
Abstract
Obliterative bronchiolitis (OB) is a form of chronic rejection after lung transplantation. Lentiviral vectors (LVs) facilitate long-term gene transduction in many tissues and organs. We hypothesized that lentiviral gene transfer of interleukin (IL)-10, a potent immune-modulating cytokine, to the lung could modulate the alloimmune responses in the lung after transplantation. C57BL6 mice received LVs encoding luciferase, enhanced green fluorescent protein (eGFP), or human IL-10 (huIL-10) through airways and underwent repeated bioluminescent imaging, immunofluorescence imaging, or ELISA of lung tissues, respectively. Luciferase activities peaked at day 7 and were stable after day 28 to over 15 months. eGFP staining demonstrated LV-mediated gene transduction mainly in alveolar macrophages. LV-huIL-10 delivery resulted in stable long-term expression of huIL-10 in the lung tissue (average 3.66 pg/mg at 1 year). Intrapulmonary allograft tracheal transplantation (BALBc→C57BL6) was used as a model of OB. LV-huIL-10 or LV-eGFP were delivered 7 days before transplantation and compared with no LV-transfection group. Allograft airways at day 28 were almost completely obliterated in all the groups. However, at day 42, allograft airways treated with LV-huIL-10 showed a spectrum of attenuation in airway fibrosis ranging from complete obliteration through bubble-like partial opening to complete patency with epithelial coverage in association with a significantly reduced obliteration ratio compared with the other groups (p<0.05). In conclusion, lentivirus-mediated gene transduction is useful in achieving long-term transgene expression in the lung. Long-term IL-10 expression has the potential to attenuate allograft airway obliteration. LV-mediated gene therapy could be a useful strategy to prevent or treat OB after lung transplantation.
Introduction
To overcome these challenges, genetic immuno-modification of allograft lungs using gene therapy is an intriguing strategy (Sato and Keshavjee, 2006). The lung is unique in its accessibility through the airways to achieve localized gene expression in the lung, avoiding systemic effects (Cassivi et al., 1999). Particularly, interleukin (IL)-10 is an attractive candidate cytokine to be used in the lung transplant setting because of its potent immunosuppressive and immunoregulatory profile (Moore et al., 2001). Indeed, for primary graft dysfunction, we have demonstrated the efficacy of adenovirus-mediated IL-10 gene transfer in attenuating the fulminant inflammatory responses and improving post-transplant graft function initially in rat lung transplant models (Fischer et al., 2001, 2003; de Perrot et al., 2003) and then in a preclinical large animal model (Martins et al., 2004). More recently, we have developed a platform to conduct gene transduction through this adenoviral vector to the donor lung ex vivo to repair injured human lungs (Cypel et al., 2009).
IL-10 is also a potentially important cytokine in regulation of the chronic immune responses after lung transplantation. Patients with OB have been reported to have low expression level of IL-10 compared with healthy post-lung transplant patients (Bharat et al., 2007). As proof-of-concept studies, the beneficial effect of IL-10 on the process of allograft airway obliteration has been demonstrated by us and others using adenoviral and Sendai viral vectors in mouse and rat subcutaneous tracheal transplant models of OB. These studies demonstrated prevention or attenuation of allograft airway fibrosis at day 21 or 28 by using gene therapy (Boehler et al., 1998; Shoji et al., 2003), suggesting that IL-10 gene therapy is a useful strategy to prevent or treat OB after lung transplantation.
However, the potential application of gene therapy to a chronic condition such as OB will be limited by the generally short-term transgene expression of conventional gene vectors. Conversely, lentiviral vectors (LVs) enjoy the benefit of long-term and stable gene expression due to genomic integration (Kafri et al., 1997; MacKenzie et al., 2002; Nguyen et al., 2006; Peranteau et al., 2008; Trittibach et al., 2008). LVs also possess an ability to transduce nondividing cells while inducing little immune response against the vector (Kafri et al., 1997). Thus, lentivirus-mediated IL-10 gene therapy targeting the lung could be a novel and ideal strategy to overcome the challenge of OB or chronic graft dysfunction after lung transplantation.
In this present study, we hypothesized that lentivirus-mediated IL-10 gene therapy can lead to extended modulation of alloimmune responses after lung transplantation. To test this hypothesis, we had two objectives: to demonstrate lentivirus-mediated gene therapy to the lung through the airways to the lung and to investigate the effect of lentivirus-mediated IL-10 gene therapy on allograft airways in the pulmonary environment using a mouse intrapulmonary tracheal transplant model of OB. We herein report successful delivery of lentivirus into the lung to induce long-term gene transduction and a beneficial effect of lentivirus-mediated IL-10 gene therapy in the lung in attenuating allograft airway fibrosis.
