
Abstract
Despite refinements of medical and surgical therapies, heart failure remains a fatal disease. Myocardial infarction is the most common cause of heart failure, and only palliative measures are available to relieve symptoms and prolong the patient's life span. Because mammalian cardiomyocytes irreversibly exit the cell cycle at about the time of birth, the heart has traditionally been considered to lack any regenerative capacity. This paradigm, however, is currently shifting, and the cellular composition of the myocardium is being targeted by various regeneration strategies. Adult progenitor and stem cell treatment of diseased human myocardium has been carried out for more than 10 years (Menasche et al., 2001; Stamm et al., 2003), and it has become clear that, in humans, the regenerative capacity of hematopoietic stem cells and endothelial progenitor cells, despite potent proangiogenic effects, is limited (Stamm et al., 2009). More recently, mesenchymal stem cells (MSCs) and related cell types are being evaluated in preclinical models of heart disease as well as in clinical trials (see Published Clinical Trials, below). MSCs have the capacity to self-renew and to differentiate into lineages that normally originate from the embryonic mesenchyme (connective tissues, blood vessels, blood-related organs) (Caplan, 1991; Prockop, 1997; Pittenger et al., 1999). The current definition of MSCs includes plastic adherence in cell culture, specific surface antigen expression (CD105+/CD90+/CD73+, CD34−/CD45−/CD11b− or CD14−/CD19− or CD79α−/HLA-DR1−), and multilineage in vitro differentiation potential (osteogenic, chondrogenic, and adipogenic) (Dominici et al., 2006). If those criteria are not met completely, the term “mesenchymal stromal cells” should be used for marrow-derived adherent cells, or other terms for MSC-like cells of different origin. For the purpose of this review, MSCs and related cells are discussed in general, and cell type-specific properties are indicated when appropriate. We first summarize the preclinical data on MSCs in models of heart disease, and then appraise the clinical experience with MSCs for cardiac cell therapy.
Preclinical Experience with MSCs for Heart Disease
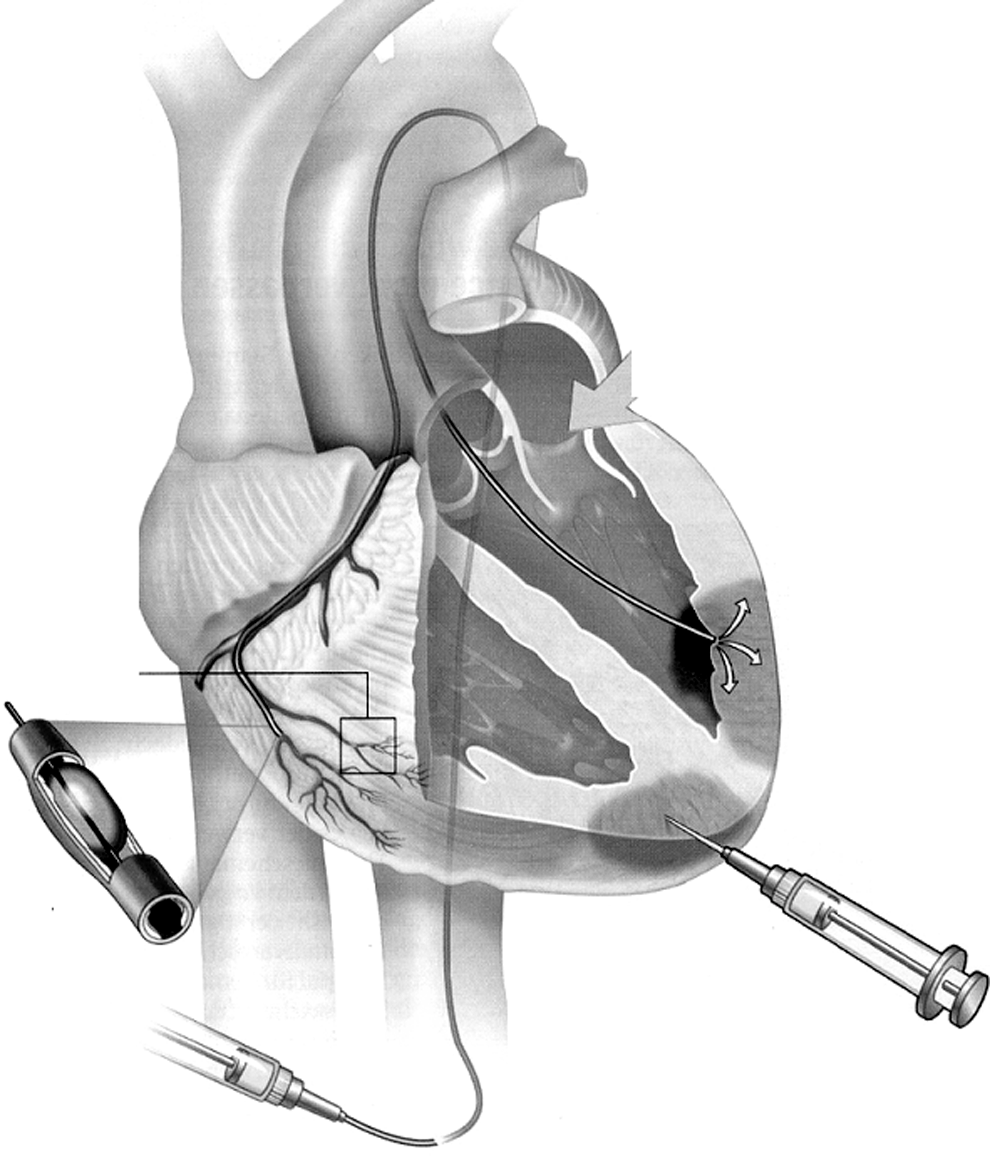
Delivery options for cardiac cell therapy. MSCs can be directly injected into the diseased heart via the epicardium (surgical approach) or transendocardially (usually requiring imaging guiding). Intracoronary injection can also be performed. Not shown here is systemic, intravenous MSC application, which requires active migration of cells to the heart.

