obtain this value, regions of interest (ROIs) were drawn over the liver parenchyma and maximal SUV values were calculated according to the following formula: [tissue activity concentration (Bq/cm3)/injected dose (Bq)] × body weight (g).
Ex vivo processing of liver biopsies
Liver biopsies were processed for TK immunohistochemistry staining with a primary polyclonal antibody (kindly provided by W. Summers, Yale University, Ithaca, NY), using the EnVision+ system (Dako, Glostrup, Denmark) according to the manufacturer's recommendations and as described (Fontanellas et al., 2009).
Total DNA was extracted from liver biopsies, using a QIAamp DNA mini kit from Qiagen (Chatsworth, CA) in a laminar flow cabinet. Viral DNA was quantified by qPCR, using the primers TK-sense (TCCCATGCACGTCTTTATCCT) and TK-antisense (TAAGTTGCAGCAGGGCGTC). Monkey β-actin was quantified with the following primers: sense (CCTGTGGCATCCACGAAAC) and antisense (TTACGGATGTCCACGTCACACT). Rat glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was quantified with the following primers: sense (CTTCCACGATGCCAAAGTTC) and antisense (GATGGTGAAGGTCGGTGTG) and mouse GAPDH was quantified with the following primers: sense (TGCACCACCAACTGCTTA) and antisense (GGATGCAGGGATGATGTTC). The number of TK copies in each biopsy was normalized with β-actin or GAPDH copy numbers. The values shown correspond to the mean copy number from two independent biopsies. Liver samples were stained with hematoxylin and eosin (H&E) for histopathological evaluation.
TK protein expression and purification
The HSV1-TK gene was amplified by PCR from plasmid pSK-TK. The resulting PCR product was directly cloned into the multiple cloning site of pcDNA3.1/V5/His-TOPO, using the TOPO cloning kit (Invitrogen, Carlsbad, CA) and positive clones were verified by sequencing. The TK gene was subcloned into the pET-14b vector (Novagen/EMD Biosciences, Madison, WI), which enables expression of the thymidine kinase protein carrying six histidine residues (His•Tag sequence) at the N terminus. The resulting plasmid pET14b-TK was transformed into BL21 competent Escherichia coli (Sigma-Aldrich, St. Louis, MO) for expression of the recombinant protein. Protein was purified from the supernatant by affinity chromatography (HisTrap; GE Healthcare Life Sciences, Piscataway, NJ), using a fast protein liquid chromatography platform (ÄKTA
Analysis of T cell-specific immune response in monkeys
To evaluate the cellular immune response induced against the vector or the transgene after rAAV5-EalbAAT-TK vector administration, monkey leukocytes were purified from peripheral blood by centrifugation through Ficoll-Hypaque (GE Healthcare Life Sciences). Isolated leukocytes (5 × 105 cells/ml) were cultured in medium alone or in the presence of purified rAAV5 capsid proteins (10 μg/ml), purified recombinant TK protein (10 μg/ml), or phorbol myristate acetate (PMA, 0.05 μg/ml) plus ionomycin (0.5 μg/ml) (P/I), in triplicate. After 48 hr cells were harvested, RNA was extracted, and interferon (IFN)-γ and GAPDH expression was analyzed by quantitative RT-PCR, using the primers IFN-γ-sense (5′-TGGAGACCATCAAGGAAGACA-3′) and IFN-γ-antisense (5′-ACAGTTCAGCCATCACTTGGA-3′), and GAPDH sense (5′-GGTCGGAGTCAACGGATTT-3′) and GAPDH antisense (5′-CCAGCATCGCCCACTTGA-3′).
Results
TK expression analysis by microPET and IHC in mice
C57BL/6 mice of both sexes were injected intravenously with two different doses of rAAV5-EalbAAT-TK vector: 5 × 1012 and 1 × 1012 GC/kg. Three weeks after injection, mice were subject to imaging for TK reporter gene expression by PET, after the administration of [18F]FHBG substrate. After PET analysis, all the animals were killed and liver sections were subjected to IHC for TK. As a negative control, two mice received a 5 × 1012 GC/kg dose of an rAAV encoding luciferase (rAAV5-EalbAAT-Luc).
