
Abstract
The capsid protein synthesis in targeted tissues resulting from residual contaminating replication-competent adeno-associated virus particles (rcAAV) remains a concern for hazardous immune responses that shut down the factor IX expression in the hemophilia B clinical trial. To systematically reduce/eliminate the effects of potential contaminating rcAAV particles, we designed a novel adeno-associated virus (AAV) helper (pH22mir) with a microRNA binding cassette containing multiple copies of liver-specific (hsa-mir-122) and hematopoietic-specific (has-mir-142-3p) sequences to specifically control cap gene expression. In 293 cells, the rep and cap gene from pH22mir functioned similarly to that of conventional helper pH22. The vector yields and compositions from pH22mir and pH22 were indistinguishable. The performance of vector produced in this new system was comparable to that of similar vectors produced by conventional methods. In the human hepatic cell line, the capsid expression was reduced significantly from cap-mir cassette driven by a cytomegalovirus promoter. In the liver, 99.9% of capsid expression could be suppressed and no cap expression could be detected by western blot. In summary, we demonstrated a new concept in reducing de novo capsid synthesis in the targeted tissue. This strategy may not only help AAV vectors in controlling undesirable capsid gene expression, but can also be adopted for lentiviral or adenoviral vector production.
Introduction
A typical method for rAAV production includes an AAV helper plasmid, an adenoviral mini-plasmid along with a vector plasmid (Ferrari et al., 1997). The adenovirus mini-plasmid supplies all necessary adenovirus helper functions for rAAV packaging except E1a and E1b, which are expressed constitutively by the host 293 cells. The AAV helper plasmid provides the essential AAV rep and cap genes. The rep gene products are essential for AAV DNA replication and packaging. The cap genes encode the structural products that form AAV capsids. In contrast, an AAV vector plasmid has only two copies of AAV ITRs along with the transgene expression cassette. The AAV helper plasmid is often engineered so that it does not share significant homology with the AAV vector plasmid to avoid potential homologous recombination events between them, which may give rise to the formation of replication-competent AAV (rcAAV) particles (Allen et al., 1997; Wang et al., 1998; Wright et al., 2003).
The rcAAV particle is an undesirable contaminant that may affect transgene expression or elicit a hazardous immune response (Manno et al., 2006; Murphy et al., 2008; Hauck et al., 2009; Li et al., 2009a; Pien et al., 2009). Various strategies have been proposed to minimize the homologous events that lead to rcAAV formation. Some strategies proposed are rather complicated and may negatively affect the rAAV production yield or vector performance (Allen et al., 1997; Wang et al., 1998; Wright et al., 2003). However, nonhomologous recombination between the AAV helper plasmid and AAV vector sequences is another significant source of rcAAV particles, which can not be avoided in any current rAAV production method (Wright et al., 2003). In this study, we have designed an rAAV production system to tightly control AAV rep and capsid activity from rcAAV particles. The new AAV helper expression cassette, with tissue-specific microRNA (miRNA) target sequences, does not have any effects on rAAV yield or affect rAAV vector performance. In addition, it is compatible with all current rAAV production systems and serves as a “free” insurance policy for potentially hazardous rep and cap expression from rcAAV particles.
Materials and Methods
AAV helper plasmid construction
The hsa-mir-122 and has-mir-142-3p sequences were obtained from the miRNA registry (Griffiths-Jones, 2004). The mir-122 target cassette was annealed by the following oligonucleotides: 5′-CCA TTG TCA CAC TCC AGT CAC AAA CAC CAT TGT CAC ACT CCA GCT AGC CAA ACA CCA TTG-3′; 5′-GCA TGC TGG AGT GTG ACA ATG GTG TTT GAG CTT GGA GTG TGA CAA TGG TGT TTG GCT AGC-3′. The mir-142-3p target cassette was annealed by the following oligonucleotides: 5′-GCA TGC TCC ATA AAG TAG GAA ACA CTA CAC GAT TCC ATA AAG TAG GAA ACA CTA CAA CCG-3′; 5′-CCT ACT TTA TGG AGT GAT GTA GTG TTT CCT ACT TTA TGG AAC CGG TTG TAG TGT TTC CTA-3′. The annealing reaction was carried out by a PCR Thermocycler (Eppendorf, Westbury, NY) using the following cycles: 94°C for 5 min; four cycles of 94°C for 30 sec, 60°C for 30 sec, 68°C for 30 sec; and a final 68°C, for 5 min in a 50-μl reaction system in PuReTaq Ready-To-Go PCR Beads (Amersham-Pharmacia Biotech, Piscataway, NJ) according to the manufacturer's protocol. The mir-122 target and mir-142-3p target cassette contains four copies of a sequence designed to be perfectly complementary to the respective miRNA. Then the miRNA target sequences were subcloned into the BaeI and SpeI sites of pH22 (AAV2/2 helper plasmid) to generate the pH22mir helper plasmid. Plasmids pH22mir122 and pH22mir142 were generated by a single restriction enzyme cut combination of SphI/AgeI and NdeI/SphI designed in annealed oligonucleotides, respectively. The mir-122 target and mir-142-3p target cassettes were also cloned to an AAV2 capsid expression plasmid driven by the cytomegalovirus (CMV) promoter using the same strategy above to obtain pCMV-AAV2mir142, pCMV-AAV2mir122, and pCMV-AAV2mir.