Materials and Methods
LV production
To construct the LV able to drive human IL-10 expression, the wild-type human IL-10 cDNA was produced by polymerase chain reaction (PCR) from the Invitrogen clone IOH29878, using 5′ and 3′ primers, respectively, containing EcoRI and BamHI restriction enzyme sites. The sequences of the primers forward and reverse were 5′-CCGGAATTCCATGCACAGCTCAGCACT-3′ (ForwardIL10Eco) and 5′-CGCGGATCCAGCTGGGTTCTAGTTTCGT-3′ (ReverseIL10Bam), respectively. The purified product was then ligated into the EcoRI and BamHI sites of the pHR… HIV-1–derived lentiviral backbone with the enhanced green fluorescent protein (eGFP) cDNA deleted. Vesicular stomatitis virus glycoprotein (VSVg)-pseudotyped LV-luciferase, eGFP, and human IL-10 were generated by transient transfection of 293T cells by using the three-plasmid system (LV plasmid constructs, the packaging plasmid pCMV ΔR8.9l, and the VSVg envelope-coding plasmid pMD.G). The transfections were performed with PEI (Sigma-Aldrich). Viral supernatants were harvested 48 hr later and concentrated at 19,000×g for 2 hr at 4°C. The concentrated viral supernatants were serially diluted and titered on 293T cells. Flow cytometric analyses were performed 72 hr later by using a FACS Calibur (BD Biosciences) for eGFP expression, luciferase expression by Xenogen IVIS Imaging System 100 (Xenogen Biosciences), or huIL-10 expression by ELISA kit (Invitrogen). Maps of lentivirus used in this study are shown in Fig. 1.
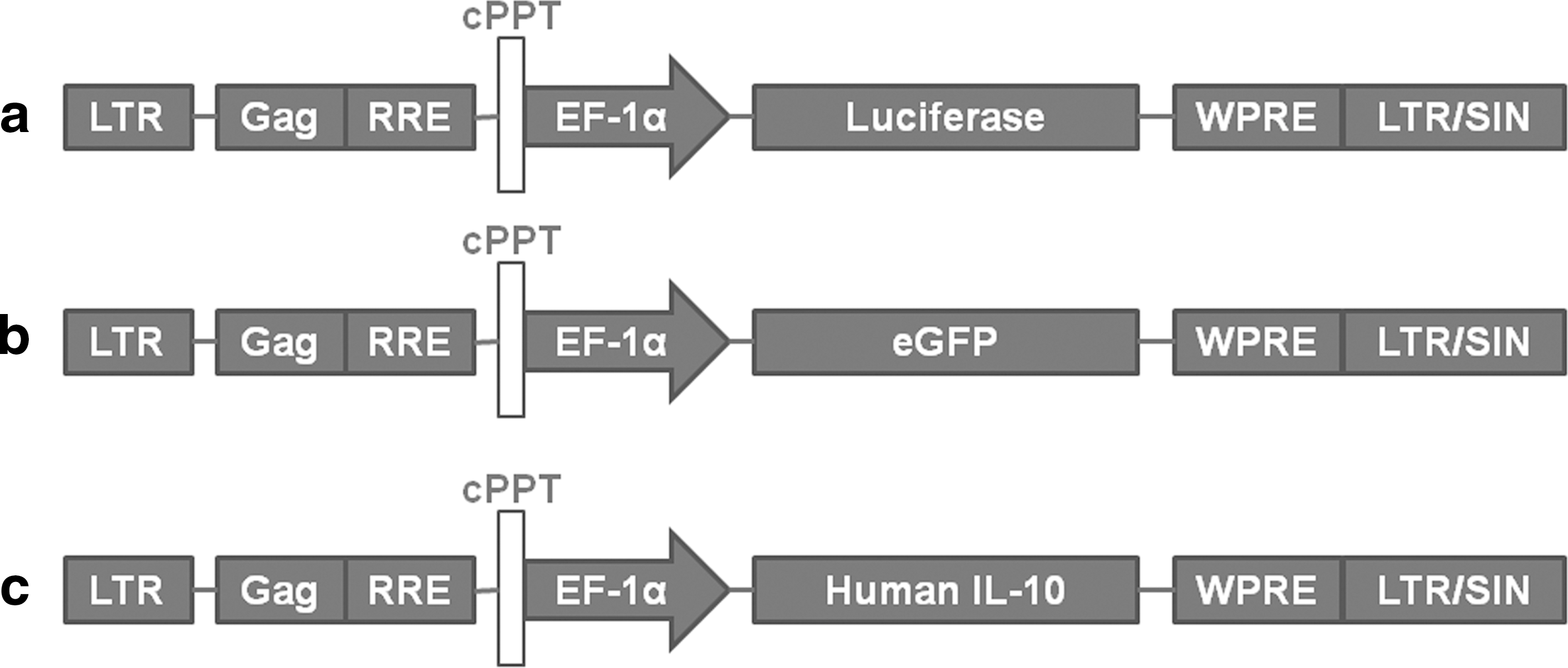
Vector maps. Schematic of LV pHR-cPPT-EF1α-TRANSGENE-WPRE gene transfer vectors used in this study. The transgenes were either luciferase, enhanced green fluorescent protein (eGFP), or human interleukin (IL)-10.
