
Abstract
Farber disease is a rare lysosomal storage disorder (LSD) that manifests due to acid ceramidase (AC) deficiencies and ceramide accumulation. We present a preclinical gene therapy study for Farber disease employing a lentiviral vector (LV-huAC/huCD25) in three enzymatically normal nonhuman primates. Autologous, mobilized peripheral blood (PB) cells were transduced and infused into fully myelo-ablated recipients with tracking for at least 1 year. Outcomes were assessed by measuring the AC specific activity, ceramide levels, vector persistence/integration, and safety parameters. We observed no hematological, biochemical, radiological, or pathological abnormalities. Hematological recovery occurred by approximately 3 weeks. Vector persistence was observed in PB and bone marrow (BM) cells by qualitative and quantitative PCR. We did not observe any clonal proliferation of PB and BM cells. Importantly, AC-specific activity was detected above normal levels in PB and BM cells analyzed post-transplantation and in spleens and livers at the endpoint of the study. Decreases of ceramide in PB cells as well as in spleen and liver tissues were seen. We expect that this study will provide a roadmap for implementation of clinical gene therapy protocols targeting hematopoietic cells for Farber disease and other LSDs.
Introduction
Gene therapy for Farber disease is feasible. Our prior studies have shown correction of enzymatic activity in immortalized Farber patient cells by transduction with a recombinant oncoretrovirus that engineered expression of human AC (Medin et al., 1999). We also showed that AC activity produced by transduced cells could be taken up by nontransduced bystander cells through the mannose-6-phosphate receptor pathway, a phenomenon described as “metabolic co-operativity” or “cross-correction.” This allows for a lower number of functionally transduced cells to have a systemic therapeutic effect. Recently, we showed that novel bicistronic oncoretroviral vectors and lentiviral vectors (LVs) could be used to generate overexpression of AC activity in Farber patient fibroblasts and B cells, leading to reductions in ceramide levels (Ramsubir et al., 2008). We also demonstrated that direct neonatal LV injections and LV-mediated transduction of human hematopoietic stem/progenitor cells (HSPCs) followed by transplantation into a murine xenotransplant model both provided sustained transgene activity (Ramsubir et al., 2008). Indeed, LVs have the ability to transduce more slowly dividing cell populations and are effective for stable gene transfer into HSPCs. Furthermore, our LV construct is also bicistronic, incorporating the human AC cDNA along with the cDNA for human CD25, which can be used to enrich transduced cells (Qin et al., 2001) and as a safety mechanism against genotoxicity (Ramsubir et al., 2007). Thus, a one-time gene therapy approach employing such a delivery vehicle may provide a lasting and safe treatment for Farber disease.
Here we tested this gene therapy strategy for Farber disease in rhesus macaques, employing LV transduction and transplantation of primary hematopoietic cells. We followed animals for at least 1 year post-transplantation to track the sustainability of the transgene in peripheral blood (PB) and BM cells. Hematological and biochemical parameters were followed to assess the recovery and health status of the animals. No untoward effects were observed. Importantly, we observed increases of AC-specific activity to supranormal levels in PB cells, BM cells, spleen, and liver throughout the study period. Additionally, ceramide levels were reduced due to increased AC activity even in these metabolically normal animals. Our results show that transplantation of LV-transduced mobilized PB cells is safe and may be efficacious for Farber disease and, by extension, other LSDs.
Material and Methods
Animals
Three male rhesus macaques, Macaca mulatta, (weight 5.0 ± 0.5 kg; 3, 6, and 7 years old, respectively) were housed individually within a negative-pressure regulated animal facility at the University Health Network (UHN). All experimental protocols and animal care procedures were approved by the UHN Animal Care Committee. All animals tested serologically negative for SIV, STLV, simian retrovirus, and herpes virus B. Animals were also routinely monitored for other infectious pathogens including tuberculosis. Telemetric devices (Data Sciences International) and vascular access ports (Access Technologies) were implanted to monitor vital functions and permit venous access, respectively. Animals were euthanized with an overdose of sodium pentobarbital. Control tissue samples were also obtained from nine other rhesus macaques of similar sizes involved in unrelated studies (weight 5.0 ± 1.0 kg; 4–7 years old).