Magnetic resonance image of a mouse heart after injection of MSCs labeled with iron oxide. The arrow indicates the area of cell injection, where the iron-loaded cells produce a black signal void. However, no statement can be made regarding function or viability of the cells.
MSCs and cardiomyogenesis
In contrast to typical hematopoietic stem cells, MSCs are able to develop along a myocyte lineage axis. Many in vitro and in vivo studies on MSCs from a wide variety of sources report the expression of several cardiomyocyte-specific markers in stimulated or transplanted MSCs (Toma et al., 2002; Ikegami et al., 2010), and the transcription factors responsible for cardiomyogenic commitment of MSCs overlap with those active during fetal cardiomyocyte development (Arminan et al., 2009; Gao et al., 2010). When human MSCs are prestimulated with a cocktail of “cardiogenic” growth factors and cytokines, their cardiac regeneration capacity in vivo is much improved (“guided cardiopoiesis”) (Behfar et al., 2010). Physical forces such as mechanical strain or electrostimulation also influence the myogenic differentiation capacity of MSCs (Ge et al., 2009; Genovese et al., 2009; Bhang et al., 2010). However, convincing proof of definitive cardiomyocyte differentiation requires a comprehensive evaluation of cardiomyocyte morphology and function including formation of a mature contractile apparatus with cross-striation, spontaneous beating, cardiomyocyte-specific plasma membrane excitation pattern and ion currents, and appropriate intercellular communication and connection structures (i.e., gap junctions, intercalated disks). Quantitative in vitro MSC differentiation into cells of cardiomyocyte morphology and function was first described in 1999 (Makino et al., 1999). It is important to note, however, that epigenetic modulation of the transcriptional profile by DNA demethylation with 5-azacytidine was employed. Demethylation reduces the stability of DNA-silencing signals and thus confers nonspecific gene activation. Under the influence of 5-azacytidine, MSCs obtained a wide range of different phenotypes, of which spontaneously beating myocyte-like cells were isolated and selectively expanded (Hattan et al., 2005). The resulting cell population showed all the morphologic, proteomic, and functional characteristics of true cardiomyocytes (Hakuno et al., 2002). More recently, Feng and colleagues suggested that mainly histone acetylation controls the cardiomyogenic MSC transcription profile, rather than DNA demethylation alone (Feng et al., 2009). However, the effects of chemically induced DNA demethylation and/or histone acetylation on long-term cellular behavior are not known, and may interfere with cell product safety. In fact, even ex vivo culture conditions alone may induce transformation of MSCs with loss of contact inhibition and a pathologic proliferation pattern (Furlani et al., 2009). Cytokines such as bone morphogenic protein (BMP)-2, hepatocyte growth factor (HGF), transforming growth factor (TGF)-β1, and exogenous Jagged1 protein (Notch receptor signaling) induce the expression of several cardiomyocyte marker proteins in human MSCs in vitro and in vivo (Forte et al., 2006; Li et al., 2006; Chang et al., 2008; Herrmann et al., 2010a), and many other strategies to promote cardiomyogenic MSC differentiation have been described, including coculture with mature cardiomyocytes (Okamoto et al., 2007; Ventura et al., 2007; He et al., 2010; Yang et al., 2010a) or cardiomyocyte extracts (Labovsky et al., 2010; Peran et al., 2010). However, many of these concepts remain controversial, and cardiomyogenic MSC differentiation could not be reproduced by other groups (Koninckx et al., 2009; Mastitskaya and Denecke, 2009; Roura et al., 2010). MSC-derived cells with “cardiomyogenic commitment” were found to have greater regenerating effects than undifferentiated MSCs (Shim et al., 2010), but truly convincing strategies for clinically relevant cardiomyocyte production by small molecule stimulation of MSCs without direct epigenetic modification have not yet been described. Taken together, cardiomyocyte-like differentiation of mesenchymal stem cells appears to be possible, provided MSCs are subjected to substantial biologic manipulation in vitro. It is unlikely, however, that physiological in vivo signals are sufficient to drive naive MSCs into a myogenic lineage without significant exogenous influence (Toma et al., 2002; Airey et al., 2004; Hattan et al., 2005). Proof of cardiomyogenic MSC differentiation in in vivo models requires elaborate immunohistology and microscopy techniques, and is prone to induce false-positive results. On occasion, subpopulations of MSCs are described that seem to have greater “stemness” (Jiang et al., 2002; Kogler et al., 2004; Yoon et al., 2005; Lee et al., 2009b). Presumably, such cells are immature progenitors of MSCs, have a higher proliferative capacity, and may be more easily driven to differentiate into various nonconnective tissue phenotypes. However, it is being discussed that extraordinary plasticity may be a phenomenon acquired under long-term culture conditions. Reports describe unusually efficient cardiomyocyte differentiation of placenta-derived MSCs, as well as adipose tissue-derived MSCs (Jumabay et al., 2010), and such neocardiomyocytes regenerated the rat heart across the species barrier (Tsuji et al., 2010). Extraembryonic fetal tissue has been previously noted as a particularly rich source of myogenic MSCs (Ventura et