Animals
Five- to 7-week-old female C57BL/6 and BALBc mice (Jackson Laboratory) were maintained on a standard diet and kept in a pathogen-free environment. All animals received care in compliance with the Guide to the Care and Use of Experimental Animals formulated by the Canadian Council on Animal Care. The experimental protocol was approved by the Animal Care Committee of the Toronto General Research Institute. All animal preparations and LV deliveries were approved by University Health Network Biosafety and Animal Care Committees, and performed in a Level 2 biological safety cabinet.
LV transfer in vivo
Animals were anesthetized by intraperitoneally injecting a mixture of ketamine hydrochloride (10 mg/kg, Ketalean, Bimeda-MTC Animal Health Inc.) and xylazine (150 mg/kg, Bayer HealthCare). After adequate anesthesia, endotracheal intubation was performed using a 22-gauge catheter (Terumo) and connected to a small-animal ventilator (Model 693, Harvard Apparatus) using HEPA-filtered air. To deliver lentivirus, the mouse was temporarily detached from the ventilator and a 10-gauge polyethylene tubing (PE10, Intramedic, Clay Adams Brand; BD Biosciences) connected to a 30-gauge needle was inserted into the endotracheal tube. Approximately 40 μl of suspension containing similar p24 titers of lentivirus or normal saline was delivered over 5 sec. The mouse was promptly placed back on mechanical ventilation until spontaneous breathing was observed. Mice were monitored daily for adverse reactions and changes in behavior.
Bioluminescent imaging
We followed the previously described methods (Silvertown et al., 2006). Briefly, D-luciferin (150 mg/kg, Xenogen Biosciences) was injected intraperitoneally into a mouse immediately before bioluminescent imaging. After 10 min, the mouse was placed under the Xenogen IVIS Imaging System 100 (Xenogen Biosciences) with a back-illuminated CCD camera, cryogenically cooled to −105 °C (Spectral Instruments; Princess Margaret Hospital, Toronto, Canada) to obtain images. Images were captured using Living Image software (Xenogen Biosciences). To quantify infectivity, the light emitted from the transduced sites was outlined as the region of interest, and the region of interest photon intensity was the measured using the Living Image software.
Immunofluorescence labeling
We followed the previously described methods (Buckley et al., 2008). Briefly, murine lungs were perfused in vivo with ice-cold 4% paraformaldehyde for 20 min prior to exsanguination to preserve lung architecture. Excised lung were further fixed in 4% paraformaldehyde overnight at 4°C and then paraffin-embedded and cut into 4-μm-thick sections. Sections were immunolabeled with the following antibodies: rabbit anti-eGFP polyclonal antibody (Abcam), rat anti-mouse mac-3 antibody (BD Pharmingen), mouse anti-pancytokeratin antibody (Abcam), and Cy3-conjugated mouse anti-human α-smooth muscle actin (SMA) antibody (1:400, Sigma-Aldrich). Antibody was detected using fluorescently labeled second antibodies; goat anti-rat IgG-Alexa594, goat anti-rat IgG-Alexa 488, and goat anti-mouse IgG-Alexa 555 (Invitrogen) if needed.
