
Abstract
Gene electrotransfer is expanding in clinical use, thus we have searched for an emergency procedure to stop transgene expression in case of serious adverse events. Calcium is cytotoxic at high intracellular levels, so we tested effects of calcium electrotransfer on transgene expression in muscle. A clinical grade calcium solution (20 μl, 168 mM) was injected into transfected mouse or rat tibialis cranialis muscle. Ca2+ uptake was quantified using calcium 45 (45Ca), and voltage and time between injection and pulsation were varied. Extinction of transgene expression was investigated by using both in vivo imaging of infrared fluorescent “Katushka” and erythropoietin evaluated by ELISA and hemoglobin. Histology was performed. Electrotransfer of Katushka and erythropoietin yielded significant expression. Maximal calcium uptake occurred after injection of Ca2+ before electropulsing using eight high voltage pulses of 1000 V/cm. Using these parameters, in vivo imaging showed that transgene expression significantly decreased 4 hr after Ca2+ electrotransfer and was eliminated within 24 hr. Similarly, serum erythropoietin was reduced by 46% at 4 hr and to control levels at 2 days. Histological analyses showed muscle damage and subsequent regeneration. Electrotransfer of isotonic CaCl2 terminates transgenic protein expression in muscles and may be used for contingency elimination of transgene expression.
Introduction
One efficient way of transfecting genes into muscle tissue is DNA electrotransfer (Aihara and Miyazaki, 1998; Mathiesen, 1999; Mir et al., 1999). This method utilizes short electric pulses to transiently permeabilize cell membranes, rendering the cells accessible for plasmid DNA. This method distinguishes itself from other transfection methods like viral or liposomal therapies in that the transfection only takes place in the tissue encompassed by the electrodes delivering the electric field (Gehl, 2003). Thus the transfected area can be precisely localized following the treatment for any post-transfection intervention. Great efforts have been put into optimizing DNA electrotransfer so that the method itself causes as little disturbance to the muscle tissue as possible (Hojman et al., 2007c, 2008), and low level of tissue damage is induced when DNA electrotransfer is performed accordingly.
Therapeutic use of DNA electrotransfer to muscle tissue involves vaccination purposes, systemic delivery of therapeutic proteins, and local intramuscular expression. The clinical advances have so far been made within DNA vaccines to muscle tissue with seven on-going clinical trials (Heller and Heller, 2006; Hojman, 2010), but muscle electrotransfer also holds great clinical promise for systemic delivery of therapeutic proteins, e.g., for protein deficiency syndromes and cancer treatment. Muscle tissue has a large capacity to produce and secrete protein and may function as a protein factory for systemic delivery (Lu et al., 2003). DNA electrotransfer to muscle tissue has proven to be highly efficient, resulting in systemic therapeutic levels of the transgenic product (Aihara and Miyazaki, 1998; Bettan et al., 2000; Lucas and Heller, 2001; Draghia-Akli et al., 2003; Fattori et al., 2005; Hojman et al., 2007b). Thus, a small volume of muscle tissue may be used to obtain systemic concentrations of protein.
We have speculated that if systemically therapeutic levels of transgene expression can be achieved by electrotransfer to just a small area of muscle (Hojman et al., 2007b), then it would be possible to extinguish this transgene expression by eliminating this small area of transfected muscle tissue. We have previously tried injections of cytotoxic compounds like 100% ethanol, but this was not successful in eliminating or even reducing the transgene expression (data not published). Others have suggested injections of bupivicaine, which is known to induce muscle necrosis and regeneration in the injected area with alterations in the gene expression pattern in the regenerated muscle (Clark et al., 1997). After pulse administration muscle cells are in a permeabilized state for a matter of minutes (Bier et al., 1999; Gehl et al., 2002). During this time, exchange of electrolytes across the plasma membrane takes place (Gissel, 2010; Hojman et al., 2008); notably, there is an inward flux of Na+ and Ca2+ and an outward flux of K+. After membrane resealing, homeostasis is restored. Since a high concentration of Ca2+ has a cytotoxic effect, it may be possible to use electrotransfer of Ca2+ as a method to eliminate the transgene expression, and this can serve as a simple contingency solution in case of unwanted side effects of the gene therapy.