Cell mobilization and harvest
Animals were injected intravenously with Granulocyte-Colony Stimulating Factor (G-CSF) (kindly provided by Amgen) at a dose of 10 g/kg/d for 5 days. Mobilized PB mononuclear cells were harvested by leukapheresis using a clinically applicable and commercially available COBE Spectra AutoPBSC apheresis machine with minor modifications (Donahue et al., 1996). Cells were suspended in RPMI-1640 media (Sigma Aldrich) containing 10% dimethylsulfoxide (Sigma-Aldrich) and 50% fetal calf serum (PAA Laboratories, Inc.), and stored frozen in liquid nitrogen until use.
Tranductions, transplantation, and post-transplantation care
The bicistronic HIV-1-based LV pHR′/huACer/IRES/huCD25 and endotoxin-free virus production have been described previously (Ramsubir et al., 2008). Viral supernatant was concentrated approximately 200-fold, titered on 293T cells for functional huCD25 expression, and stored at −80°C until use. Transduction and transplantation methods were modified from previously studies (Donahue et al., 1996; An et al., 2000, 2001; Kiem et al., 2004). Mobilized PB mononuclear cells were thawed and prestimulated for 24 hr with 100 ng/ml rhuSCF, 100 ng/ml rhuFlt3-L, 20 ng/ml rhuIL-6 (all kindly supplied by Amgen) and rhuTPO (50 ng/ml) (R&D Systems). The cells were then transduced once with an overnight incubation at an approximate multiplicity of infection (MOI) of 15, 7.5, and 4, for the first, second, and third animal, respectively, using T75 flasks precoated with RetroNectin (TaKaRaBio). Post-transduction expression of huCD25 was analyzed by flow cytometry (BD FACSCalibur). Autologous transduced cells were infused back into the central venous circulation of each animal following myeloablation by irradiation (10 Gy split over 2 days using a Cobalt-60 irradiator [Theratronics]). Cells were resuspended in phosphate-buffered saline and infused as a single bolus with a delivery rate of 2 ml/min. Post-transplantation care included antibiotics, antifungals, antivirals, and fluids (see Supplementary Table S1 for details; Supplementary Data are available online at
DNA purification and qualitative PCR
Genomic DNA was extracted from PB and BM cells using the Gentra Puregene Blood Kit (Qiagen) according to the manufacturer's instructions. PCR was carried out using a Mastercycler PCR system (Eppendorf). The oligonucleotide primers for detection of the vector WPRE sequence were as published (Lizee et al., 2003). Primers for detection of control β-actin sequence (actin-F and actin-R) are listed in Supplementary Table S2 as are PCR conditions. The PCR products were analyzed using 4% agarose gels.
Real-time quantitative PCR and linear amplification-mediated PCR
Quantitative PCR (qPCR) was carried out using a Rotor Gene RG300 system (Corbett Research). For detection of the WPRE and β-actin sequences, primers were as above. Sequence of the probe was as published (Lizee et al., 2003). PCR conditions are listed in Table S2. For each PCR, a no-template reaction was included as a negative control. Each DNA sample (300 ng total genomic DNA) was tested in triplicate, and mean values are reported. Serial dilutions (five dilution points) of DNA from a 293T cell line containing one provirus (established in our laboratory and confirmed by Southern blot analysis) added into DNA from a nontransduced 293T cell line were prepared to create standard curves. The limit of detection was 324 vector positive cells (VPCs)/million 293T cells (data not shown).
Linear amplification-mediated (LAM)-PCR was performed to analyze LV integration sites (Ailles et al., 2002). The protocol and sequences of the primers (LTR2, LC1, LTR3, LC2) are listed in Table S2. The final PCR products were separated on polyacrylamide and/or agarose gels. For identification of LV integration sites, DNA sequencing analysis was performed on the amplicons isolated from agarose gels by the Gel Extraction Kit (Qiagen). Blast searches were done against the NCBI and Baylor databases.