Sections were dewaxed and incubated in 4% bovine serum albumin (BSA)/phosphate-buffered saline (PBS) overnight. Primary antibody was added in 2% BSA/PBS for 4 hr and then sections washed serially in PBS before fluorescent secondary antibody was added in 2% BSA/PBS and incubated in the dark for 1 hr. Sections were then washed serially in PBS before being mounted and imaged using a fluorescence microscope.
IL-10 cytokine levels measurement
Tissues were homogenized and incubated at 4°C in cell lysis buffer containing 10 mM HEPES (pH 7.9), 10 mM KCl, 0.1 mM EDTA, 0.1 mM EGTA, 1 mM dithiothreitol (DTT), 0.6% octylphenoxy-polyethoxy-ethanol (Nonidet P-400), 0.5 mM phenylmethylsulfonyl fluoride, aporotinin, and leupeptine. Homogenates were then sonicated and centrifuged at 11,000×g for 10 min at 4°C. Supernatants and plasma samples were assayed using the Human IL-10 UltraSensitive ELISA kit (Invitrogen). The optical density of each well was read at 450 nm according to the manufacturer's instruction with a microplate reader (μQuant; Bio-Tek instruments Inc.) using KC4, version 3.01 (Bio-Tek Instruments Inc). The final concentration was calculated by converting the OD readings against a standard curve. The protein content was determined by using the method of Bradford (1976).
Intrapulmonary tracheal transplantation
This procedure was initially described in rats (Dutly et al., 2005). We applied the technique to mice as follows. In brief, a donor mouse trachea was excised from the cricoid cartilage to tracheal bifurcation. The trachea was then loaded into an 18-gauge Angiocath (Becton Dickenson) the tip of which was cut to produce a sharp needle-wide tip. A recipient mouse was intubated and a left thoracotomy was performed. The prepared 18-gauge Angiocath with the loaded trachea was then inserted into the lung parenchyma of the lower part of the left lung and the injection site was sealed with a small titanium surgical clip (Ethicon).
Morphometric quantification of airway luminal occlusion and α-SMA staining
Luminal occlusion was morphometrically quantified as described previously (Sato et al., 2008). In brief, luminal occlusion was defined as the area containing tissue inside of the cartilage ring. The area of occlusion and the total inner area of the cartilage rings were morphometrically quantified using ImageJ (W. Rasband, National Institute of Mental Health). The former was divided by the latter to yield the occlusion ratio of the graft lumen. Mucus, which is usually produced by airway epithelial cells, in the lumen was not calculated as obliteration. Area positive for α-SMA staining in the graft lumen was morphometrically quantified by ImageJ in a same threshold.
Real-time RT-PCR
Gene expression of IL-6, MIP-2, and IL-1β was examined by real-time reverse-transcriptase (RT)-PCR. RNA was extracted from formalin-fixed paraffin-embedded samples using High Pure FFPE micro RNA kit (Roche Diagnostics). RNA of each sample was converted into cDNA using Transcriptor First Strand cDNA Synthesis Kit (Roche). cDNA was amplified using SYBR Green I Master (Roche) in Roche LightCycler 480 following the manufacturer's instruction. The following primer sequences were used: IL-6: Forward, 5′-CACTTCACAAGTCGGAGGCTT-3′; Reverse, 5′-TCAGAATTGCCATTGCACAAC-3′. MIP-2: Forward, 5′-ACTGCGCCCAGACAGAAGTC-3′; Reverse, 5′-AGTTAGCCTTGCCTTTGTTCAGTATC-3′. IL-1β: Forward, GCAACTGTTCCTGAACTCAAC-3′; Reverse, 5′-ATCTTTTGGGGTCCGTCAACT-3′. GAPDH: Forward, 5′-GGTGAAGGTCGGTGTGAACG-3′; Reverse, 5′-AATCTCCACTTTGCCACTGCA-3′. Data were normalized by GAPDH.
Statistics
Data are expressed as means±SEM. To compare three groups, one-way analysis of variance (ANOVA) was followed by post hoc Tukey tests. All statistical analyses were performed using GraphPad Prism 5.03 (GraphPad Software). Values of p<0.05 are reported to be significant.
Results and Discussion
Technical development of lentivirus-mediated long-term gene expression in the lung
To develop the technique of long-term gene transduction to the lung using LVs, we first tested two types of VSVg-pseudotyped LVs encoding a reporter gene (Fig. 1). After intratracheal administration of luciferase-encoding lentivirus (LV-luciferase), long-term stable luciferase activity localized to the thoracic region, particularly to the left lung was observed over 15 months (Fig. 2A and B). Luciferase activities of treated mice peaked at 7 days and sustained over 15 months after vector administration (Fig. 2C).