Representative microPET imaging results are shown in Fig. 1A. Expression of the TK reporter gene was confined to the liver. Washout of the tracer to the intestines and gallbladder was detected. The signal intensity was dose dependent and was consistently stronger in males than in females (also shown in Fig. 1B). Representative images of TK IHC analysis are shown in Fig. 1C and the mean values of the percentage of transduction for each group of animals appear in Table 1. The analysis of TK expression by IHC showed a high correlation between PET SUVmax values and the percentage of transduced hepatocytes (r 2 = 0.96; Fig. 1D). Importantly, our data show that PET technology can detect TK hepatic transduction levels in mice down to 1% efficiency. Of note, no retention of radioactive tracer in the liver was observed in mice injected with rAAV5-EalbAAT-Luc (data not shown).

In vivo and ex vivo analysis of the expression of the HSV-TK reporter gene in C57BL/6 female and male mice after injection of rAAV5-EalbAAT-TK via the tail vein. (
SUVmax, maximal standard uptake value.
One of the main characteristics of rAAV vectors is their capacity for sustained transgene expression. To monitor long-term expression of TK after rAAV administration, three C57BL/6 female mice were injected with 1 × 1012 GC/kg; 4 and 8 weeks later, TK expression was analyzed by microPET. As shown in Table 2, similar SUVmax values were obtained after the first and second measurements, indicating that TK expression was stably maintained after rAAV injection. After the second analysis, animals were killed and TK expression was analyzed by IHC. Maximal SUV values correlated with the percentage of transduced hepatocytes (data not shown). The same experiment was performed in C57BL/6 male mice, obtaining similar results (data not shown).
GC, genome copies; SUVmax, maximal standard uptake value.
TK and luciferase expression analysis in rats
Four male and four female Sprague-Dawley rats were injected intravenously, via the tail vein, with rAAV5-EalbAAT-TK at a dose of 1 × 1012 GC/kg. One male rat and one female rat were injected with the corresponding dose of rAAV5-EalbAAT-Luc (negative control) and as a positive control one male was injected with adenoviral vector encoding TK under the control of a cytomegalovirus promoter (Ad-CMV-TK) at a dose of 5 × 1011 plaque-forming units (PFU)/kg. TK expression was analyzed by microPET 1 and 3 weeks after AAV injection and 2 days after adenovirus injection. Representative microPET imaging results are shown in Fig. 2A. A strong signal was detected in the animals injected with the adenovirus expressing TK, whereas a faint signal was detected in male rats injected with rAAV5-EalbAAT-TK. Female rats injected with rAAV5-EalbAAT-TK or rAAV5-EalbAAT-Luc demonstrated similar levels of radiotracer retention. Similar results were obtained 1 and 3 weeks after AAV injection. These results indicate that at this dose rAAV5 does not transduce the liver of female rats (Fig. 2A). IHC corroborated the absence of liver transduction in female rats and a low transduction level (<0.15%) in male rats. A large proportion of hepatocytes was transduced in rats administered adenovirus expressing TK (29%; Fig. 2B). To confirm the lower transduction efficiency of AAV5 in Sprague-Dawley rats the presence of viral genomes in the liver was quantified by qPCR in rats and in C57BL/6 mice injected with the same dose of virus. As shown in Fig. 2C, the number of genomes in the liver correlates with TK expression levels. To determine whether the lower transduction level was due to the development of an immune response against the virus or the transgene, lymphocytes obtained form rat peripheral blood 21 days after vector injection were stimulated with recombinant TK protein or AAV5 empty capsids, as positive control lymphocytes were stimulated with concanavalin A. Twenty-four hours after stimulation lymphocytes were harvested, RNA was extracted, and IFN-γ expression was analyzed by qPCR. No IFN-γ was detected after stimulation with AAV5 capsid or recombinant protein (data not shown). To further confirm the lower transduction level mediated by AAV serotype 5 in Sprague-Dawley rats in comparison with C57BL/6 mice, four animals of each species were injected with an AAV5 virus expressing luciferase (rAAV5-EalbAAT-Luc) at a dose of 2.5 × 1012 VG/kg and transgene expression was monitored for 1 month. As shown in Fig. 2D, luciferase expression was sustained over time in both groups, but luciferase expression was 10 times higher in C57BL/6 mice.