Recombinant AAV vector preparation
rAAV vectors with LacZ or human factor IX (hFIX) gene were produced by the triple plasmid cotransfection method as described previously (Lu et al., 2008; Chen et al., 2009). AAV vectors were packaged using pH22 (AAV2 rep and AAV2 cap genes) or pH22mir helper plasmids. In brief, AAV helper plasmid, adenovirus function helper plasmid, and AAV-CMV-LacZ or AAV-ApoE-hAAT-hFIX vector plasmid were cotransfected into 293 cells cultured in roller bottles at a ratio of 1:1:1. The transfected cells were harvested 3 days later. AAV vectors were purified by two rounds of CsCl gradient ultracentrifugation. After the collected AAV vectors were buffer-exchanged extensively against PBS with 5% D-sorbitol, vector purity and genome titer were analyzed by silver staining and real-time PCR (Applied Biosystems, Forster City, CA), respectively. The final vectors were stored at −80°C before administration. Vectors produced using pH22mir were identified by adding mir to the vector name.
Tissue culture and transfection
For transfection, HEK 293 and HHL5-B7 cell lines were obtained from the American Type Culture Collection (Manassas, VA) and cultured in Dulbecco's modified Eagle medium (DMEM) (Invitrogen, Carlsbad, CA) with 10% fetal bovine serum (FBS; HyClone, Logan, UT), penicillin (100 U/ml) (Invitrogen), and streptomycin (100 μg/ml) (Invitrogen) at 37°C in a moisturized environment supplied with 5% CO2. Transfections were carried out using Lipofectamine 2000 (Invitrogen) following the manufacturer's instructions. After transfection, the cells were grown in DMEM with 2% FBS to minimize cell death. For direct comparison, a green fluorescent protein (GFP) expression plasmid was also included in the transfection mix to ensure similar transfection efficiency. Cells were harvested at 72 hr post transfection for further analysis.
For AAV vector transduction in vitro, HeLa cells were seeded at 80% confluency in a six-well plate. Twenty-four hours later, AAV2-CMV-LacZ or AAV2mir-CMV-LacZ vectors were added to wells at a dose of multiplicity of infection (MOI) of 10,000 and 1,000. Histochemical staining was performed 48 hr after infection. To determine the transduction efficiency, LacZ-positive cells were quantified from five random fields under × 20 magnification with a DIAPHOT 200 microscope (Nikon, Melville, NY).
Animal procedures
Adult C57BL/J6 mice (25 g) were obtained from the Jackson Laboratory (Bar Harbor, ME); they were housed in a pathogen-free environment and supplied with a normal diet in the Children's Hospital of Philadelphia.
For muscle-directed gene transfer, AAV2-CMV-LacZ or AAV2mir-CMV-LacZ vectors were injected into the left tibialis anterior of 4-week-old C57BL/J6 mice at a dose of 5 × 1011 vector genomes (vg)/injection site. Muscle tissues were harvested and embedded in Tissue-Tek O.C.T. (optimal cutting temperature) compound (Sakura Finetek, Torrance, CA) 4 weeks after vector delivery. Twenty-micrometer sections were cut and subjected to X-gal staining procedures.
For liver-targeted gene transfer, AAV2-ApoE-hAAT-hFIX or AAV2mir-ApoE-hAAT-hFIX vectors were injected via the tail vein at a dose of 1 × 1012 vg/mouse. Plasma was then collected retro-orbitally at a predetermined schedule after vector delivery.