Materials and Methods
Animals and muscle preparation
All animal experiments were conducted in accordance with the recommendations of the European Convention for the Protection of Vertebrate Animals used for Experimentation and with permission from the Danish Animal Experiments Inspectorate. Animal experiments were performed on 8- to 10-week-old female C57Black/C or NMRI mice from Taconic or 4- to 8-week-old Wistar rats. The animals were maintained in a thermo-stated environment under a 12-hr light/dark cycle with free access to food and drinking water.
Plasmid constructs and in vivo DNA electrotransfer
The plasmids encoding genes of interest included pHD-mEPO encoding murine EPO under the control of an rtTA-dependent promoter (Hojman et al., 2007b) and pCMV-Kat encoding the far-red fluorescent protein Katushka (Kat) (Evrogen). The plasmids pTet-On, encoding the rtTA transactivator, and pTetS, encoding the tS silencer, were both obtained from Clontech. All DNA preparations were performed using Qiafilter Plasmid Maxiprep kits (Qiagen), and the concentration and quality of the plasmid preparations were verified by spectrophotometry and gel electrophoresis.
The animals were anesthetized 15 min prior to DNA electrotransfer by intraperitoneal injection of Hypnorm (0.4 ml/kg, Janssen Saunderton) and Dormicum (2 mg/kg, Roche). The plasmid solution (1.5 μg in mice and 2 μg in rats in 20 μl) was injected intramuscularly along the fibers into the tibialis cranialis muscle using a 29-gauge insulin syringe. Plate electrodes with a 4-mm gap in mice and a 6-mm gap in rats were fitted around the hind legs. Good contact between the electrode and skin was ensured by hair removal and use of electrode gel. DNA electrotransfer was performed using a combination of a high voltage (HV; 800 V/cm [applied voltage = 320 V in mice, 480 V in rats], 100 μsec) pulse followed by a low voltage (LV; 100 V/cm [applied voltage = 40 V in mice, 60 V in rats], 400 msec) pulse (Cliniporator) with volts per centimeter being the voltage to distance between the two plate electrodes. Induction of gene expression was obtained by administering drinking water containing doxycycline (doxycycline hyclate, Sigma-Aldrich) at 0.2 mg/ml in distilled water (Hojman et al., 2007a).
Ca2+ electrotransfer
Ca2+ injection solution (9 mg/ml, 225 mM, Sandox) approved for clinical use was diluted to 168 mM with sterile water, and 20–30 μl was injected into the tibialis cranialis. Electrotransfer was performed as described in the previous section by using eight pulses with a duration of 100 μsec, at 1000 V/cm (applied voltage 400 V in mice, 600 V in rats), delivered at a frequency of 1 Hz.
Calcium 45 electrotransfer
Calcium 45 (45Ca) was added to the 168 mM Ca2+ solution, resulting in a final activity of 0.5 μCi/ml. Twenty microliters was injected before, after, or both before and after (thus 40 μl in total) placement of electrodes and pulse application (Ca2+ + EP). Electrotransfer was performed as described by using eight pulses of 100-μsec duration, delivered at a frequency of 1 Hz at increasing pulse amplitudes. At the times given the animals were killed and the tibialis cranialis muscle was excised, weighed, and soaked overnight in 2.5 ml of 0.3 M trichloroacetic acid (TCA). 45Ca activity of the TCA extract was determined by liquid scintillation counting (Tri-carb2100 TR, Packard). Results are expressed as counts per minutes per gram of tissue (CPM/g wet wt.).