al., 2007), and it remains to be seen whether this approach can be further developed into a clinical cell therapy protocol. MSCs were also shown to stimulate proliferation and differentiation of resident cardiac stem cells, and may thereby indirectly support neomyogenesis (Hatzistergos et al., 2010). As discussed below, the perivascular niche appears to be the most important reservoir of solid-organ resident MSCs, and Galvez and colleagues provided a first link between cardiac progenitor cells and perivascular cells (Galvez et al., 2008). They isolated MSC-like cells termed “cardiac mesangioblasts” by spontaneous in vitro emigration from juvenile mouse hearts, which were Sca-1pos, c-kitpos, CD34pos, CD44pos, CD31pos, and CD45neg, and formed clonal aggregates. When exposed to low serum concentration, they spontaneously differentiated into phenotypically and functionally competent cardiomyocytes. Of note, the authors traced the origin of these cardiac mesangioblasts to a CD31pos/CD34pos and NG2pos subpopulation of perivascular cells that resemble myoendothelial cells from other tissues (see previously). However, the complex relationship between MSCs, perivascular cells, and cardiac stem cells is still incompletely understood.
MSCs and cardiac blood vessel growth
Because the essence of ischemic heart disease is insufficient blood supply, angiogenic processes are of particular importance for myocardial regeneration. On appropriate growth factor and/or cytokine stimulation, MSCs readily acquire an endothelial cell phenotype in vitro, form capillary-like networks in extracellular matrix gels, and also express and secrete proangiogenic growth factors (Chen et al., 2009; Duffy et al., 2009; Garzoni et al., 2009; Rubina et al., 2009). In vivo, increased blood vessel growth has been demonstrated in numerous models of myocardial infarction by both histology and tissue perfusion scans (Z. Li et al., 2009; Poncelet et al., 2010), and can be further augmented by, for instance, treatment of MSCs with epidermal growth factor (Belmadani et al., 2009). In addition to endothelial cell differentiation, MSCs can also participate in the formation of the blood vessel media by forming smooth muscle cells and fibroblasts. In fact, the vascular tree seems to be the predominant reservoir for tissue-specific MSCs. The perivascular niche is increasingly recognized as a major reservoir for mesenchymal stem cells and their putative tissue-specific progeny, a concept that is especially appealing in an organ as highly vascularized as the heart (Tamaki et al., 2002). Perivascular cells have been identified that coexpress myogenic and endothelial cell markers and have a robust myocyte differentiation capacity. Such myoendothelial cells are found among pericytes and have been isolated from various human tissues including skeletal muscle, pancreas, adipose tissue, and placenta. They are discussed as representing the solid-organ reservoir of tissue-specific mesenchymal stem cells and express CD146, NG2, and PDGFRB (platelet-derived growth factor receptor, β polypeptide) but no hematopoietic, endothelial, or myogenic markers (Caplan, 2008; Crisan et al., 2008). Human myoendothelial cells from skeletal muscle have been shown to be more potent for cardiac regeneration in mice than satellite cells or endothelial cells (Okada et al., 2008), and when mesangioblasts from fetal mice were injected into infarcted mouse hearts, they colonized mainly the smooth muscle layer of blood vessels, and exerted protective effects on ischemic cardiomyocytes (Galli et al., 2005).
MSCs and cardiac extracellular matrix remodeling
A striking finding in many experimental and clinical studies on cell therapy for myocardial cell therapy is a change in diastolic function and delay of the progression of scar formation of the infarcted left ventricle (Shake et al., 2002; Schaefer et al., 2006; Dixon et al., 2009; Schneider et al., 2009). This beneficial impact on connective tissue remodeling processes in the diseased heart indicates that MSCs and related cell types may directly interact with extracellular matrix components (B. Jin et al., 2009; Tan et al., 2010). In the field of bone and cartilage regeneration, this has been extensively studied. Mechanical forces (Park et al., 2007; Castillo and Jacobs, 2010) and extracellular matrix components influence MSC biology (Mauney et al., 2010; Sittichokechaiwut et al., 2010; Tan et al., 2010), and MSCs also express and secrete matrix constituents as well as matrix-modulating enzymes (Steingen et al., 2008). In animal models, a beneficial impact on heart failure-induced fibrosis after MSC transplantation was clearly demonstrated (L. Li et al., 2008; Molina et al., 2009). In general, MSCs shift the balance of matrix metalloproteinases (MMPs) and their tissue inhibitors (TIMPs), so that matrix-degrading effects dominate (Mias et al., 2009; Molina et al., 2009). These matrix-remodeling actions may require the secretion of hepatocyte growth factor by transplanted MSCs (Tang et al., 2010), and can be enhanced by overexpression of heme oxygenase-1 (Shu et al., 2010). MSCs act not only directly on extracellular matrix components; they have also been shown to suppress the proliferation of cardiac fibroblasts (L. Li et al., 2009) and to promote their MMP secretion (Mias et al., 2009). However, our understanding of cardiac extracellular matrix biology is only rudimentary, and the exact role of MSCs in restoring a physiological matrix composition is still largely elusive.