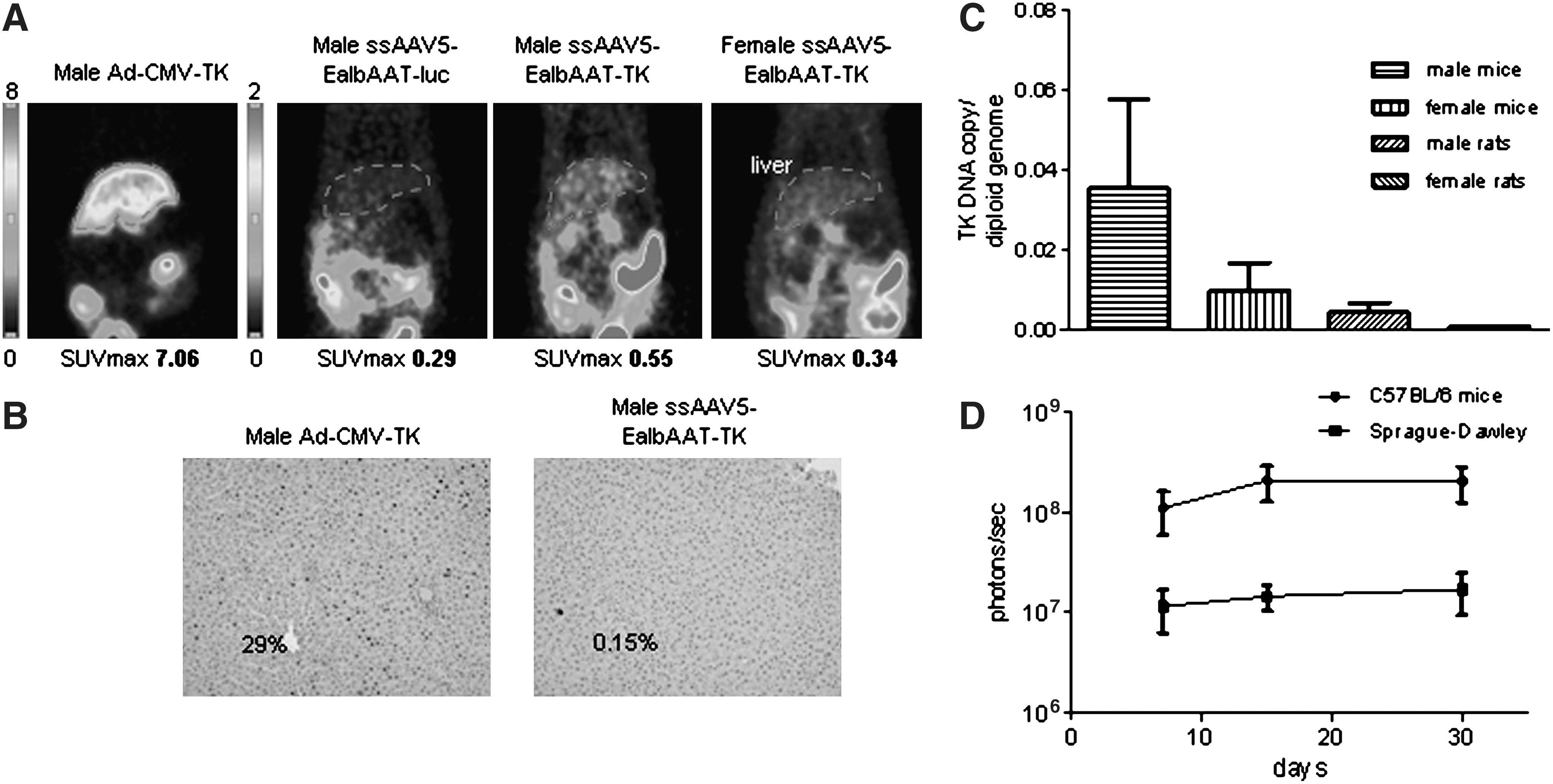
In vivo analysis of the expression of the HSV-TK reporter gene in Sprague-Dawley rats after injection via the tail vein of Ad-CMV-TK, rAAV5-EalbAAT-TK, or rAAV5-EalbAAT-Luc. (
TK expression analysis by microPET and IHC in nonhuman primates
Two male and two female monkeys were injected with rAAV5-EalbAAT-TK by intravenous injection. One male (male low dose, MLD) and one female (female low dose, FLD) received a low dose of 1 × 1012 GC/kg, and one male (male high dose, MHD) and one female (female high dose, FHD) received a high dose of 2.5 × 1012 GC/kg. rAAV5 vector delivery led to no untoward clinical, hematological, or serum chemistry responses in macaques (Pañeda et al., Unpublished observation). TK expression analyses by microPET were performed before and 15, 30, and 60 days after rAAV5-EalbAAT-TK injection, after [18F]FHBG substrate administration. Liver biopsies were taken 30 and 70 days after virus injection. All the animals were killed 3 months after vector injection.