Blood analyses
In mice, blood samples (20 μl) were drawn from the retro-orbital sinus and hemoglobin levels were measured from a drop of blood using the HemoCue Hb201 + (HemoCue AB). At the termination of the experiments, 500 μl of blood was drawn from the mice by cardiac puncture and processed to serum. In rats, blood samples (200 μl) were drawn from the tail vein at the indicated time points. A drop was used for determination of hemoglobin and the rest was processed to serum. A Mouse/Rat Erythropoietin ELISA kit (RnD Systems) was used to determine erythropoietin (EPO) serum levels.
In vivo bio-imaging
Mice were anesthetized and placed in a custom-made bed, which allowed stable and reproducible imaging of the legs. In vivo scanning was performed using the Optix MX-2 Optical Molecular Image System (Advanced Research Technologies), which uses time domain optical imaging, allowing for quantitative determination of emission in the region specific to the Kat protein. Kat excitation was performed with a 635-nm (LDH-P-635) pulsing laser and emission was detected with a 650 long pass filter. The scan was performed over a Cartesian grid in prioritized raster fashion with 1-mm resolution, and each scan took 5 min on average. Data analysis was performed using the Optiview 2.2 software performing background subtraction, intensity measurements, and lifetime analysis of the fluorochromes. Lifetime values are reported as means of lifetimes in the gated transfected areas.
Histology
Twenty-four hours or 2 weeks after Ca2+ electrotransfer, muscles were isolated and fixed in 1 ml formalin buffer. After paraffin embedding, transverse sections of 2 μm were prepared and stained with hematoxylin and eosin (HE).
Statistics
Two-way ANOVA with ad hoc Bonferroni post-tests was performed to determine significant differences between treatment groups and time points, and p < 0.05 was considered statistically significant.
Results
45Ca uptake in muscles after electrotransfer
To determine the conditions for maximal Ca2+ influx with combinations of Ca2+ injection (20 μl of 168 mM CaCl2) and electroporation, we tested 45Ca uptake in mouse muscle injected with Ca2+ before, after, or both before and after electroporation. Injection of Ca2+ before electroporation increased 45Ca uptake sevenfold (p < 0.001), while injection of 45Ca after electroporation had smaller effect with a 2.3-fold increase in Ca2+ influx (p < 0.01). Injecting 45Ca both before and after electroporation did not induce additional increases in the 45Ca uptake compared to injection before electroporation, even though the received the double amount of Ca2+ (Fig. 1A).