Measurement of AC-specific activity and ceramide levels
AC activity was measured by a fluorimetric HPLC-based assay modified from He et al. (1999). Briefly, cell samples were suspended in 50 μl of sucrose and lysed by repeated freeze–thaw. Total protein concentration was measured using a commercial Lowry protein assay (BioRad). Sample lysate aliquots containing 10 μg of total protein (in a total volume of 20 μl of 0.25 M sucrose) were mixed with 20 μl of the reaction buffer (0.2 M phosphate/citrate buffer pH 4.5, 0.5% sodium taurocholate, 0.5% Triton X-100, and 0.1% bovine serum albumin) containing 200 μM of NBD-C12-Ceramide (Matreya) substrate, and the mixture was incubated at 37°C for 16 hr. The reaction was stopped by adding 380 μl of ethanol and centrifuged at 10,000 × g for 10 min. Ten microliters of reaction supernatant was resolved on a Symmetry C18 column (Waters) with methanol:water (95:5) operated at a flow of 1 ml/min on a Waters HPLC instrument. The fluorescent NBD-C12 product was detected at 530 nm following excitation at 430 nm and was quantified based on known standards. NBD-C12 retention time was 2.4 min. The activity of AC in the samples was determined from the NBD-C12 standard curve and represented in picomoles of the product per milligram of total protein in the injected sample per hour (pmol/mg/hr) following subtraction of the blank. Each sample was analyzed in quadruplicate at least.
Ceramide concentration in cell lysates was determined using Escherichia coli diacylglycerol kinase as previously reported (Bielawska et al., 2001). The E. coli strain was kindly provided by Drs. D. K. Perry and Y. A. Hannun (Medical University of South Carolina, Charleston, SC). Determinations were performed in duplicate at least.
In situ hybridization analyses
For in situ hybridization analyses, procedures are listed in Table S2 as are the forward and reverse primers (WPRE-F-DIG, WPRE-R-DIG) for PCR-based DIG labeling. Counter-staining tissue sections were detected by inverted optical microscope at the UHN Advanced Optical Microscopy Facility.
Other safety parameters
Biopsies of multiple organs including liver and spleen were taken pre- and post-transplantation at different time points (various weeks pre-infusion, and at 3 months and 6 months post-infusion, for spleen and liver) and sent to the Ontario Veterinary College, University of Guelph, for evaluation by a pathobiologist. In addition, chest X-rays were taken pre- and post-irradiation to evaluate any fibrosis of the lungs. General immunological recovery was checked later on by testing for a positive eyelid reaction after injection of tuberculin antigens. General biochemistry parameters and CBCs were measured on blood samples from each animal by Toronto Medical Laboratories (UHN).
Statistics
Data are presented as mean results ± standard error of the mean (SEM) unless specified otherwise. Critical p-values were determined by Student's t-test and p < 0.05 was considered significant.
Results
Transduction of mobilized PB cells and transplantation
Each leukapheresis procedure yielded approximately 3 × 109 to 5 × 109 cells per mobilization cycle; two leukapheresis cycles were performed for each animal. For transductions, the entire mobilized PB mononuclear cell population was thawed and prestimulated overnight prior to incubation with the LV preparation. Post-thaw viability of cells was approximately 80%. Seventy percent to 76% of the cells in culture were found to be huCD25+ as measured 24–48 hr post-transduction by flow cytometry. Variable numbers of cells (see below) were infused into each animal due to differences in mobilization, transductions, and cell recoveries.
Engraftment kinetics and biochemical analyses
After infusion, blood leukocyte counts diminished and then rebounded to reach 1000 cells/μl at an average of 21 days (Fig. 1A). Recovery from neutropenia was more rapid in the third animal likely because we transplanted 14.4 × 106 cells/kg as compared to 3.1 × 106 and 0.4 × 106 cells/kg for animals 1 and 2, respectively. Animal 1 received a whole blood transfusion on day 6 and an infusion of platelets on day 8 and 11. Platelets were recovered 25, 34, and 21 days after transplantation for animals 1, 2, and 3, respectively (Fig. 1B). Hemoglobin recovery was delayed in the first animal (until 106 days), while occurring at approximately 45 days in the other two animals (Fig. 1C). Biochemical analyses (creatinine, urea, protein, albumin, bicarbonate, aspartate aminotransferase, alanine aminotransferase, and alklaine phosphatase) were performed on peripheral blood samples from each animal (Supplementary Fig. S1). Transient changes in some biochemical parameters after transplantation were observed in some animals that subsequently returned to normal levels.