Figure 3A illustrates microPET imaging from the studies carried out in monkeys before (basal studies) and 15 days after injection of the vector. Specific retention of the radiotracer due to TK activity is confined to the liver. A strong signal can also be detected in the intestines and gallbladder, due to metabolic washout of the radiotracer. MicroPET analysis was repeated 30 and 60 days after vector injection. Quantitative analysis of radioactive tracer retention in the liver was estimated by region of interest (ROI) measurement in the liver, using static microPET images, and expressed as SUVmax. As shown in Fig. 3B, radiotracer incorporation was maximal 15 days after vector administration and correlates with the dose of rAAV5-EalbAAT-TK injected. SUVmax values were higher in males than in females at the same dose, as has been previously described in rodents (Pañeda et al., 2009). However, contrary to what we observed in mice, expression was not maintained over time in monkeys, in which after 30 days TK expression decreased and returned to background levels 1 month later (60 days). TK expression was analyzed by IHC in liver biopsies and in whole liver after sacrifice. Low or no TK expression was detected in liver biopsies as shown in Fig. 3C: 0.039% in MLD; 0.59% in FLD, 0.79% in FHD, and 0 in MHD. Surprisingly, transduction efficiency was higher in females than in males. In fact, there was an inverse correlation between the TK expression level detected by PET and by IHC. A possible