For hydrodynamic injection, animals were injected via the tail vein with 0.125 ml of saline per gram of body weight, containing 100 μg of pCMV-AAV2mir, pCMV-AAV2mir122, pCMV-AAV2mir142, or pCMV-AAV2 plasmid with 30 μg of column-purified pAAV-CB-EGFP plasmid within 5–10 sec. The animals were allowed to recover before being returned to their cages. Mice were sacrificed by CO2 at 24 hr after hydrodynamic injection, and the liver tissue was harvested. The liver tissue was then frozen with liquid nitrogen, kept at −80°C, and subsequently analyzed by western blot.
Histochemical staining
For X-gal staining, HeLa cells or frozen muscle sections were fixed in 2% formaldehyde/0.2% glutaraldehyde/PBS at 4°C for 5 min. After being washed with PBS three times, cells or muscle sections were incubated in 5 mM potassium ferricyanide/5 mM potassium ferrocyanide/2 mM MgCl2/1 mg/ml X-gal in PBS at 37°C overnight.
ELISA for hFIX detection
Citrated plasma samples were collected and assayed biweekly for the expression of hFIX in the C57BL/J6 mice described above. hFIX levels were determined by ELISA using a coating antibody (Sigma) able to recognize hFIX in mouse plasma (Herzog et al., 1997). Plates were washed with 0.05% Tween-20/PBS, blocked with 5% dry milk in PBS, incubated with diluted mouse plasma samples, and detected by 1:2,000 dilution of affinity-purified horseradish peroxidase–conjugated goat anti-hFIX antibody (Enzyme Research Laboratories, South Bend, IN). o-Phenylenediamine (Sigma) buffer was freshly prepared and added to each after the last washing. Levels of hFIX were determined by OD450 and quantified against the linear standard curve generated with serially diluted purified human plasma factor IX (Wyeth, Philadelphia, PA).
Western blot
The western blot of the AAV capsid and enhanced GFP (EGFP) was performed on cell lysate or mouse liver tissues. To analyze AAV capsid expression, cells or liver tissues were lysed with radioimmunoprecipitation buffer (50 mmol/L Tris–HCl, pH 7.4, 150 mmol/L NaCl, 1 mmol/L phenylmethylsulfonyl fluoride, 1 mmol/L EDTA, 1% Triton X-100, 1% sodium dodecyl sulfate). After sonication to break the genomic DNA, 4 × sample buffer and reducing agent (Invitrogen) were added and boiled for 5 min. Protein concentration was then determined by Coomassie (Bradford) protein assay kit (Pierce, Rockford, IL) according to the manufacturer's protocol. Fifty micrograms of the resulting samples were analyzed using 4–12% gradient sodium dodecyl sulfate–polyacrylamide gel electrophoresis gels (Invitrogen) and transferred to nitrocellulose membrane (Invitrogen) for western blotting. After the membrane was blocked for 1 hr in PBS (Invitrogen)/5% dry milk powder/0.05% Tween 20, a mouse anti-AAV capsid antibody (American Research Products, Belmont, MA) at a 1:50 dilution was added and incubated overnight. The membrane was then washed, incubated with a sheep anti-mouse antibody (1:1,000 dilution; Sigma), and developed using an enhanced chemiluminescent substrate (Amersham-Pharmacia Biotech). The expression of EGFP was also analyzed by western blot, except that a mouse anti-EGFP antibody (Invitrogen) was added after block.
Statistical analysis
All data are shown as means ± SD. Analysis was performed using two-tailed Student's t tests and one-way ANOVA with Bonferroni multiple comparison posttest. The differences were considered significant when p was <0.05. Analysis was performed using the SPSS 11.0.
Results
Construct novel AAV helper with miRNA targeting sequences to control capsid gene expression
To reduce/eliminate the capsid gene expression from the potential rcAAV particles, we added a new feature to the AAV helper expression cassette. As shown in Fig. 1, four copies of hsa-mir-122 and has-mir-142-3p sequences were cloned into an AAV helper construct to obtain pH22mir. In pH22mir, miRNA targeting sequences were located right after the stop codon of the cap gene, but before the poly(A) sites. In contrast to the traditional AAV helper construct (pH22), full rcAAV particles generated from pH22mir would also carry these two special miRNA targeting sequences, because they are an integral part of AAV helper expression cassettes. The recombinant particles that do not include miRNA target sequences would not have the poly(A) sequences, and thus would be defective for expression of the viral gene products. In the rcAAV particles with the poly(A) site, miRNA targeting sequences would be present to control the AAV viral protein expression.