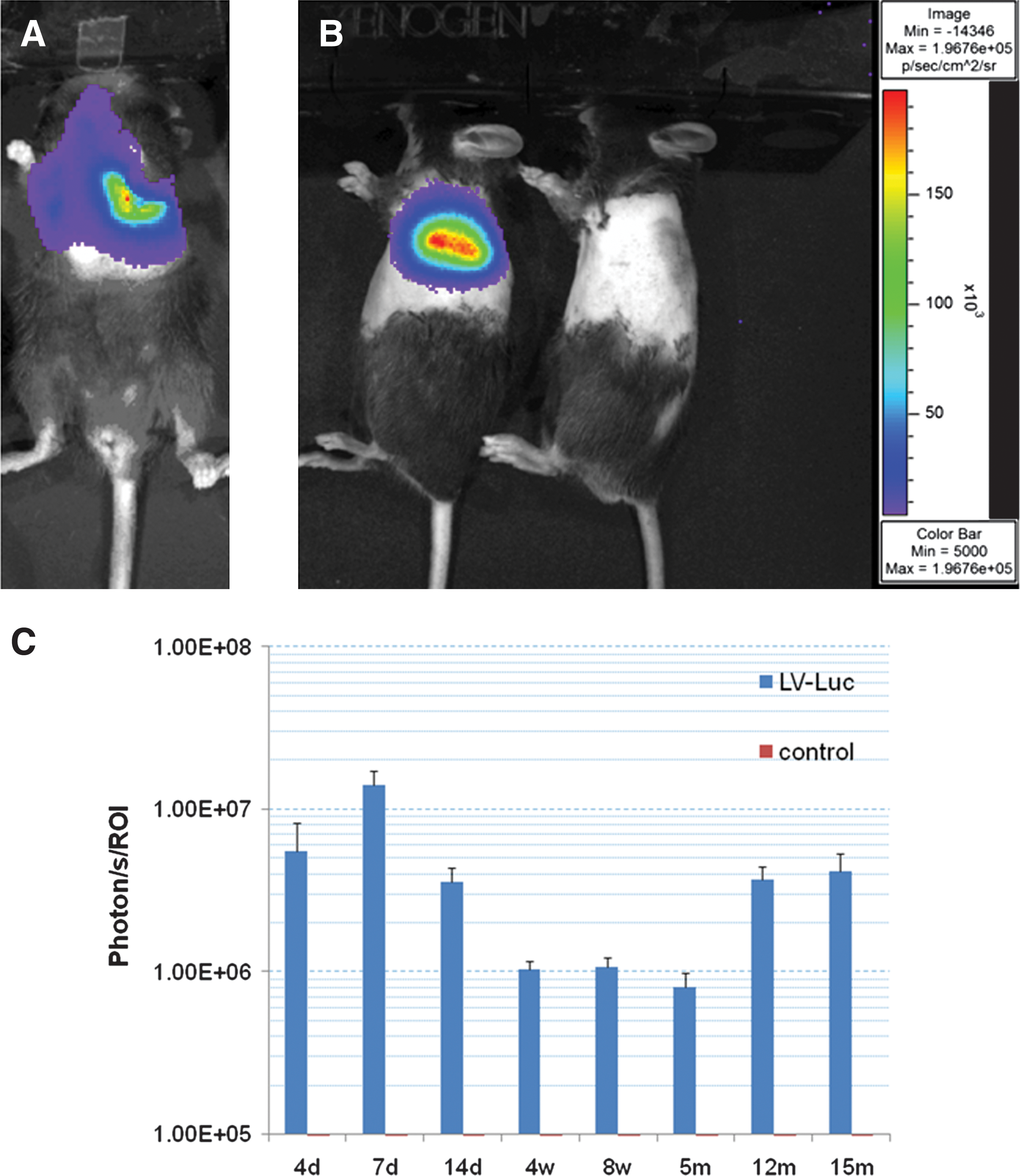
Localized long-term expression of lentivirus-mediated luciferase gene transfer in the lung.