explanation for this finding is that the more robust transgene expression levels observed in male monkeys induce a stronger immune response, resulting in faster disappearance of the transgene. Analysis of the biopsy obtained 70 days after vector injection, or at sacrifice, showed no TK expression in any of the animals, confirming the result obtained by microPET. Analysis of viral DNA in liver biopsies by qPCR revealed the disappearance of vector genomes from the liver (Fig. 3D). Taken together, the data suggest that rAAV5-EalbAAT-TK-transduced hepatocytes were eliminated, most likely due to an immune response that appears to have developed faster in male monkeys.
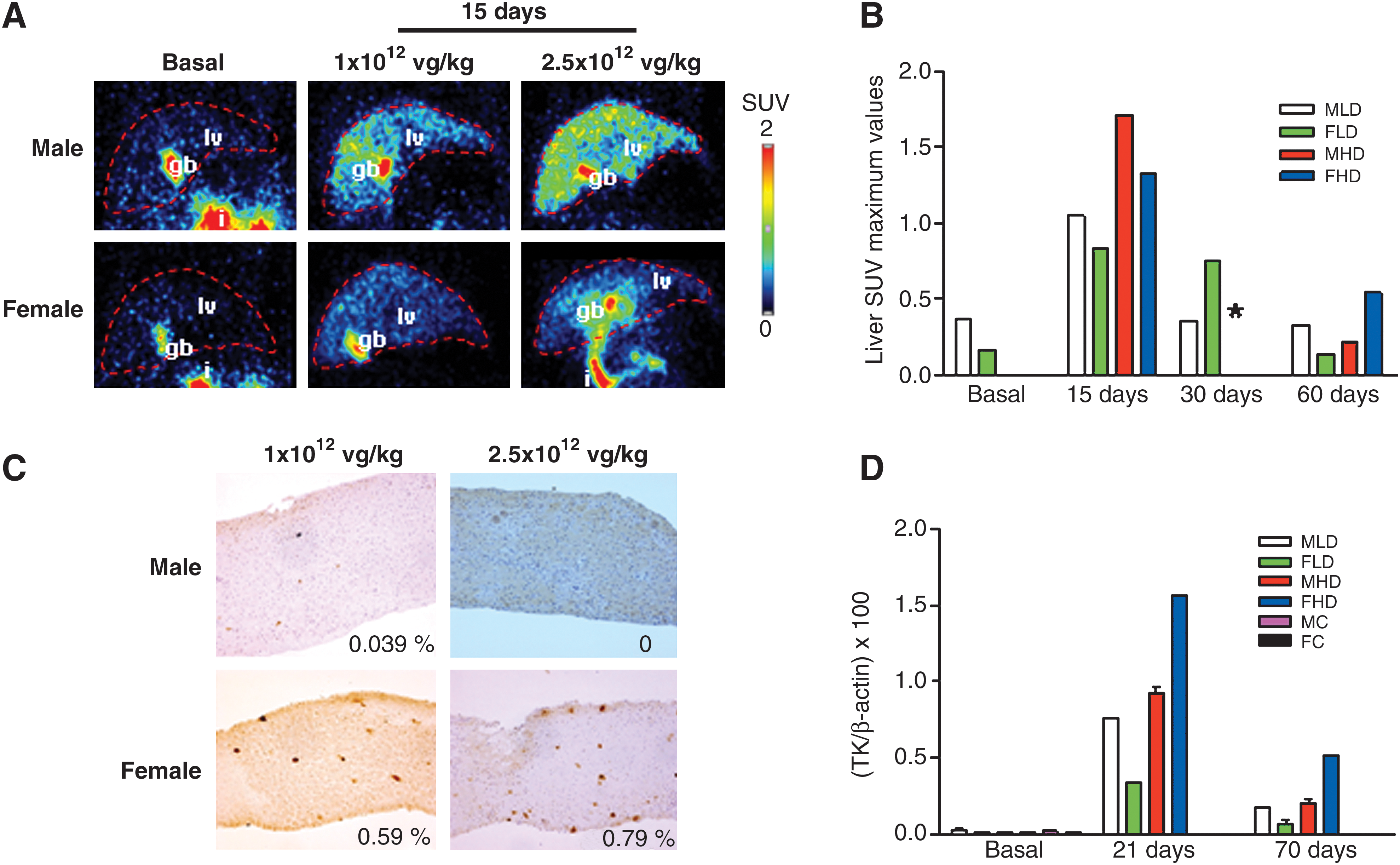
In vivo analysis of the expression of the HSV-TK reporter gene in Macaca fascicularis monkeys after injection of rAAV5-EalbAAT-TK into the saphenous vein. (
PET-CT data in nonhuman primates
All monkeys were analyzed in a clinical PET-CT scanner 15 days after rAAV5-EalbAAT-TK administration and 2 hr after the microPET study was performed. This study, performed 3 hr after radiotracer injection, should provide detailed information about the biodistribution of TK expression, because a whole body CT scan is superimposed over the PET image. Figure 4 illustrates PET-CT images of male and female monkeys that received high doses. For both sexes, the radioactive tracer was detected in liver by virtue of rAAV-delivered TK (and in intestines and bladder due to washout of the radiotracer).