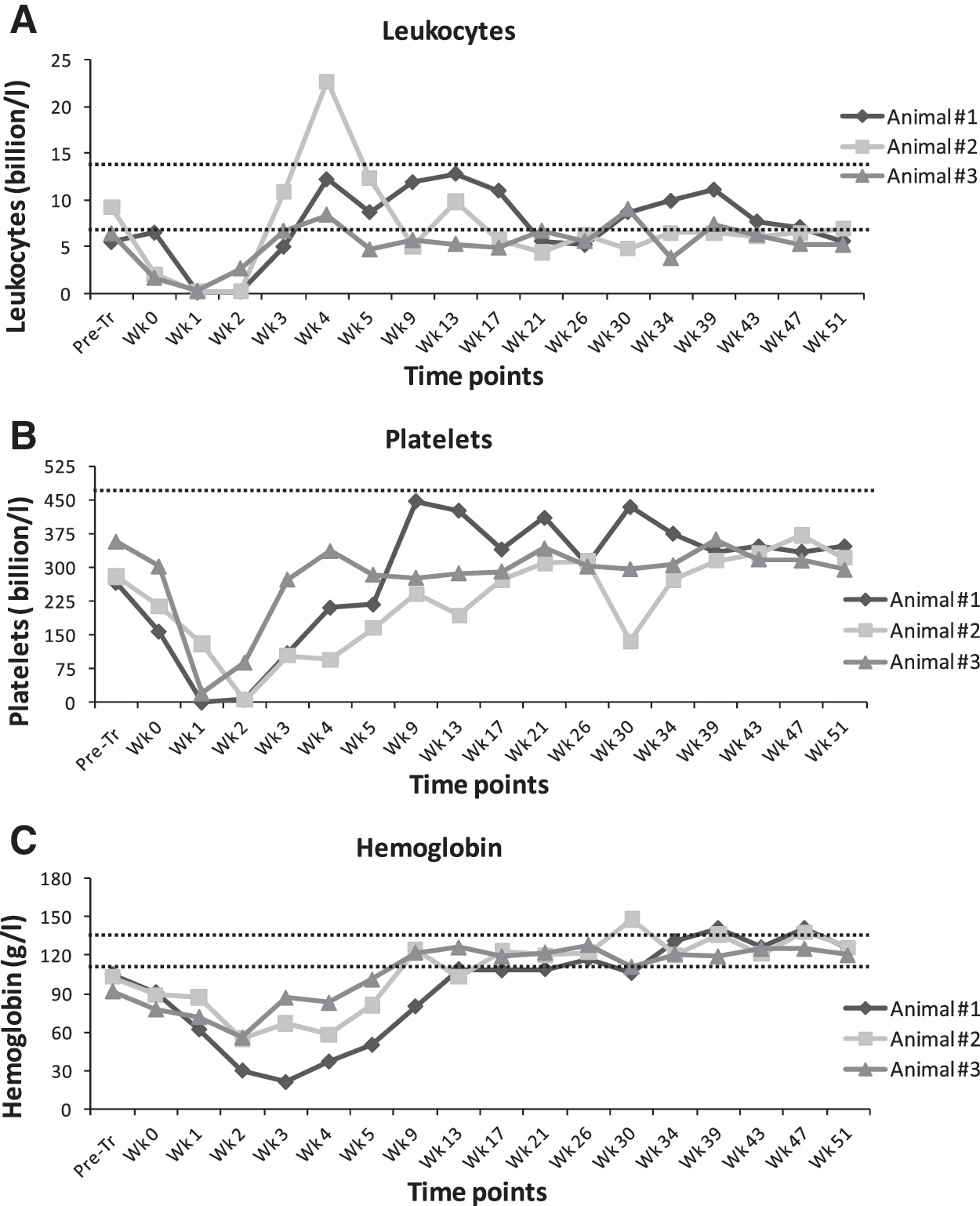
Hematological recovery kinetics for animals 1, 2, and 3. Hematological recovery of
Persistence and quantitation of vector in PB and BM cells
The WPRE present in our LV was used as a marker sequence for qualitative genomic PCR and quantitative real-time qPCR analyses. The qualitative PCR data in Fig. S2 show that the vector sequence could be detected until the endpoint of the experiment (i.e., 1 year) in both PB and BM cells from all three animals. Next, we developed a sensitive qPCR assay to estimate the number of vector positive cells (VPCs)/million in PB and BM cells obtained from the transplanted animals (Fig. 2). Figure 2A shows there were up to 2.5 × 105 VPCs/million PB cells in two of the animals at 1 month. This level of marking of PB cells decreased over time (for example down to 0.1 × 105 VPCs/million at 11 months in the third animal) except in animal 1. For BM, analyses of samples from animals 1, 2, and 3 again showed higher initial marking values that subsequently decreased over time, except in animal 1 (Fig. 2B). In animal 2, values decreased to a level less than our quantitation limit for this assay at 14 months. In animal 3, some vector positive cells above background levels were still detected in BM at 11 months (Fig. 2B).