Schematic representation of conventional helper plasmid for AAV2 (pH22) and improved AAV helper plasmid (pH22mir). AAV rep and cap open reading frames are indicated in the figure. In the pH22mir, four copies of mir122 and four copies of mir142-3pT sequences are cloned to the 3′ end of the cap region.
AAV helper expression cassette with hematopoietic and hepatic miRNA target sequences supported efficient production of AAV vectors in 293 cell line
One key requirement for the novel helper cassette in pH22mir is that it could provide optimum levels of rep and cap gene products necessary to support rAAV production. To test the functionality of pH22mir, we compared the production of rAAV-CMV-lacZ vector using pH22 or pH22mir. The vector production results using our typical triple plasmid transfection method are presented in Fig. 2. After CsCl gradient ultracentrifugation, viral capsids in each fraction were analyzed by silver staining. There was no alteration in the physical composition of the viral capsid in each fraction (Fig. 2A). Moreover, the vector distribution and yield were similar at each density of the gradient. These results suggested that rAAV vector production was not affected by including the hematopoietic and hepatic miRNA target sequences in the AAV helper expression cassette.
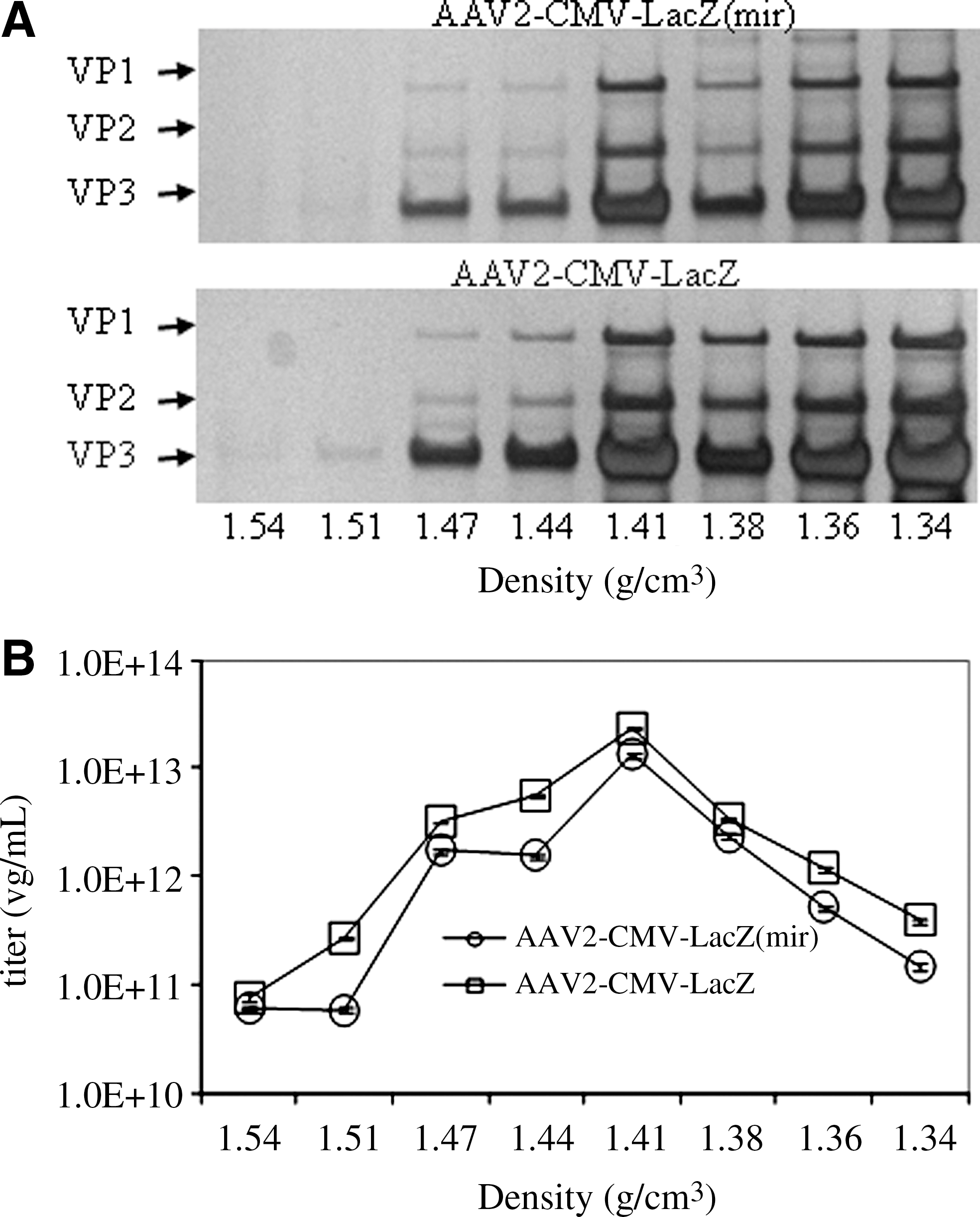
pH22mir efficiently supported AAV vector packaging. Vector plasmids, pAAV-CMV-lacZ, and adenovirus helper plasmid were cotransfected into 293 cells along with pH22 or pH22mir. The transfected cells were then collected at 72 hr post transfection, and the vectors were purified by CsCl gradient. At the end of ultracentrifugation, the gradient was collected at 1 ml per fraction and the vectors in each fraction were analyzed. (