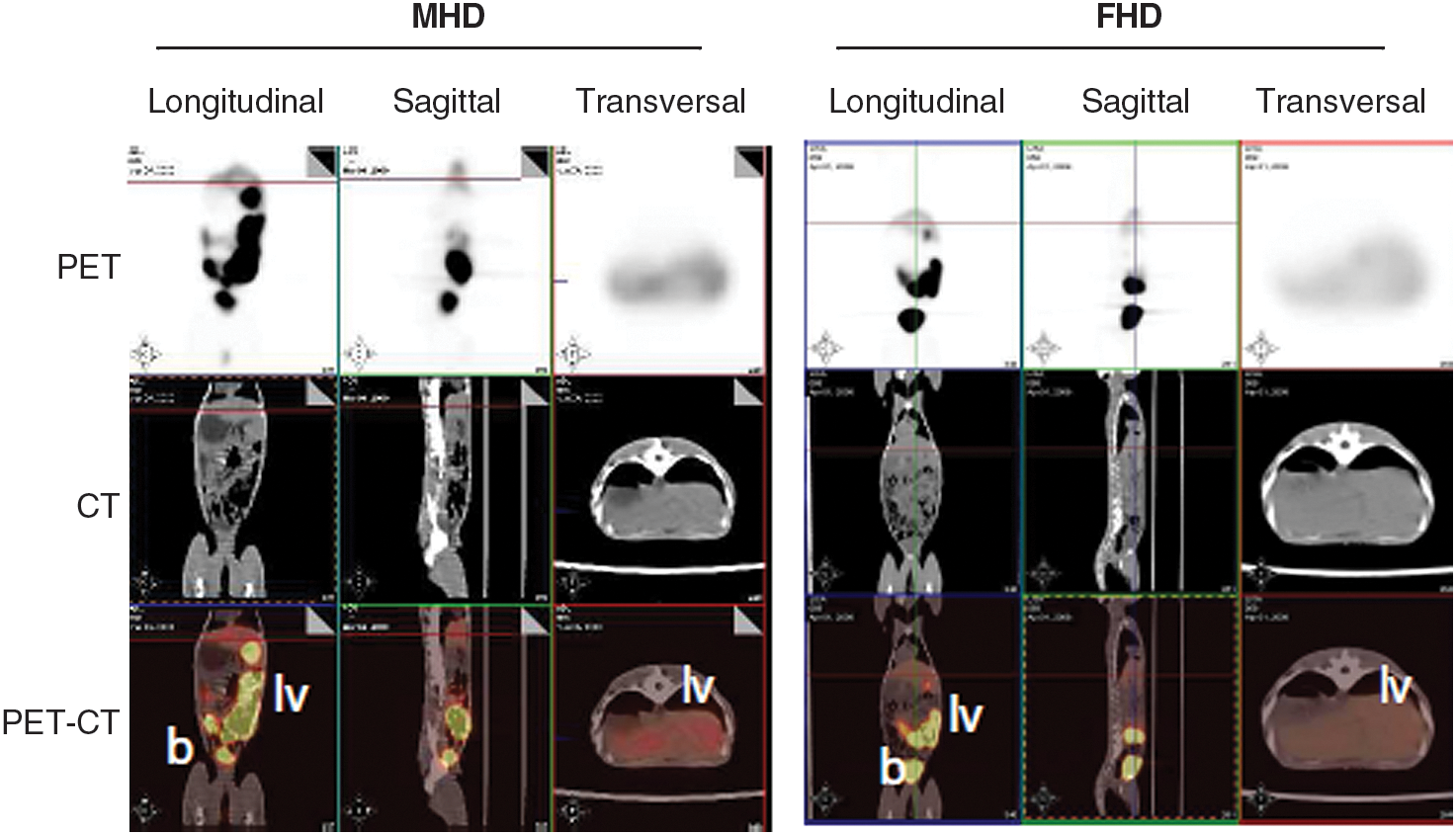
PET-CT images of male and female monkeys 15 days after injection of rAAV5-EalbAAT-TK (2.5 × 1012 GC/kg), and 2 hr after administration of the radioactive tracer. lv, liver; b, bladder.
Analysis of immune response against TK and AAV5 capsid proteins
The disappearance of TK expression from the liver of rAAV5-EalbAAT-TK-injected monkeys suggests an immune response against the recombinant vector, with the non-self proteins being the recombinant TK protein and the viral capsid proteins. Before and every month after virus administration, lymphocytes were obtained from each monkey. At the end of the study, the lymphocytes were incubated with purified rAAV5 capsid proteins, purified recombinant TK protein, PMA plus ionomycin (P/I), or medium alone. Forty-eight hours after incubation with the various stimuli, cells were harvested and IFN-γ and GAPDH expression was analyzed by qPCR. No IFN-γ production was detected before vector administration after stimulation with recombinant TK protein of viral capsid protein. However, as shown in Fig. 5A, IFN-γ production was detected after incubation with recombinant TK protein but not with AAV5 capsid proteins in lymphocytes obtained from the four monkeys after virus injection. Interestingly, the immune response was stronger and could be detected for a longer time in male rather than female monkeys. To determine whether the immune response resulted in a toxic effect, complete serum biochemistry analysis was performed 7, 15, 30, 60, and 90 days after viral injection. Urea, bilirubin, alkaline phosphatase (ALP), aspartate aminotransferase (AST), alanine aminotransferase (ALT), total protein, albumin, γ-glutamyltransferase (GGT), creatinine, Na+, Cl–, K+, glucose, cholesterol, calcium, and creatine phosphokinase (CPK) concentrations in serum were determined. All values were in the normal range except for AST, which was elevated on days 7 and 15 after vector injection into the MHD monkey, and on day 30 in the rest of the animals (Fig. 5B). Thus, an immune response against the transgene was deemed more likely to be responsible for the disappearance of transduced hepatocytes.