Determination of integration events in all three animals by real-time quantitative PCR (qPCR). The WPRE sequence in the lentiviral vector (LV) was used to estimate the number of vector positive cells/million in
Molecular analyses and other safety parameters
As can be seen from the LAM-PCR analyses (Fig. S3), multiple marked hematopoietic cell clones exist in BM and PB samples from all three animals. Sequencing of several predominant bands following extraction confirmed unique integration sites for each recovered band (Supplementary Table S3). In situ hybridizations were performed on sections generated from biopsies at 28 weeks post-transplantation of organ tissues (heart, liver, and spleen) from animal 3. No WPRE-positive staining was observed in the heart (Fig. 3). Positive staining was observed in the spleen and in cells with a distinct hematological morphology within blood vessels of the liver (Fig. 3).
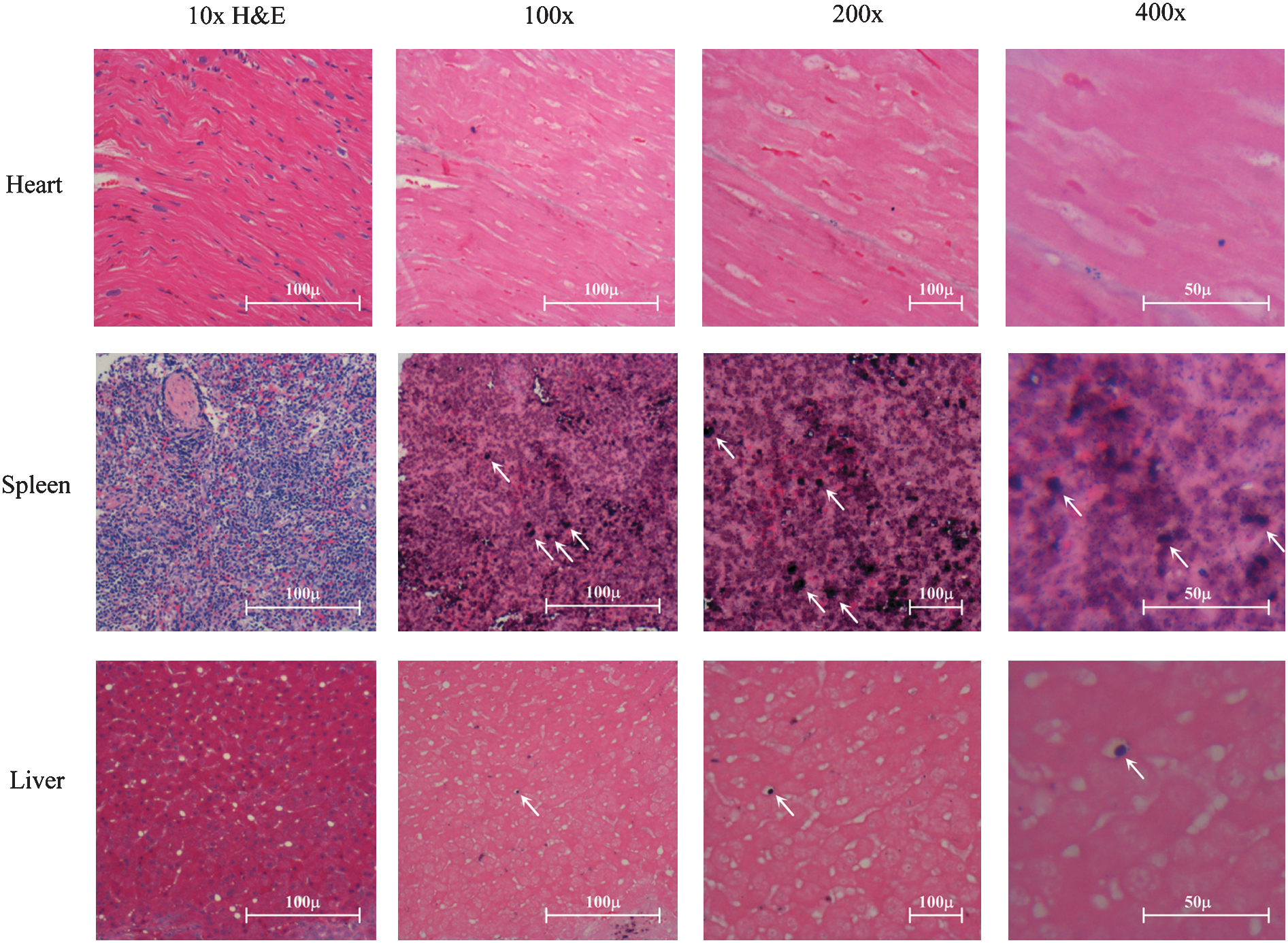
In situ hybridization analysis of LV presence in liver, spleen, and heart tissue sections of animal 3. Cross-sections of rhesus heart, spleen, and liver tissues of animal 3 obtained at biopsy at 28 weeks were either developed with hematoxylin and eosin (H&E) stain (column 1) or used for in situ hybridization with a LV-specific primer (columns 2–4). Select vector-positive cells are shown by arrows where indicated.
Note that we did not find any radiological changes in lung fields (fibrosis/mass) when we compared pre- and post-transplantation X-ray films of all animals. All three animals also showed general immunocompetence as documented by a positive eyelid reaction after BCG vaccination, which we performed 