To determine whether vectors produced by using pH22mir are functional in vitro, AAV2-CMV-lacZ produced by traditional pH22 or pH22mir was used to transduce HeLa cells. At the two MOIs tested, both vectors achieved similar levels of transduction efficacy (Fig. 3). The same vectors were also tested for their transduction potency in vivo. AAV2-CMV-lacZ vectors produced by either method were injected into the left tibialis anterior muscle of C57BL/J6 mice at a dose of 5 × 1011 vg/injection site. The muscle tissues were harvested 4 weeks post administration, and the lacZ expression was detected by X-gal staining. As shown in Fig. 4A, no expression difference was observed between vectors. To characterize the vector performance in the liver, we generated factor IX vector under the control of an apoE-hAAT liver-specific promoter. As presented in Fig. 4B, the expression profiles from both batches of vectors were indistinguishable in the liver. It showed a typical expression profile of rising continuously until it reached a plateau at 4–6 weeks after tail-vein injection of vectors. The experiments confirmed that the vectors produced using an AAV helper carrying specific miRNA targeting sequences were able to confer the “normal” behavior of AAV vectors.
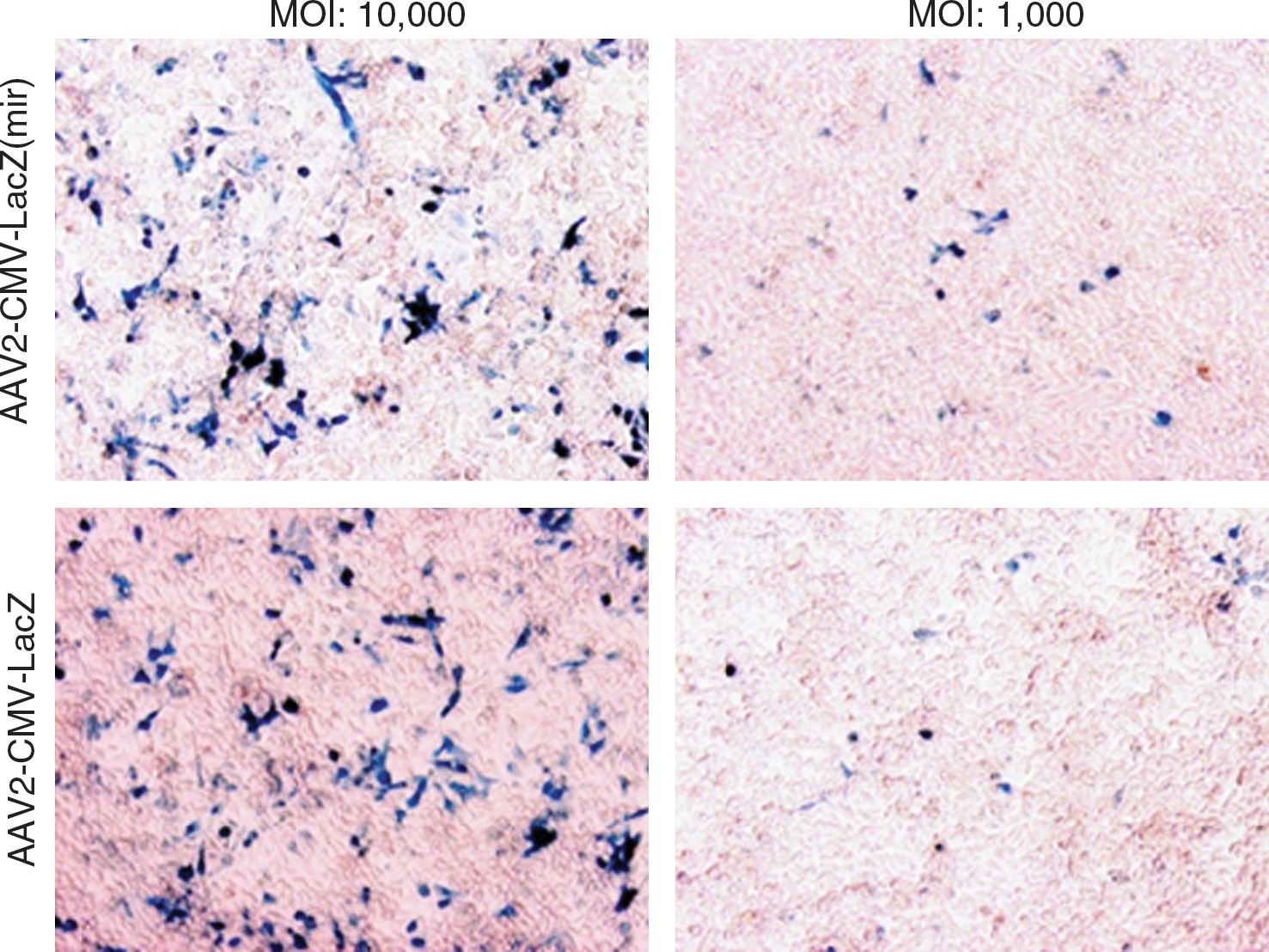