Paracrine MSC effects in the heart
Similar to many other somatic progenitor and stem cell types, MSCs secrete a wide spectrum of growth factors and cytokines that can act on surrounding cells in the recipient tissue (reviewed in da Silva Meirelles et al., 2009), and many of the MSC effects described so far involve paracrine activity (Cheng and Yau, 2008; Tang et al., 2010). Whereas proangiogenic paracrine MSC actions in ischemic hearts have been extensively studied (see previously) (Z. Li et al., 2009), the interactions with resident cardiac progenitor cells and extracellular matrix are less well understood. Among the multitude of secreted proteins are vascular endothelial growth factor (VEGF), HGF, insulin-like growth factor (IGF)-1, TGF-β, basic fibroblast growth factor (bFGF), leukemia inhibitory factor (LIF), stroma-derived factor (SDF)-1, angiopoietin, interleukins, and numerous chemoattractants (da Silva Meirelles et al., 2009). Proteomic analysis of conditioned MSC medium revealed that the secretion profile changes depend on factors such as inflammatory milieu, hypoxia, and mechanical stress (Hu et al., 2008; J.H. Li et al., 2008; Thangarajah et al., 2009). da Silva Meirelles and colleagues stratified the paracrine effects of MSCs as trophic (antiapoptotic, supportive of proliferation and/or differentiation, angiogenic), immunomodulatory, antiscarring, and chemoattractant (da Silva Meirelles et al., 2009), most of which are potentially important for myocardial regeneration. In fact, the paracrine MSC actions are powerful enough to influence heart function from remote tissues. Shabbir and colleagues injected a high dose of bone marrow MSCs into the skeletal muscle of hamsters, where they remained trapped but still improved the function of failing hearts including angiogenesis, inhibition of cardiomyocyte apoptosis, and antifibrotic effects (Shabbir et al., 2009). This remote “trophic” effect was linked to MSC-derived IL-6-type cytokines through engagement of the skeletal muscle JAK (Janus kinase)–STAT3 (signal transducer and activator of transcription-3) axis (Shabbir et al., 2010).
MSCs and cardioprotection
Regarding ischemic heart disease, perhaps the most important paracrine MSC action is the secretion of “survival factors” that exert antiapoptotic effects on terminally ischemic cardiomyocytes. When myocytes in culture are subjected to hypoxia, MSC conditioned medium can significantly reduce the number of apoptotic cells (Cselenyak et al., 2010), and MSC-secreted proteins directly influence the function of cardiomyocyte transmembrane ion channels (Benzhi et al., 2009). The responsible signaling cascades in the MSC-protected cardiomyocytes have been partially studied and include activation inflammation- and stress response-associated signaling pathways mediated by IGF-1 and inhibition of the transcription factor NF-κB (Lai et al., 2009; Enoki et al., 2010; Rogers et al., 2010). Rogers and colleagues induced inflammatory damage in cardiomyocyte cultures by incubation with endotoxin or IL-1, and found that MSCs help preserve intracellular calcium handling (Rogers et al., 2010). MSC-induced cardioprotection can also be reproduced in the isolated Langendorff-perfused heart subjected to ischemia–reperfusion injury (Erwin et al., 2009), and the cardioprotective effect is enhanced by preconditioning of MSCs with TGF-β or by activating tumor necrosis factor (TNF) receptor-2 (Herrmann et al., 2010b; Kelly et al., 2010). Evidence suggests that secreted phospholipid particles (exosomes) rather than or in addition to cytokine protein in solution mediate MSC cardioprotection (Lai et al., 2010). In the in vivo setting, MSC cardioprotection is reflected by a reduced number of apoptotic cells in the vicinity of transplanted MSCs (Li et al., 2007) as well as by the preservation of antiapoptotic signaling (H. Li et al., 2009), and can be augmented by genetic modification of MSCs (Gnecchi et al., 2005). Although strictly speaking it is not regeneration, this “salvaging” rather than “regenerating” capacity of MSCs may help explain the frequent in vivo finding of reduced infarct size after MSC therapy in the absence of formation of neocardiomyocytes.