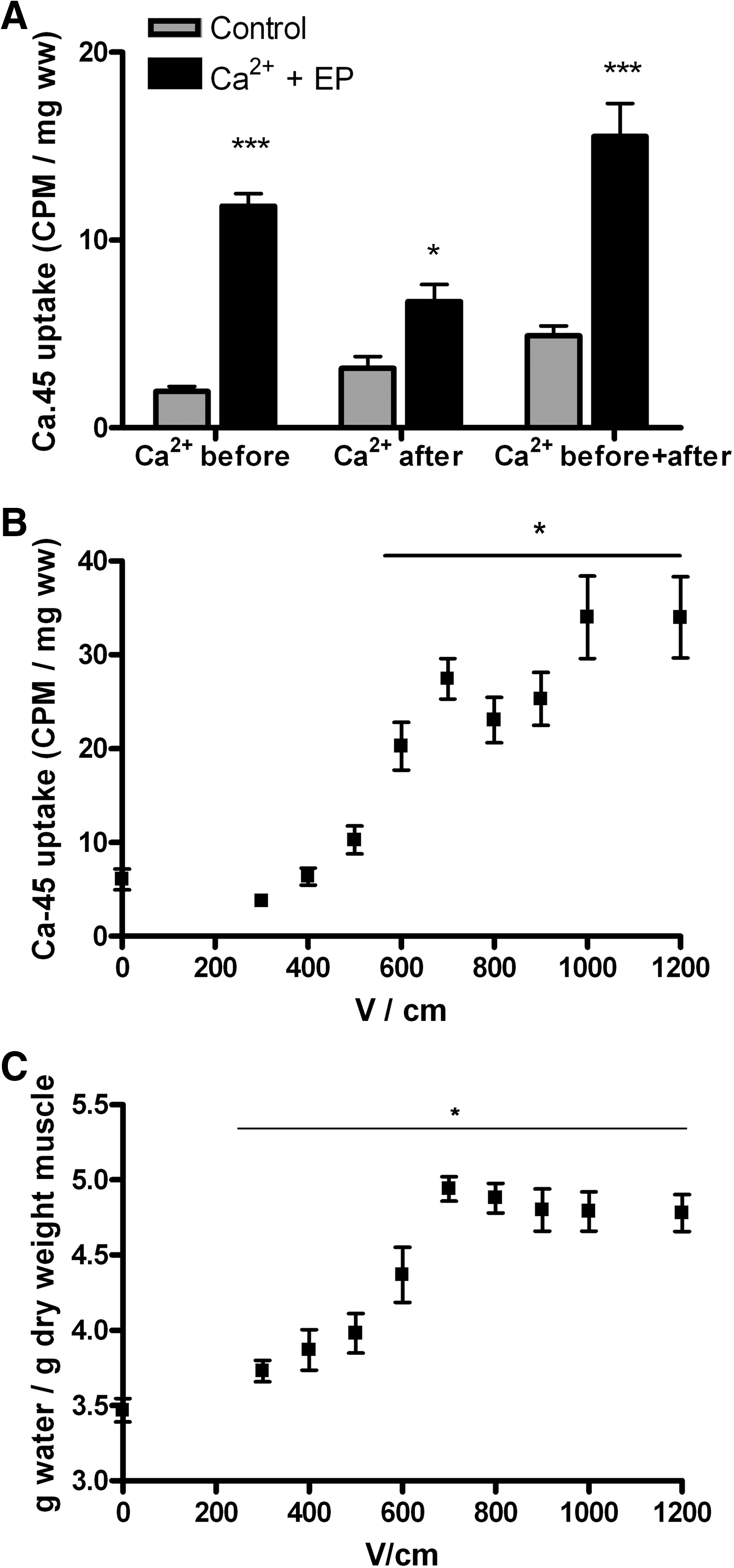
Calcium influx in muscles during electroporation (EP). Combinations of Ca2+ injection (20 μl of 168 mM CaCl2 with 0.5 μCi 45Ca) and EP were tested.
We tested the voltage dependency of Ca2+ uptake and found a significant increase in Ca2+ uptake at pulse amplitudes above 500 V/cm, reaching a plateau in Ca2+ uptake at 1000 V/cm (Fig. 1B). In these muscles, we also determined the water content and found that this increased from pulse amplitudes above 300 V/cm and reached a plateau at 700 V/cm (Fig. 1C). Based on these results, we decided to use eight pulses (100 μsec) of 1000 V/cm with Ca2+ injection before the application of the electric pulses for Ca2+ electrotransfer.
Termination of Kat expression
To evaluate if Ca2+ electrotransfer could terminate transgene expression, muscles from mice were transfected with the far-red fluorescent molecule Kat and fluorescence was followed by in vivo imaging. Four days after Kat electrotransfer, all muscles had a clear Kat signal with mean intensity of 3305 ± 464 NC and mean lifetime of 2.04 ± 0.01 nsec (N = 25). The muscles were then treated with Ca2+ electrotransfer, Ca2+ injection only, electroporation only, or no treatment (control) (Figs. 2 and 3). In the Ca2+ electrotransfer–treated muscles, fluorescence intensity dropped significantly (p < 0.05) already 4 hr after treatment and remained significantly reduced at the level of untransfected muscles from 24 hr and at the following time points (Fig. 3). The half-life of Kat in aqueous solutions is 20 min (Shcherbo et al., 2007), thus our results indicate that expression of Kat is immediately turned off at the time of Ca2+ electrotransfer. There was no significant difference between the electroporated, Ca2+ injected, and control group, and all three groups peaked in fluorescence intensity 72 hr after Ca2+ electrotransfer (thus 7 days after Kat electrotransfer). This is consistent with previous reports, showing that Kat expression peaks 1 week after Kat electrotransfer (Hojman et al., 2009).