Analysis of TK and AAV5 capsid immune response and transaminase levels in serum. (
Discussion
Recombinant AAV vectors are currently among the most frequently used viral vectors for gene therapy (Grieger and Samulski, 2005; Alexander et al., 2008; Daya and Berns, 2008; Terzi and Zachariou, 2008; Luebke et al., 2009; Stieger et al., 2009; Markakis et al., 2010). Because human populations are natural hosts for AAV one of the limitations for the clinical use of rAAV is the presence of anti-AAV neutralizing antibodies. The use of various pseudotyped AAV capsids (serotypes) has allowed for broad tissue tropisms and avoidance of preexisting immunity against AAV (Boutin et al., 2010; Markakis et al., 2010). As the prevalence of neutralizing antibodies against AAV5 and AAV8 is lower in humans it is likely that these serotypes may be useful in patients with high-titer neutralizing anti-AAV2 antibodies (Boutin et al., 2010).
Molecular imaging plays a major role in the field of gene therapy, allowing quantitative monitoring of the magnitude of gene expression, as well as analysis of the location and duration of expression over time (Gambhir et al., 1999; Peñuelas et al., 2002, 2005a,b; Fontanellas et al., 2009). Radionuclide imaging methods such as PET have excellent depth sensitivity and can detect accumulation of gene expression within vector-transduced organs anywhere in the body. When combined with other forms of tomographic imaging, such as CT, fusion images of functional and anatomic data provide more detailed in situ information about reporter gene expression and localization. In this study, rAAV5 expressing herpes simplex virus thymidine kinase under the control of a liver-specific promoter (rAAV5-EalbAAT-TK) was used to monitor rAAV5-mediated gene expression and duration in nonhuman primates.
First, we tested the sensitivity and robustness of this technique in rodent models. We found a direct correlation between SUVmax values obtained after PET analysis of rAAV5-EalbAAT-TK-injected mice with the liver transduction efficiency calculated by immunohistochemistry as TK-positive cells (r 2 = 0.957). Our results are in agreement with previous studies performed by Liang and colleagues (2002), who reported a linear correlation between hepatic FHBG retention and HSV-TK activity in liver extracts of mice injected with adenovirus encoding TK (r 2 = 0.96) (Liang et al., 2002). Because rAAV vectors have been reported to mediate long-term expression in rodents and nonhuman primates, it was considered important to monitor TK expression over time, noninvasively, by PET imaging. Our results indicate that in rodents TK expression is maintained for at least 2 months after vector injection. We found dramatic differences in liver transduction depending not only on the sex, as previously reported, but also on the species. After a single injection of 1 × 1012 GC/kg in C57BL/6 male mice, 3% of the hepatocytes were transduced, whereas only 0.05% of the hepatocytes from Sprague-Dawley male rats showed TK expression. Despite the low transduction efficiency of hepatocytes, TK activity was detected and quantified by microPET in these rats, demonstrating the high sensitivity of this technique to detect gene expression in living subjects.
rAAV-mediated gene transfer studies in nonhuman primates are not abundant (Nathwani et al., 2002, 2006, 2007; Davidoff et al., 2005; G. Gao et al., 2006; G.P. Gao et al., 2006; Jiang et al., 2006). In rhesus macaques, injection of rAAV serotypes 2, 5, and 8 expressing factor IX, recombinant human chorionic gonadotropin (rhCG), or erythropoietin (Epo) resulted in long-term expression of the transgene (G.P. Gao et al., 2006; Jiang et al., 2006; Nathwani et al., 2007). In our study, four monkeys were injected with two different doses of rAAV5-EalbAAT-TK and were analyzed on days 15, 30, and 60 after viral administration by microPET after [18F]FHBG administration. We found that radiotracer incorporation could be detected in the liver of rAAV5-EalbAAT-TK-injected monkeys 15 days after vector injection. PET-CT analysis showed that TK expression was restricted to the liver. The signal intensity depended on the dose of virus administered and on the sex of the monkey. Interestingly, the intensity of radiotracer incorporation was higher in male than in female monkeys, as previously shown in rodent models. This observation is particularly important, because macaques are more likely than murine or other small animals to reflect the transduction efficacy of AAV vectors in humans. Thus, differences dependent on sex would also be expected in humans and should be taken into account when designing clinical trials.