MSCs and immunology in the heart
Acute myocardial infarction as well as many forms of heart failure is associated with immunological processes in the heart, but when and how those are beneficial or detrimental remains unclear. MSCs respond to inflammation throughout the body and migrate to the site of injury, where they can actively suppress immunological processes and reduce inflammation by inhibiting T cell proliferation. Overall, MSCs influence solid-organ injury by shifting the local balance of pro- and antiinflammatory cytokines. For instance, Du and colleagues showed in a rat model of myocardial infarction that MSCs inhibited NF-κB activity, attenuated the production of TNF-α and IL-6, and increased the expression of antiinflammatory IL-10 within the infarcted heart (Du et al., 2008). MSCs also attenuate primary inflammation of the heart as seen in acute myocarditis (Ishikane et al., 2010), as well as cardiac dysfunction during endotoxemia (Weil et al., 2010). However, the immunomodulatory effects of MSCs seem to be transient, dose dependent, and require a certain degree of inflammatory response (Renner et al., 2009). After systemic intravenous application, most MSCs embolize in the lung and spleen, but heart function is still influenced (Assis et al., 2010). One possible mode of action is the induction of antiinflammatory factors from trapped extracardiac MSCs. Indeed, Lee and colleagues showed that TNF-α-stimulated gene-6 (TSG-6) protein is secreted from these cells and clinical improvement of myocardial infarct depended on this effect (Lee et al., 2009a). MSCs not only influence cardiac immunology; they also react to cytokines that are present in the diseased heart (reviewed in Abarbanell et al., 2009). For instance, stimulation of TNF-α receptor (TNFR)-1 and/or -2, which play important roles in heart failure, alters the expression pattern of VEGF, TNF-α, IL-6, and IL-1 in MSCs (Markel et al., 2007, 2008; Bao et al., 2008). TGF-β1 has been shown to induce VEGF expression, in addition to its cardiomyogenic effects (see previously) (Wang et al., 2008), and HGF as well as IGF-1 stimulate the migration of MSCs (Forte et al., 2006). Last, Toll-like receptor-2 has been shown to play a key role in MSC-mediated cardiac recovery (Herrmann et al., 2010a). The complex interplay of MSCs, inflammation, and myocardial regeneration is a fascinating field of research, and is still far from being completely understood.
Allogeneic versus autologous MSCs
As mentioned previously, bone marrow-derived mesenchymal stem cells are to some extent immunoprivileged, and allogeneic MSCs may escape detection and elimination by the host immune system because they do not express MHC class II receptors and only low levels of MHC class I receptors (Grinnemo et al., 2004; Amado et al., 2005). Hence, they might be harvested from young and healthy donors for allogeneic transplantation in the heart. A commercial off-the-shelf cell product is currently in the early clinical testing phase in patients with ischemic heart disease (see Published Clinical Trials, below). Whether MSCs from other sources can also escape the immune system after allogeneic transplantation is under investigation. It is hoped that especially fetal or neonatal cells from the placenta, amnion membrane or fluid, umbilical cord, and cord blood are even less immunogenic and have greater “stemness” because they are more immature than MSC-type cells from adult donors. However, it remains to be seen whether such cells begin to express immunogenic surface antigens on differentiation into cell types needed for cardiac repair. This is specifically important if the therapy aims at a direct regenerative effect of MSCs through differentiation and integration into the heart tissue, for example, in the form of immunoaccessible endothelial cells. Differentiation of allogeneic endothelial precursor cells into endothelial cells in a rat allogeneic aortic transplantation model induced reexpression of MHC class I and II receptors, associated with inflammation and rejection in vivo (Ladhoff et al., 2010). This induced immunogenicity may necessitate the use of immunosuppression to optimize the regenerative MSC effect. Min and colleagues injected cord blood-derived MSCs into infarcted rat hearts, quantified firefly luciferase bioluminescence signals in vivo, and found that retention of human cord blood MSCs in the rat heart is greatly improved by immunosuppression treatment (Min et al., 2006), and similar observations regarding the need for immunosuppression treatment were also made in a pig myocardial infarction model (Poncelet et al., 2010). In another large animal model, a low dose of allogeneic MSC-like cells (STRO-3+ mesenchymal precursor cells) had a greater effect on a left-ventricular remodeling and angiogenesis process than a higher dose (Hamamoto et al., 2009), and it may be speculated that only the low dose of cells was tolerated by the recipient immune system. Another aspect of allogeneic MSC application is the potential sensitization of the immune system against allograft organ transplantation later in life, and particularly on repeated exposure of immunocompetent organisms to allogeneic MSCs, there may be an immunological response (Grinnemo et al., 2004). Pretreatment of MSCs with cytokines, including IL-10 and HGF, in order to modulate immunomodulating properties, differentiation, and immunotolerance may be one option to increase the efficacy of these cells (see Modifying MSCs for Cardiac Regeneration, below). Although these issues need to be thoroughly addressed in clinical trials, HLA matching of mesenchymal stem cells will reduce this potential risk and should be rigorously observed in clinical trials. Access to adequate cell MSC resources for HLA matching may be best provided by abundantly available neonatal tissues such as cord blood or placenta. In a rat model of myocardial infarction, xenogeneic amniotic membrane-derived mesenchymal stem cells transdifferentiated into cardiomyocytes in situ and survived for more than 4 weeks. Immunologic tolerance was caused by HLA-G expression, and activation of FOXP3-positive regulatory T cells (Tsuji et al., 2010). Cord blood and amniotic MSCs may play a role in feto–maternal tolerance, which is modulated by IL-10 and progesterone, and both increased HLA-G expression and also cardiomyocyte transdifferentiation of amnion-derived MSCs (Tsuji et al., 2010).