Vector produced by pH22mir mediates efficient transduction in vitro. HeLa cells were infected with AAV2mir-CMV-LacZ or AAV2-CMV-LacZ vectors at an MOI of 10,000 and 1,000. The expression of LacZ was analyzed at 72 hr post infection by X-gal staining.
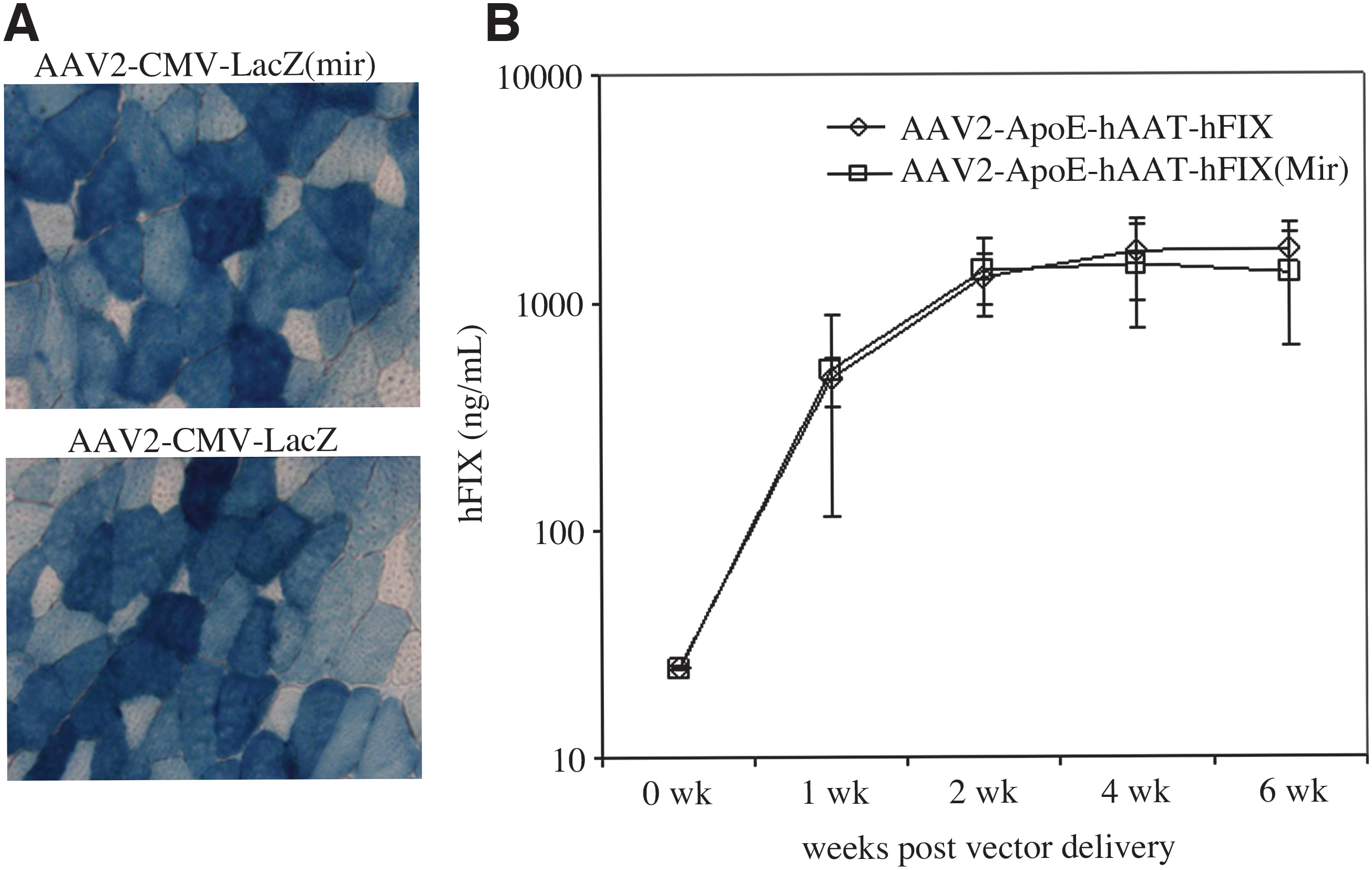
Vector produced by pH22mir mediates efficient transduction in mouse muscle and liver. (
AAV helper expression cassette with miRNA target sequences efficiently suppressed de novo cap gene expression in hepatic cells
We then attempted to demonstrate the potential benefits of including various miRNA targeting sequences in the AAV helper expression cassette. As the generation of rcAAV particles through nonhomologous recombination is a low-frequency event, the potential problems in clinical trials have always been difficult to demonstrate in animal studies. The human clinical trial of hemophilia B has suggested that AAV capsid may have been the culprit for the loss of therapeutic gene expression. The low level of cap expression from minor rcAAV particles is difficult to detect through conventional assays. Hence, we used a construct pCMV-AAV to demonstrate the benefits of miRNA target sequences in the new AAV helper. In pCMV-AAV2, the AAV cap gene is under the control of a CMV promoter. Another feature of pCMV-AAV2mir122 and pCMV-AAV2mir142 is the addition of four copies of mir122 sequences and mir142 sequences, respectively. We first demonstrated its function in tissue culture cells (Fig. 5). As expected, the expression of capsid protein in the 293 cell line was normal for all constructs tested. It is interesting