In vivo imaging of Katushka (Kat) expression following Ca2+ electrotransfer. Time-domain in vivo imaging of Kat expression in muscles prior to and 4, 24, 48, 72, and 168 hr after Ca2+ electrotransfer. Four days prior to Ca2+ electrotransfer all muscles were transfected with Kat. The pictures depicted a representative mouse (same mouse at all six time points) for the four conditions: Kat (untreated); Ca2+ (20 μl of Ca2+, 168 mM); EP (eight high voltage [HV] pulses of 1000 V/cm); and Ca-EP (Ca2+ electrotransfer with 20 μl of Ca2+, 168 mM) followed by eight HV pulses of 1000 V/cm). Each group consisted of eight mice.

Fluorescence intensity and lifetime of Kat expression. Fluorescent signals from Fig. 2 were analyzed for
To determine the stability of the fluorescence signal, lifetime, which is an intrinsic property of the fluorochrome, was measured. For Kat, the lifetime is 2.1 nsec. In the Ca2+ electrotransfer treated muscles, the lifetime decreased dramatically, and was significantly reduced 24 hours after Ca2+ electrotransfer and at the following time points (p < 0.001) (Fig. 3B), indicating that the integrity of the Kat molecule is decreasing. At 48 hr, the lifetime had reached the level measured in untransfected control muscles (data not shown), indicating that the Kat expression was completely eliminated. No reductions in the lifetime signal in the electroporated, Ca2+ injected, or control groups were observed, consistent with the finding that Kat expression remains stable in these muscles.
Termination of EPO expression
In addition to the termination of a locally expressed transgene, it was investigated if Ca2+ electrotransfer could also terminate expression of systemically secreted transgenic proteins, such as EPO. EPO electrotransfer with 2 μg to one rat tibialis cranialis muscle resulted in significant increases in serum EPO (EPO: 460.4 ± 123.5 pg/ml, control: 27.5 ± 2.2 pg/ml, n = 7–14, p < 0.05) and hemoglobin levels (EPO: 9.6 ± 0.6 mmol/L, control: 7.4 ± 0.3 mmol/L, n = 7–14, p < 0.05) as previously described (Hojman et al., 2007b). Fourteen days after EPO electrotransfer, half of the EPO-transfected muscles were treated with Ca2+ electrotransfer, while the rest remained untreated. As depicted in Fig. 4A, the serum EPO levels decreased rapidly and were reduced 46% within 4 hr of termination compared with animals not receiving Ca2+ electrotransfer (p < 0.05). The half-life of serum EPO is 3–7 hr (Gothelf et al., 2010). After 48 hr, the serum EPO levels in the Ca2+ electrotransfer–treated animals (37.2 ± 6.3 pg/ml, n = 7) were at the level of control animals (25.0 ± 0.1 pg/ml). In continuation, the hemoglobin levels in the Ca2+ electrotransfer treated rats returned to control levels 2 weeks after Ca2+ electrotransfer (p < 0.001, Fig. 4B). At 4 hr, an initial increase in serum EPO levels was observed for both control groups (control and EPO). This increase is associated with hypoxia during the anesthesia, as has been reported previously (Gothelf et al., 2009).

Termination of muscular erythropoietin (EPO) expression. Blood samples were taken prior to; 4, 24, 48, 72 hr; and 1 and 2 weeks after Ca2+ electrotransfer for the determination of hemoglobin and serum EPO levels.
Similarly, serum EPO levels in mice were determined after Ca2+ electrotransfer, electroporation, Ca2+ injection, or no intervention, respectively. As shown in rats, Ca2+ electrotransfer completely eliminated EPO expression, and serum EPO levels returned to baseline within 2 weeks. Treatment with electroporation or Ca2+ injection did not have any effects on serum EPO and hemoglobin levels in mice (Fig. 4C).