MSC behavior in age and heart failure
Similar to endothelial progenitor cells and hematopoietic stem cells (Heeschen et al., 2004), MSCs are also subject to age- and disease-related changes (Hacia et al., 2008; Kasper et al., 2009; reviewed in Sethe et al., 2006). This is particularly relevant for cardiac regenerative medicine, because the vast majority of patients are older and chronically ill. Overall, the proportion of MSCs among bone marrow mononuclear cells decreases with age, and MSC yield is lower the older the marrow donor is (Sethe et al., 2006; Tokalov et al., 2007b). Although the fundamental ability to differentiate into fat, bone, and cartilage cells as well as into endothelial cells is preserved in older MSCs (Roura et al., 2006), quantitative differences are revealed on comparison with younger cells (Tokalov et al., 2007a; Fan et al., 2010). For instance, the collagen-secretory activity of MSC-derived chondrocytes from older donors is much reduced (Zheng et al., 2007). On the other hand, Roura and colleagues found no difference in the adipogenic and 5-azacytidine-dependent myogenic differentiation capacity between MSCs from older and younger human donors (Roura et al., 2006). Overall, the gene expression profile of aging MSCs shows an increased abundance of differentiation- and growth arrest-related transcripts and downregulation of transcripts involved in RNA processing (Hacia et al., 2008). Zhang and colleagues specifically studied the impact of donor age on the efficacy of MSCs for cardiac cell therapy in a rat model (Zhang et al., 2005). MSCs from older rats did not improve heart function, whereas the in vivo regenerative capacity of smooth muscle cells from aged donors was preserved. Aged human MSCs also display reduced cardiac regeneration capacity in the rat infarction model (Fan et al., 2010), and one study showed that only MSCs from young and healthy donors can effectively regenerate senescent rat hearts (Khan et al., 2009b). In the clinical setting, the impact of age and disease is more difficult to assess. When we studied the clonality of MSCs from patients requiring cardiac surgery, we found that diabetes mellitus and chronic pulmonary disease were predictors of a higher number of colony-forming units per culture (Choi et al., in preparation). Similarly, when neonatal cord blood MSCs are cultured in the presence of serum from patients with heart failure, the proliferation rate tends to be increased. On the other hand, Grauss and colleagues transplanted MSCs from patients with ischemic heart disease into infarcted murine hearts and found that these clearly exert regenerative effects (Grauss et al., 2007). This apparent mismatch between animal and human data clearly warrants further investigation.
Sources of MSCs for cardiac cell therapy
As mentioned previously, MSCs for cardiac cell therapy can be obtained from manifold sources. Bone marrow MSCs have been studied most extensively and clinical studies have also been performed (see Clinical Cardiac Cell Therapy with MSCs, below). The other major source of MSCs in the clinical setting is adipose tissue. The key criteria of cell products for cardiac regeneration have been demonstrated in vivo and in vitro for MSCs from both sources. The main advantage of adipose tissue MSCs is said to be the availability of large numbers of cells from liposuction aspirate, without the need for expansion in culture, and cell products are currently being tested in clinical studies on patients with heart disease (see Clinical Cardiac Cell Therapy with MSCs, below). On the other hand, it is not clear to what extent such freshly isolated adipose tissue-derived cells are indeed MSCs; hence they are often termed “adipose tissue stem cells.” Cultivated plastic-adherent adipose tissue MSCs with well-defined characteristics are also being used in the clinical setting, but their advantage over bone marrow MSCs remains elusive. In the experimental setting, MSCs from fetal or neonatal sources have been particularly interesting. They are believed to have greater plasticity and lower immunogenicity, but again much of the available data is controversial (reviewed in Benzhi et al., 2009). Cord blood-derived MSCs, as well as their proposed multipotent subpopulations, have been extensively studied in models of heart disease, but their isolation from cord blood can be difficult (Cheng et al., 2003; Bieback et al., 2004; Kim et al., 2005; Yamada et al., 2007). Wharton's jelly of the umbilical cord is also a rich source of MSCs, but those have been studied mainly in the context of heart valve tissue engineering. MSCs from placental tissue as well as MSCs isolated from amniotic fluid have received significant attention; their surprising cardiomyogenic differentiation potential has been discussed previously. MSC-like cells that were derived from cardiac tissue, including cardiac adipose tissue, have also been used successfully for experimental cardiac regeneration, but transfer to the clinical setting is difficult for reasons of source tissue availability (Hoogduijn et al., 2009; Bayes-Genis et al., 2010). An exception are so-called cardiac-derived proliferating cells, which can been obtained in large numbers from endomyocardial biopsy samples, but they do not completely meet the MSC definition (Haag et al., 2010).