1 month prior to sacrifice. There were no significant pathological changes observed in organs of any animal as determined by histopathological assessments of samples taken pretransplantation, and between 3 to 7 months post-transplantation, other than mild interstitial nephropathy in animals 1 and 2, which is a common background lesion in adult rhesus macaques and could also be a consequence of total body irradiation (Bielefeldt-Ohmann et al., 2004). A number of instances of mild focal arteritis post-transplantation were noted in the spleen and testes of animals 1 and 2, respectively, which is not uncommon given that vasculitis in various organs has been reported before in subjects undergoing irradiation (Bielefeldt-Ohmann et al., 2004).
AC activity and ceramide levels in PB and BM cells
AC-specific activity was determined in PB and BM samples from animals 2 and 3 by a fluorimetric HPLC-based assay that we developed (insufficient samples were collected from animal 1 for this assay). Significant increases of up to two-fold over pretransplantation levels were observed in BM cells from animal 2 at 17, 50, and 67 weeks posttransplantation (Fig. 4A). Likewise, AC activity in BM and PB samples of animal 3 increased up to two-fold over background levels within the first 6 months post-transplantation of transduced cells (Fig. 4B). AC activity in a BM sample collected at 30 weeks post-transplantation from animal 3 showed a decline from those levels but still remained above pretransplantation levels. Meanwhile, PB samples from the same animal showed a supranormal degree of AC activity that remained high until week 26, which was the last sample measured.