Safety of the procedure
Four hours after Ca2+ electrotransfer, the Ca2+ electrotransfer treated legs were markedly swollen compared to the control legs. A slight swelling of the electroporated legs was also observed. To assess functional impairments, motor and reflex function were tested 24 hr after Ca2+ electrotransfer in the treated muscles as previously described (Hojman et al., 2007c). No impairments in running and grabbing behavior were observed for any of the treatments (data not shown). Testing at 4 hr could not be performed because the mice were still affected by the anesthesia.
Histological analysis was performed on routine 2-μm sections of formalin-fixed, paraffin-embedded mouse muscles, isolated from the mice 24 hr after Ca2+ electrotransfer and stained with hematoxylin-eosin (HE). Evaluation was blinded and the findings showed four patterns (Fig. 5): (1) normal morphology in control muscles, 2) normal muscle morphology with only one focus of coagulation necrosis and mixed inflammation in the Ca2+-injected muscles, 3) diffuse mixed inflammation and diffuse coagulation necrosis in the electroporated muscles, and 4) diffuse mixed inflammation, diffuse coagulation necrosis, and extravasation of red blood cells in the Ca2+ EP-treated muscles.
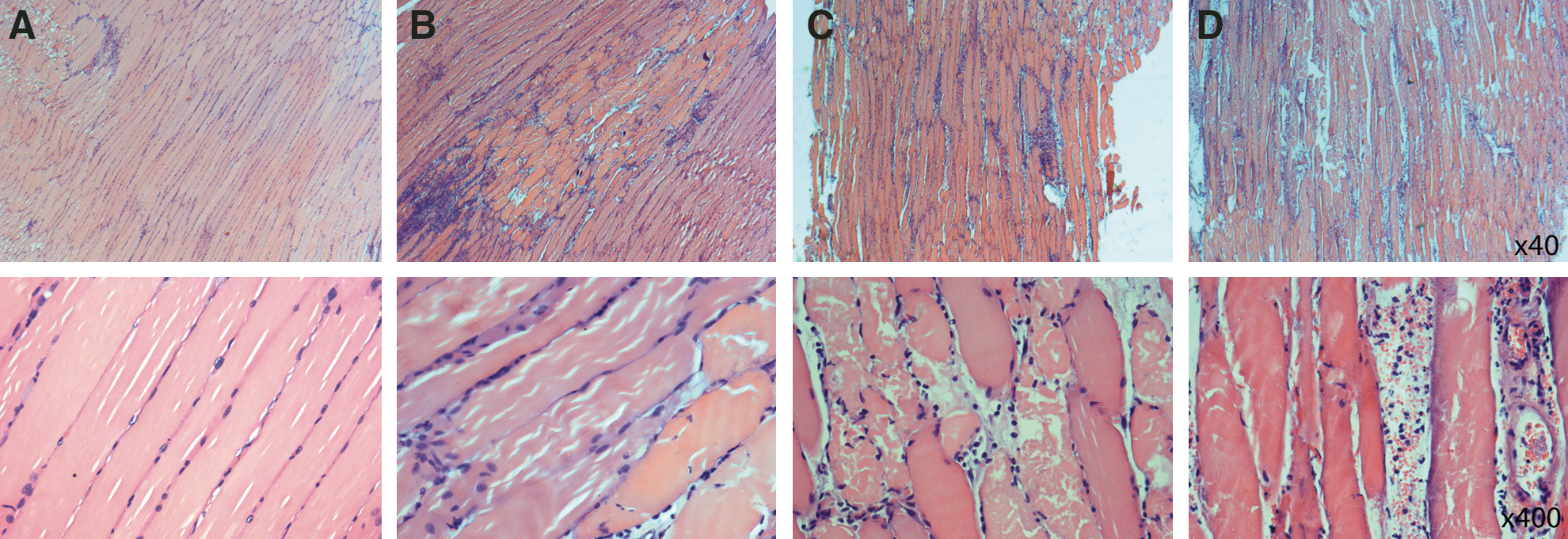
Histological analysis of muscle after Ca2+ electrotransfer. Twenty-four hours post treatment with Ca2+ injection only, EP only, Ca2+ electrotransfer, or no intervention, respectively; muscles were taken for histological examination.