Modifying MSCs for cardiac regeneration
Overexpression of antiapoptotic proteins and growth factors, and inhibition of MSC differentiation by blocking Wnt and BMP signaling pathways, have proven powerful strategies to enhance the cardiac regeneration capacity of MSCs (Mangi et al., 2003; Li et al., 2007; Fan et al., 2009; Alfaro et al., 2010; Huang et al., 2010). Although the clinical application of genetically modified MSCs may be problematic, there are also numerous examples of modified MSCs for cardiac cell therapy that do not require interference with the genome or viral vectors. Bartosh and colleagues demonstrated that when MSCs are cultured in three-dimensional aggregates, their antiinflammatory properties are significantly enhanced (Bartosh et al., 2010), and MSCs enriched for Stro-1 expression were also shown to have higher cardiovascular paracrine activity (Psaltis et al., 2010). Hypoxic culture conditions also effectively stimulate the paracrine activity of MSCs and activate survival-relevant signaling cascades (Chacko et al., 2010), as does treatment with heat shock proteins (Chang et al., 2009). Anoxic preconditioning enhanced the antiapoptotic and remodeling capacity of bone marrow MSCs in diabetic cardiomyopathy (J.H. Li et al., 2008), and hypoxic MSC conditioning had similar beneficial effects in rat myocardial infarction (Hu et al., 2008). Preconditioning by oxidative stress, diazoxide-mediated NF-κB activation, and nitric oxide donors has also been shown to enhance the cardiac regeneration potential of MSCs (Rebelatto et al., 2009; Afzal et al., 2010) as well as hyperbaric oxygenation of the recipient heart (Khan et al., 2009a). In addition to enhancing their paracrine activity (see previously), preconditioning also seems to influence MSC adhesion to ischemic myocardium (Song et al., 2009). Such simple preconditioning protocols should be readily transferable to the clinical setting. Adjunct pharmacologic therapy can also support the therapeutic capacity of MSCs in the heart, as has been shown for statins and estradiol in bone marrow MSCs in vitro and in vivo (Erwin et al., 2009; Dong et al., 2010), for sildenafil and adipose tissue-derived MSCs in a rat model of cardiomyopathy (Lin et al., 2010), and for oxytocin in cord blood MSCs (Kim et al., 2010). Phosphodiesterase inhibition also promoted MSC survival and proliferation in infarcted rat hearts (Haider et al., 2010). Mechanical prestimulation of the myocardial vascular bed by ultrasound-induced stimulation of microbubbles was shown to augment engraftment of MSCs (Ghanem et al., 2009). Last, embedding MSCs in extracellular matrix components or artificial polymer scaffolds (J. Jin et al., 2009; Tay et al., 2010; Wall et al., 2010) can facilitate cell retention in the heart and reduce relocation of MSCs to other organs (Huang et al., 2009; Tan et al., 2009), and microencapsulation of MSCs in alginate polymers has also been proposed (Dai et al., 2009; Al Kindi et al., 2010; Yu et al., 2010). Again, this is a strategy with clinical potential, and other combinations of MSCs and carrier substances are under development.
Clinical Cardiac Cell Therapy with MSCs
Published clinical trials
So far, the clinical experience with cardiac cell therapy with MSCs is limited (Table 1). Mainly because of regulatory, financial, and intellectual property restrictions regarding clinical-grade MSC production in line with Good Manufacturing Practice (GMP), most of the clinical cell therapy studies have been performed with freshly isolated bone marrow cells. From animal models, there were concerns regarding ectopic formation of connective tissue in the heart, and microinfarction on intracoronary injection, but, so far, those have not proven clinically relevant (Grieve et al., 2010). As early as 2004, Chen and colleagues published a clinical trial on 69 patients with acute myocardial infarction (Chen et al., 2004). After acute percutaneous coronary intervention (PCI) and recanalization of the infarcted vessel the patients were randomized for intracoronary injection of autologous bone marrow-derived MSCs. There were no MSC-related serious adverse events, and up to 3 months after transplantation the MSC-treated group displayed significantly better regional and global left-ventricular function than the control group. The difference in left-ventricular ejection fraction (LVEF) at 3 and 6 months was approximately 10%, which is similar to the data from trials using other bone marrow-derived cell products. Long-term follow-up data have not been published. From studies using bone marrow mononuclear cells (BMNCs), however, it is known that LV function in the control group tends to further improve over time, eliminating the advantage of the cell therapy group. Subsequently, the same group reported on a randomized trial in patients with severe chronic ischemic cardiomyopathy, using a similar protocol. Again, MSC-treated patients had significantly improved heart function, exercise capacity, and change of New York Heart Association (NYHA) class (Chen et al., 2006). In 2005, Katritsis and colleagues reported on 11 patients with acute myocardial infarction who were treated with a combination of autologous bone marrow MSCs and endothelial progenitor cells (Katritsis et al., 2005). At follow-up, left ventricular function and myocardial perfusion were better in cell-treated patients than in control patients, and scintigraphic imaging provided evidence of cellular repopulation/regeneration of previously nonviable infarct scars. Later, the authors provided electrophysiology and stress echocardiography data on a subgroup of patients with a history of malignant arrhythmia, which indicated a reduced arrhythmogenic potential (Katritsis et al., 2007). In another pilot study, the safety of autologous intracoronary MSC injection into patients with myocardial infarction was confirmed (Yang et al., 2010b). There are also anecdotal reports on autologous MSC treatment in children with severe heart failure (Zeinaloo et al., 2010).