Long-term analysis showed that, contrary to the long-term expression of human factor IX or rhCG observed in nonhuman primates (NHPs) (Nathwani et al., 2002, 2006, 2007; Davidoff et al., 2005; G. Gao et al., 2006; G.P. Gao et al., 2006; Jiang et al., 2006), expression of TK disappeared from the liver of monkeys injected with rAAV5-EalbAAT-TK. PET analysis performed on days 30 and 60 after viral injection showed a significant decrease or disappearance of radiotracer signal from the liver, compared with day 15. Immunohistochemistry, performed on day 21, confirmed the disappearance of TK expression in male monkeys that received the highest dose of rAAV5 vector and also showed low expression in the rest of the monkeys. Analysis of liver biopsies obtained on day 60 and from different liver lobes (on sacrifice) revealed the disappearance of TK expression from all treated monkeys. This was corroborated by the loss of vector genomes.
Interestingly, the expression levels on day 21 were lower in males than in female monkeys, in contradiction with the microPET data. Because we have observed a direct correlation between TK expression analyzed by IHC and by PET in rodents, our explanation for the discrepancy in NHPs is that the factor responsible for the disappearance of TK-transduced cells developed faster in males than in females.
These results suggest that administration of the rAAV5-EalbAAT-TK virus has induced the development of a specific immune response that eliminated the transduced hepatocytes. Because the only foreign components of the virus are the transgene and capsid proteins, we have analyzed the production of IFN-γ by the lymphocytes after stimulation with rAAV5 capsid proteins and recombinant TK protein. We have observed IFN-γ production only after incubation with TK, indicating that an immune response against the transgene had developed. The more robust transgene expression in male monkeys would likely make TK more immunogenic, explaining why gene expression is more rapidly lost in males. Furthermore, analysis of transaminase levels in serum showed a transient elevation of AST, indicating a certain degree of liver injury that developed in the male monkey receiving the highest dose (MHD). Our results correlate with those described by Gao and coworkers, where primates elicited vibrant cytotoxic T cell responses to green fluorescent protein (GFP) that correlated with hepatitis and loss of transgene expression (Gao et al., 2009; Wang et al., 2010). These studies indicate that, under some conditions, primates may activate more robust T cell responses to transgene products than mice. Furthermore, Mercier-Letondal and colleagues showed that the administration of HSV1-TK-expressing, gene-modified T cells to patients induces a T cell response, preferentially targeting TK, that eliminates the transduced cells (Mercier-Letondal et al., 2008).
In summary, PET technology is a noninvasive method with high value to analyze vector transduction with the reporter gene thymidine kinase in mice and rats. The quantitative SUVmax value is correlated with the transduction efficiency estimated by IHC. TK expression can be detected by microPET and PET-CT in the liver of monkeys after AAV5-mediated gene transfer. However, because of the immunogenicity of the transgene, TK expression was detected only transiently. This model could be useful for the development of new strategies to circumvent immune response against the transgene in NHPs.
Footnotes
Acknowledgments
The authors are grateful to Dr. William Summers for the anti-HSV-TK antibody, and to Izaskun Bilbao and Margarita Ecay for microPET imaging. The PET laboratory crew is acknowledged for FHBG synthesis. The authors thank Cristina Olagüe, Africa Vales, Nora Arriazu, and Roberto Ferrero for excellent technical assistance, as well as Javier Guillén, Elena Ciordia, Alberto Espinal, and Susana Rosell for animal care and vivarium management. Dr. Lanciego is acknowledged for supportive advice. Dr. Lasarte and Marta Martínez are acknowledged for help with the production of recombinant HSV-TK. This work was funded in part by grants from the UTE project CIMA, and by grants SAF2006-03623 and SAF2009-08524 from the Spanish Department of Science and Fundación Mutua Madrileña.