Evaluation of acid ceramidase (AC)-specific activity and ceramide levels in peripheral blood (PB), bone marrow (BM), spleen, liver, and brain samples. AC-specific activity in select samples including pretransplantation values were measured from
Ceramide levels were measured in PB cells from animal 3 (Fig. 4C). Three weeks after cell infusion, ceramide content declined by more than 40% from pretransplantation levels (Fig. 4C). Furthermore, even after 30 weeks total ceramide levels were still below pretransplantation values (Fig. 4C). Ceramide to DAG ratios were also determined in spleen and liver samples from animals 1 and 2 (insufficient samples were available from animal 3). Ceramide and DAG are two very similar lipids, both structurally and functionally. These two lipids were simultaneously quantified using an enzyme (the bacterial DAG kinase) that phosphorylates both, and 32P-labeled derivatives were subsequently quantified. This method provides a way to quantify ceramide without the variances inherent in total organ/tissue protein quantification by normalizing the values to the content of a similar lipid that is not subject to such variances in sampling. Comparisons of pretransplantation ratios with end-point ratios demonstrated dramatic declines over time (Fig. 4D). Next, to establish a baseline ratio of ceramide to DAG levels in the context of naturally occurring variations in these macaques, PB cell samples were obtained from 4 independent nonstudy macaques housed within our facility and combined with pretransplantation values obtained from animal 3. We then compared that ratio against a sample from animal 3 obtained at 30 weeks post-transplantation. Here the ceramide/DAG ratio was again found to be significantly reduced (Fig. 4D).
We also assayed AC-specific activity on organ samples obtained at sacrifice from animals 1–3 in comparison to organ samples from nine independent control animals. Significant increases in AC-specific activity were observed in spleen and liver (Fig. 4E). Note that the fidelity of the liver sample from animal 2 was poor and demonstrated variable (and high) protein concentration compared with the other samples. No significant changes were detected in the brains of animals receiving transduced cells compared to controls. To further examine the brain given the relevance of this organ in Farber disease, we also assayed for long-term ceramide reduction and compared results against those obtained from the control animals (Fig. 4F). No significant differences were seen although there were trends towards decreased ceramide in the test animals.
Discussion
Most LSDs lack definitive treatments. Bone marrow transplantation (BMT) has been implemented in a few trials as the sole treatment modality for Farber disease; partial resolution of peripheral manifestations has occurred (Vormoor et al., 2004; Ehlert et al., 2006). We hypothesize that while BMT provides infusion of metabolically normal cells, those cells are not equipped to provide additional AC activity beyond basal parameters. Indeed, for a number of LSDs, engineered overexpression of such lysosomal hydrolases in cells from the hematopoietic system, for example, has led to diffusion of enzyme activity and then to distant metabolic cooperativity effects. Leukocytes themselves also appear to be dysregulated in Farber disease. Further, hematopoietic cells have also been demonstrated to have the capacity to migrate through the blood–brain barrier and take up residence in the brain; evidence from other LSDs indicates that BMT using normal cells can lead to some neurological improvements (Whitley et al., 1993; Peters et al., 1996).
For these reasons and the facts that hematopoietic cells can be readily collected and transduced ex vivo, we are developing gene therapy for Farber disease targeting this population. We employ recombinant LVs due to the efficient and integrative nature of this delivery schema along with the favorable safety profile (Neschadim et al., 2007). As mentioned, we have previously demonstrated the feasibility of gene therapy for Farber disease in cell culture and mice. For this present study we sought to extend these findings into a more clinically relevant model. Indeed, we chose macaques due to their size (closer to our target pediatric patient population) and their genetic relatedness to humans. Studies have also shown long-term persistence of retroviral vector-transduced cells in this large-animal model (An et al., 2000, 2001; Kiem et al., 2004). Furthermore, we chose to target mobilized PB cells in this study without enrichment for more primitive populations. Selection would have impacted our yield; we reasoned that infusion of higher numbers of LV-transduced cells comprised of this broad and developmentally varied population would provide an immediate spike in AC activity that could reduce substrate effectively in the short term. Furthermore, transduction of this entire population might provide for more long-term benefits from correction, albeit at a lower frequency, of more primitive repopulating cells.