C, controlled; CR, case report; EPC, endothelial progenitor cell; LAD, left anterior descending artery; LVEF, left ventricular ejection fraction; MACE, major adverse cardiac event; MI, myocardial infarction; MSC, mesenchymal stem cell; RDBPC, randomized, double blinded, placebo controlled; RSBPC, randomized, single blinded, placebo controlled; SPECT, single-photon emission computed tomography.
Among the first allogeneic MSC products to reach the clinical arena in the form of a Good Clinical Practice (GCP) compatible trial was Prochymal by Osiris Therapeutics (Columbia, MD). This bone marrow MSC-based cell product was administered intravenously in a double-blinded trial on 53 patients with ST-elevation myocardial infarction (Hare et al., 2009). The allogeneic cell infusion was well tolerated, and there was a statistically significant greater increase in LV ejection fraction and lower incidence of arrhythmias and chest pain (see Table 1). The availability of an allogeneic off-the-shelf MSC product for cardiac cell therapy is clearly an intriguing prospect. However, the previously mentioned issues regarding immunological tolerance need to be studied in greater detail, so that unwanted long-term effects can be ruled out. Allogeneic MSCs from neonatal tissue have also been used in the clinical setting. For instance, Ichim and colleagues reported one case of cardiac cell therapy with allogeneic placenta-derived MSCs combined with umbilical cord-derived CD34+ cells, resulting in profound clinical improvement (Ichim et al., 2008). Systematic safety and efficacy studies, however, have not been reported yet. As mentioned previously, the logistics of GMP-compatible MSC preparation are complex and require the highest safety standards. Chin and colleagues addressed this issue in a feasibility study involving three patients with ischemic heart disease (Chin et al., 2010). Autologous bone marrow MSCs were expanded in an off-site laboratory, cryopreserved, shipped, and thawed. All cell preparations were then found to meet the release criteria and could be injected into the myocardium during coronary artery bypass graft (CABG) surgery.
Ongoing clinical trials
Because of the positive safety and at least promising efficacy data from the above-mentioned trials, the interest in MSCs for clinical cardiac cell therapy remains high. Large-scale multicenter trials are imperative in which the left-ventricular function as a primary end point needs to be revisited, and long-term survival, incidence of late major adverse cardiac events (MACEs), and clinical symptoms need to be considered. Moreover, a detailed analysis of the immunological response to allogeneic MSC products is required. At the time of preparation of this paper,
ADRC, adipose-derived regenerative cell; AMI, acute myocardial infarction; BMC, bone marrow mononuclear cell; CHF, chronic heart failure; DCM, dilative cardiomyopathy; LVAD, left ventricular assist device; LVEF, left ventricular ejection fraction; MSC, mesenchymal stem cell; NRC, nonrandomized controlled; R, randomized; RDBPC, randomized, double blinded, placebo controlled; SAE, sustained ventricular arrhythmias, ectopic tissue formation, or sudden unexpected death; TE-SAE, death, nonfatal myocardial infarction, stroke, hospitalization for worsening heart failure, cardiac perforation, pericardial tamponade, ventricular arrhythmias >15 sec or with hemodynamic compromise or atrial fibrillation.
In addition to the ongoing formal and registered trials, many patients with heart failure are being treated with MSC products worldwide outside of clinical studies, formally classified as compassionate use or simply as clinical routine. In many countries, the legal framework allows conducting cardiac cell therapy for purely medical purposes. For instance, in the Far East, there is a booming market for cell therapy with autologous adipose tissue-derived MSCs, obtained from a small lipoaspirate and expanded in culture. The number of patients thus treated far exceeds that of patients recruited for clinical trials. As long as cell preparation and treatment comply with the local regulatory situation and internationally accepted scientific and medical standards, these activities are acceptable. Unfortunately, however, the scientific community will probably not have access to systematically collected and analyzed data, which would so greatly enhance our body of knowledge on MSCs for the treatment of heart disease.
Conclusion
Within the context of cardiac cell therapy, MSCs seem to be more versatile tools than progenitor cells of hematopoietic/endothelial lineage. Although they most likely do not generate de novo contractile cells in vivo, they can exert numerous beneficial effects on the diseased myocardium. Moreover, they are a robust platform for cell-based delivery of expression products. Their unique immunological profile may permit the use of allogeneic MSCs, and therefore the development of off-the-shelf cell products. Although the use of MSC products has so far proven safe in patients with heart disease, a relevant clinical benefit in terms of heart function, relief of symptoms, and patient survival remains to be demonstrated. The impact of heart disease on both autologous and allogeneic MSC function is highly variable, and patient-specific assays need to be developed to optimize the likelihood of successful cardiac cell therapy. After more than 15 years of research on myocardial cell therapy, the human heart remains an organ that resists regeneration. Cell therapy with unmodified MSCs alone will probably not change this. However, in conjunction with other regeneration tools and for specific forms of heart disease, MSCs may ultimately help improve the outlook for patients with heart failure.
Footnotes
Author Disclosure Statement
No competing financial interests exist.