Investigation of mouse muscles 14 days after treatment showed a variety of pathological changes ranging from small to larger foci of necrosis and inflammation. Overall, the changes were more subtle and the main part of the muscle fibers had regained striation. One muscle treated with Ca2+ electrotransfer showed a large focal area with changes of probable necrosis that could not be characterized further. This area was also visible by eye when removing the muscle. The focal effects might be explained by differences in muscle fiber diameter and orientation. Surprisingly, two muscles receiving no treatment showed slightly diffuse inflammation and reactive changes. In contrast, muscles receiving Ca2+ electrotransfer showed clear signs of regeneration.
Discussion
In this study, we show that loading muscles expressing transgenic proteins with Ca2+ completely abolished transgene expression and production. The elimination of transgene expression was already significant 4 hr after Ca2+ electrotransfer and was complete at 48 hr. No impairments in running and grabbing behavior were observed, although major histological changes were seen. At 14 days after treatment, the muscles showed clear signs of recovery including regaining muscle fiber striation. Thus, this method represents a safe and efficient way of terminating transgene expression.
Clinical perspective for Ca2+ electrotransfer
As gene electrotransfer to muscle tissue is increasingly used in clinical trials (Daud et al., 2008; Sallberg et al., 2009; Hojman, 2010), solutions for control of expression have been investigated. A specific issue is having an emergency shut-off strategy available, and precise and quick termination of transgene expression is especially highly warranted. Gene therapy with viral or chemical vectors transfect disperse tissues, thus precise localization of the site of transfection is not possible. In contrast, physical transfection methods like electrotransfer offer precise positioning of the transfected area due to the intrinsic properties of the transfection methods, i.e., transfer only takes place in the tissue encompassed by the electrodes delivering the electric field. Thus in this case, it is possible to relocate the transfected area for post-transfection interventions, e.g., in order to terminate the transgenic signal. Inducible promoter systems have been proposed to control gene expression. Currently no inducible promoter systems have been approved for use in the clinical setting because concerns like the inducer drugs, which in the current systems are either antibiotics or hormone analogues, exist. Furthermore, although extensive efforts have been put into optimizing these inducer systems, leakage in gene expression does still occur, and withdrawal of the exogenous inducer does not therefore necessarily result in complete elimination of the transgene expression. In contrast, we foresee that Ca2+ electrotransfer can be introduced fairly easily into the clinic because both application of electric pulses and Calcium-Sandoz is routinely used in the clinical setting. The set-up for Ca2+ electrotransfer will mimic DNA electrotransfer or electrochemotherapy regimens, and thus would be directly applicable in facilities that are already applying these procedures.
Electroporation has been used extensively in the clinic for electrochemotherapy for cancer patients (Heller et al., 1998; Gothelf et al., 2003; Marty et al., 2006; Mir et al., 2006). Thus the clinical feasibility of electrotransfer-based techniques is well established. In this study, we combined electrotransfer with Ca2+. Calcium is an endogenous ion with little toxic effect; in fact the only foreseeable side effect of this molecule is hypercalcemia. For Ca2+ electrotransfer in rats, we injected 30 μl of 168 mM Ca2+, which equals 5 μmol and thus yields a 27% increase in plasma Ca2+ concentration if all Ca2+ is recovered in the blood. Yet the rats did not show any signs of adverse effects or discomfort. In patients, far lower increases in plasma Ca2+ concentrations are foreseen. Current clinical trials with muscle electrotransfer have treated muscle volumes of a maximum of 1 cm3, with the muscle fibers transversing this area transfected in their full length. In this case, one treatment of Ca2+ electrotransfer (100 μl of 168 mM) will induce a potential 1% increase in plasma Ca2+ concentration.