that, in the HHL-5 cell line, which was derived from human liver cells (Pien et al., 2009), the expression from plasmid with mir122 was greatly reduced. As in vitro results may differ from what happens in vivo, we also tested them in vivo. All these plasmids were injected into the mouse liver through hydrodynamic injection. In pCMV-AAV2mir, it contained both mir122 and mir142 sequences. As shown by western blot using antibody recognizing AAV VP1, VP2, and VP3, liver-specific mir122 can considerably reduce/eliminate the capsid gene expression in mouse liver (Fig. 6). The reduction was even more remarkable than what was observed in the HHL-5 cells. In contrast, the controls showed that expression from cotransfected GFP plasmid remained unchanged. This study confirmed that the capsid expression can be reduced/eliminated by having miRNA targeting sequences incorporated into the rcAAV particles.

miRNA target cassettes regulated the expression of AAV2 capsid in vitro. pCMV-AAV2, pCMV-AAV2mir, pCMV-AAV2mir122, or pCMV-AAV2mir142 plasmids were transfected into HEK 293 and HHL5 cell lines. pAAV-CB-EGFP plasmid was also included in each transfection to monitor the transfection efficiency. Cells were harvested 72 hr post transfection, and cell lysates were prepared for western blot analysis (4–12% gradient polyacrylamide gel electrophoresis gel). AAV capsid was probed with a mouse anti-AAV capsid monoclonal antibody (American Research Products) (upper panel), and EGFP was probed with a mouse anti-EGFP monoclonal antibody (Invitrogen) (lower panel).