Using LV-eGFP, we examined the cell types in which lentiviral gene transduction occurred. Concordant with previous reports (Borok et al., 2001), immunofluorescence labeling for pan-cytokeratin and eGFP demonstrated that alveolar epithelial cells were transduced with this vector at 8 weeks after infection (data not shown). Interestingly, however, the number of cytokeratin-positive cells seemed relatively minor among all the eGFP-positive cells. Additional labeling with mac-3 (a macrophage marker) revealed that transduced cells are predominantly alveolar macrophages (Fig. 3A). Semi-quantification of mac-3 and eGFP demonstrated that about 70% of transduced cells are macrophages at 8 weeks (Fig. 3B). More interestingly, 6 months after infection, the ratio of double positive cells for eGFP and Mac-3 to eGFP-positive mac-3–negative cells remained consistent (Fig. 3B).

Lentivirus-eGFP transfection on alveolar macrophages in the lung.
This finding initially might be interpreted to be somewhat disappointing, partly due to the assumed short life span of macrophages. However, alveolar macrophages have in fact recently been demonstrated to survive more than 9 months (Murphy et al., 2008). Furthermore, another study has recently demonstrated similar results (Wilson et al., 2010). In that study, VSVg-pseudotyped lentivirus also mainly transfected alveolar macrophages with a comparable range of transduction ratios and resulting long-term expression of their transgene, alpha-1 antitrypsin. Thus, alveolar macrophages appear to be a potentially important target of gene therapy to the lung using VSVg-pseudotyped LVs. Perhaps these cells can actually be envisioned as the “workhorses” implementing gene therapy.
Stable, long-term local gene expression of human IL-10 in the mouse left lung
We then tested VSVg-pseudotyped lentivirus encoding human IL-10 (LV-huIL-10), our target immunosuppressive cytokine of interest. We detected stable expression of huIL-10 in the transfected lung for over a year (Fig. 4). Notably, we found no detectable levels of huIL-10 in the serum over time (data not shown). These results demonstrate that intra-airway administration of LV-huIL-10 enables stable long-term gene expression localized to the lung. Since IL-10 is a soluble protein and systemic effect, which may result in added immunosuppression, is a potential concern (Huhn et al., 1996; Xing et al., 1997), this feature of LV-huIL-10 gene therapy appears ideal to locally treat a chronic pathological condition in the lung.

Long-term expression of human IL-10 in the lung after lentivirus-mediated gene transfection. Human IL-10 levels in the left lungs from mice (n=4 per time point) treated with lentivirus-human-IL-10 were measured by ELISA. No statistical difference was found among time points. Average measurements of untreated mouse left lungs (n=4) were subtracted as background.
Lentivirus-mediated IL-10 gene therapy attenuates allograft airway obliteration
To examine whether lentivirus-mediated IL-10 gene therapy exerts a beneficial effect on the process of allograft airway obliteration in the pulmonary milieu, we used a mouse intrapulmonary tracheal transplant model. We pretreated recipient animals with lentivirus encoding huIL-10 or eGFP before intrapulmonary tracheal transplantation and the result was also compared to a no gene therapy group (Fig. 5A). The levels of huIL-10 in the lung at posttransplant days 28 and 42 were comparable to those detected in the lung without transplantation (posttransplant day 28, 6.53±0.85 pg/mg; day 42, 4.94±0.85 pg/mg; see Fig. 4 for huIL-10 levels without transplantation).