Pulse parameters for Ca2+ electrotransfer
In this study, we used two different kinds of pulse regimens for DNA and Ca2+ electrotransfer, respectively. Previous studies have shown that electrotransfer of small molecules and plasmid DNA relies on different mechanisms. Small molecules and ions depend on simple diffusion through the permeabilized membrane, and short (μsec) pulses of high amplitude result in the best delivery. Because Ca2+ is an ion and has a very large concentration gradient, we chose this pulse regimen. This mechanism is also explored in the clinical electrochemotherapy protocols of transfer of drug molecules, e.g., bleomycin (Mir et al., 2006). The large and highly anionic plasmid DNA molecule, however, does not enter cells through simple diffusion. Instead, electrotransfer of plasmid DNA relies on a line of events including (1) electrophoretic migration of DNA towards the plasma membrane, (2) DNA accumulation and transport over the plasma membrane, and (3) intracellular trafficking of the DNA to the nucleus (Favard et al., 2007). For this process to occur, longer electric pulses (milliseconds) are needed (Lucas and Heller, 2001), and thus we have chosen a pulse regimen of a HV and a LV pulse, which has previously been shown to give good transfection efficacy in muscles (Andre et al., 2008).
Role of Ca2+ influx on muscle fitness and homeostasis
Ca2+ plays an important role in muscles as a main regulatory and signaling molecule, and thus the concentration of Ca2+ in the cytosol is tightly controlled and is maintained at 50–100 nM in the resting state (Berchtold et al., 2010). This is in sharp contrast to the extracellular fluid, where the Ca2+ concentration without Ca2+ injection is 1.3 mM. Thus the concentration gradient for Ca2+ across the membrane is very high. Permeabilization of the membrane may lead to massive influx of Ca2+ (Gissel, 2010), which may lead to Ca2+ overload defined as excessive accumulation of Ca2+ in the muscle cell, resulting in loss of control of intracellular free Ca2+. This, in turn, may lead to activation of calpains and possibly PLA2, increased reactive oxygen species production and mitochondrial Ca2+ overload (Nethery et al., 2000; Goll et al., 2010; Sandercock and Mitchell, 2010). All these effects will come into play together, resulting in proteolysis of intracellular components, disruption membrane integrity, and loss of ATP production. This may start a vicious cycle of continuing cell damage and Ca2+ influx and eventually result in cell death (Cerella et al., 2010). Our results indicate that the Ca2+ electrotransfer results in Ca2+ overloading and thus the vicious cycle, resulting in elimination of the transfected fibers.
Conclusion
Gene therapy is rapidly expanding to clinical use, and safe and efficient methods for termination of transgene expression are warranted. In this study, we provide a simple and efficient way of quickly eliminating all transgene expression in muscles by Ca2+ electrotransfer. The massive influx of Ca2+ during Ca2+ electrotransfer disrupts normal cell function, eliminating transgene expression, and induces marked histological changes. This procedure represents a feasible way for quick elimination of transgene expression in emergency situations since (1) only a small volume of muscle will be affected in patients, (2) Ca2+ is a nontoxic molecule, when it is not introduced to cells by electrotransfer, and (3) muscles have a high capacity of regeneration and normal muscle morphology will be restored within 2 weeks.
Footnotes
Acknowledgments
Ruth Rousing, Hanne Villumsen, Lone Boye, Lone Christensen, and Vibeke Uhre are acknowledged for their technical assistance. PH is supported by a grant from the Danish Medical Council, IS is supported by a grant from the Danish Cancer Foundation, HGH is supported by a grant from the Lundbeck foundation, and JG is a research fellow of the Royal Swedish Academy of Science supported by the Acta Oncologica Foundation.
Author Disclosure Statement
All authors declare they have no conflicts of interest.