We examined a number of parameters in this study. We observed no permanent deleterious consequences from any aspect of the protocol in any animal. None of the proviral integration sites sequenced disrupted coding sequences of known genes. We also observed long-term persistence of LV-transduced cells. We found that a relatively low percentage of transduced cells as measured by one sensitive molecular assay could generate detectable AC activity levels above background and lead to ceramide reduction even in these enzymatically normal hosts. This mirrors results we have observed in normal mice concerning gene therapy for Fabry disease (Takenaka et al., 1999). To reiterate, given that all the animals already expressed wild-type levels of AC activity pretransplantation and that some modest percentage of PB/BM cells were found to be LV-positive over time, additional enzyme increases above background is a highly significant finding. While encouraging, however, observed percentages of transduced cells decreased over time—except in animal 1. This could be due to the possibility that a true stem cell was not infected in animals 2 and 3 followed by subsequent death of transduced short-term progenitor cells with limited self-renewal capacity. Some cell clearance may have also resulted from an ensuing immune reaction against the transduced cells since we used human transgenes engineering expression of proteins not native to macaques. We also transduced whole BM cells, which contain more mature antigen-presenting cells that can possibly present peptides from both huAC and huCD25 as foreign. Alternately, because stem cells cycle, perhaps two of the three animals demonstrated a switch from one source (transduced) of hematopoietic cells to another (nontransduced). Given our significantly enhanced spleen and liver enzyme activity levels at the end-point of the study, however, it may also be possible that our BM PCR data is simply a result of a negative sampling bias because samples were collected from only one site and engraftment may not have been uniform throughout the host. In that scenario it is then likely that a reservoir of highly-expressing transduced cells engrafted elsewhere in order to continually supply systemic AC activity in trans.
In future studies, enrichment of more primitive populations will allow concentration of LV stocks on hematopoietic stem cells with an enhanced capacity for repopulation. This should lead to increased percentages of marked PB cells over time as these cells self-renew and differentiate. This may also lead to better immunological tolerance by reducing the frequency of infected mature antigen-presenting cells. It may also facilitate brain remodeling by transduced cells because we did not observe detectable differences in AC activity or ceramide/DAG ratios in that organ in our present study. We plan to instill transduced cells into recipients by direct intrafemoral injection as well (Mazurier et al., 2003). Our own studies have also shown that reduced conditioning regimens may be invoked and result in establishment of stable hematopoietic cell chimeras that can lead to systemic LSD correction (Liang et al., 2007). We also plan to incorporate a novel safety element we recently described (Sato et al., 2007) into future therapeutic vectors generated to allow for cell fate control should deleterious outcomes occur. By extension, such a safety component will also open the door to broadening the feasibility of allogeneic transplantation for Farber disease if matched donors cannot be found or if autologous cell recovery is limiting or otherwise compromised.
In conclusion, transplantation of autologous LV-transduced hematopoietic cells is a safe and effective therapy. This strategy can be utilized for the treatment of monogenic defects like Farber disease and other LSDs; this work in this large animal model thus opens a pathway to clinical gene therapy for the treatment of such inherited metabolic disorders.
Footnotes
Acknowledgments
We acknowledge Vanessa Rasaiah, Kenneth Zhang, and Dr. Takahiro Nonaka for managerial and technical support; Dr. Robert Donahue from National Heart, Lung, and Blood Institute and Dr. Robert Andrews, Fred Hutchinson Cancer Research Center (FHRCH), for guidance and discussions; Dr. Carmen Bedia from Institut National de la Santé et de la Recherche Médicale U858, Toulouse for some enzyme measurements; Mui Chua and Pamela Harmon from Princess Margaret Hospital (PMH) for guidance with the leukapheresis procedure; Dr. Badru Moloo and Sandra Lafrance from Animal Resources Centre, UHN, for their assistance; and Dr. Patricia V. Turner from the Laboratory Animal Pathology, University of Guelph, for histopathological reports. Funding for A.N. was provided by the Canadian Institutes of Health Research Training Program in Regenerative Medicine. This work was supported by grant 5R21NS051500 from the NIH to J.A. Medin and a grant from the Association Vaincre les Maladies Lysosomales to T.L. and J.A. Medin.
Conflict of Interest Statement
The authors have no conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.