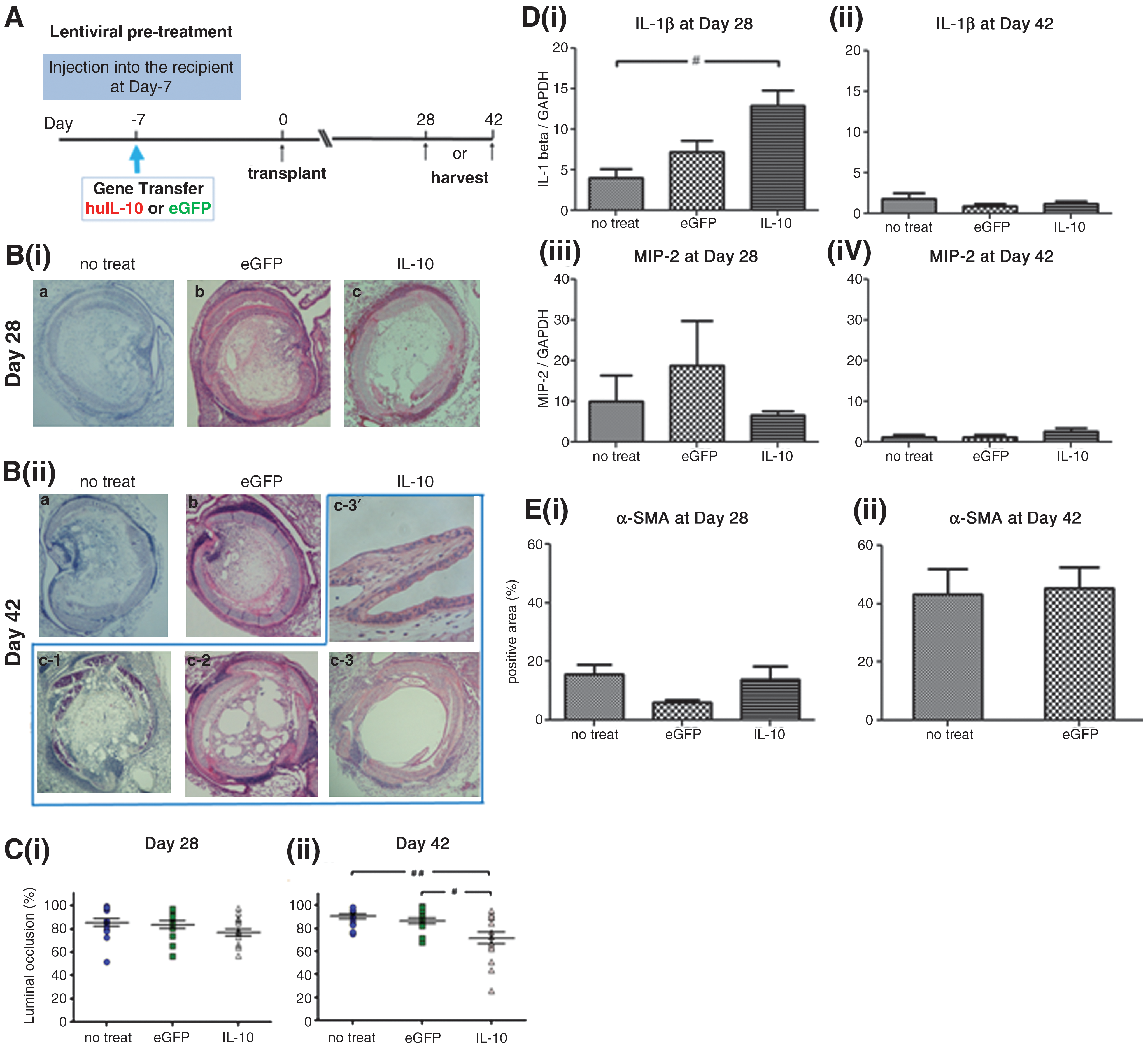
The effect of lentivirus-mediated long-term expression of human IL-10 on allograft airway fibrosis.
At day 28, allograft airways in all the groups were almost completely obliterated (Fig. 5Bi); however, 2 out of 15 allografts treated with LV-huIL-10 showed partially preserved epithelial cells (data not shown). Morphometric quantification of the luminal obliteration ratio did not reveal any significant difference among groups (Fig. 5Ci). Interestingly, at day 42 we found bubble-like or honeycomb-pattern openings in the lumen of some allografts treated with LV-huIL-10 (Fig. 5Bii). Moreover, we found a preserved epithelial lining in 4 out of 15 allografts treated with LV-huIL-10 at 42 days, but not in other groups (Fig. 5Bii c-3). Morphometric quantification of the obliteration ratio at day 42 showed a significantly lower obliteration ratio in the LV-huIL-10 group compared with the LV-eGFP or no gene therapy group (Fig. 5Cii).
We further examined the effect of IL-10 gene therapy on inflammation and fibrosis. Real-time RT-PCR analysis demonstrated general declines in inflammatory cytokines MIP-2 and IL-1β from day 28 to day 42 (Fig. 5D); IL-6 was not detectable at either time point in any group. There was a trend of reduced MIP-2 expression in the huIL-10 gene therapy group (Fig. 5Di), but the same group showed significantly higher expression of IL-1β at day 28 compared with the no-treatment control (Fig. 5Diii), suggesting that a complex immunomodulatory mechanism was involved in the long-term IL-10 gene therapy. IL-10 gene therapy did not reduce α-SMA staining (Fig. 5Ei) at day 28 compared with other groups, suggesting that IL-10 gene therapy does not have a significant impact on the kinetics of early fibrosis. For unclear reasons, the eGFP group showed reduced α-SMA staining at day 28 (Fig. 5Ei). In the no-treatment and eGFP groups, the area positive for α-SMA (Fig. 5Eii) and piaosirius red (PSR) collagen (data not shown) increased from day 28 to 42, although the result was not comparable with the IL-10 group because of the partial luminal opening (Fig. 5C).
The results demonstrate that lentivirus-mediated IL-10 gene therapy has beneficial effects on allograft airway fibrosis in the lung. Given that the intrapulmonary tracheal transplant model of OB is an animal model of allograft airway rejection (Dutly et al., 2005) and that IL-10 is a well-recognized immunosuppressive cytokine, the result appears to support our original hypothesis. However, strikingly, the effect of IL-10 gene therapy was not evident at the early time point (i.e., day 28). The present observation suggests that near complete airway obliteration at day 28 might be reduced or attenuated over time by long-term local IL-10 expression through an unknown mechanism. The spectrum of histological findings among allograft airways treated by IL-10 gene therapy at day 42 (Fig. 5Bii, c-1 to c-3) might suggest different stages of fibrosis resolution—from bubble-like partial openings to complete re-canalization of the airway lumen.
This finding is fundamentally different from previous studies using IL-10 gene therapy (Boehler et al., 1998; Shoji et al., 2003) and many other experimental treatments for allograft airway rejection in that these studies only demonstrated the preventive effect of treatments. Established allograft airway fibrosis has been considered to be a static and irreversible condition. By using long-term IL-10 gene therapy, however, it might in fact be possible to modify and treat advanced disease of OB after lung transplantation.
The mechanistic basis of the effect of IL-10 remains to be explored further. Although contribution of immunosuppressive effect of IL-10 (Moore et al., 2001) or induction of regulatory T cells (Groux et al., 1997) are possible explanations for the beneficial effect of long-term IL-10 expression, modulation of tissue remodeling is another important possibility. As we previously demonstrated, allograft airway fibrosis is not a static but a dynamic process undergoing further remodeling after the obliteration of the airway lumen is completed (Sato et al., 2008). The results of α-SMA staining suggests that IL-10 had little effect on fibrosis by day 28, but the effect became obvious at a later time point when fibrosis was undergoing further maturation. Intriguingly, Nakagome et al. (2006) have shown that IL-10, which was delivered by liposome-based plasmids, attenuates bleomycin-induced pulmonary fibrosis through alveolar macrophages by inhibiting the production and activation of TGF-beta. Since TGF-beta is a potent profibrotic cytokine (Leask and Abraham 2004), regulation of TGF-beta or other factors involved in tissue remodeling and fibrosis by IL-10 is an interesting possibility to be explored further.
The present approach using LV-huIL-10 is preliminary in that there is room for optimizing the timing of lentivirus gene delivery and examining the effect of repeated administration of LVs to maximize the effect. Also, the animal model has multiple limitations as a model of human OB after lung transplantation (Sato et al., 2009). However, we herein demonstrated the feasibility of long-term local lentivirus-mediated IL-10 gene therapy targeting the lung to attenuate allograft airway rejection. Based on this proof-of-concept study, we are further exploring better gene therapeutic strategies using lentivirus and mechanisms of the beneficial effect of IL-10 on allograft airway fibrosis.
In actual clinical settings, administration of LVs could be conducted during ex vivo lung perfusion with or without concurrent adenoviral-mediated IL-10 gene therapy (Cypel et al., 2009). Adenoviral vectors are advantageous in regulating acute-phase inflammation by rapidly achieving high levels of gene expression but only for a short period of time, whereas LVs enable long-term gene expression to cover the period after the effect of adenoviral gene therapy has been lost. Furthermore, the less immunogenic property of LVs may also enable repeated gene transduction, whereby a higher level of gene expression could be achieved or an ideal level of gene expression could be maintained for a longer time period. As such, our adenoviral and lentiviral gene therapeutic strategies with or without ex vivo lung perfusion are complementary to each other, aiming at the same purpose—preventing and treating damage to lung allografts.
Footnotes
Acknowledgments
This work was supported by a research fellowship from the International Society for Heart and Lung Transplantation to Dr. Shin Hirayama. We thank Orlay Lopez-Perez and Cindy Guo for assistance with vector constructions.
Author Disclosure Statement
No competing financial interests exist.