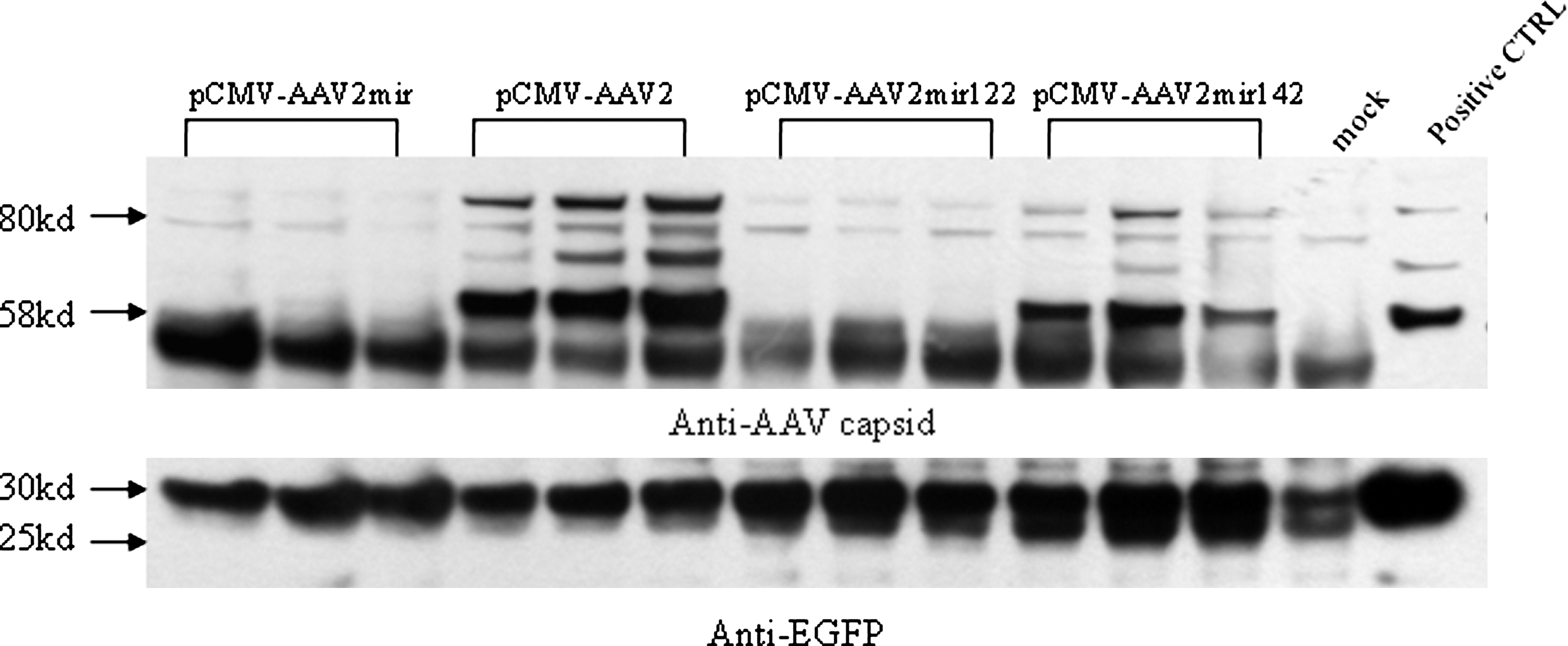
miRNA target cassettes regulated the expression of AAV2 capsid in vivo. The pCMV-AAV2, pCMV-AAV2mir, pCMV-AAV2mir122, or pCMV-AAV2mir142 plasmids were delivered to C57BL/J6 mice (n = 3 for each construct), along with pAAV-CB-EGFP plasmid as a transfection efficiency control, by hydrodynamic injection. Liver tissues were harvested 24 hr post transfection, and 50 μg of liver homogenate from each mouse was then prepared for western blot analysis (4–12% gradient polyacrylamide gel electrophoresis gel). AAV capsid was probed with a mouse anti-AAV capsid monoclonal antibody (American Research Products) (upper panel), and EGFP was probed with a mouse anti-EGFP monoclonal antibody (Invitrogen) (lower panel).
Discussion
The rcAAV particle contamination could be a key factor not only affecting the safety of rAAV vectors, but also determining whether or not transgene expression can persist long-term (Manno et al., 2006; Murphy et al., 2008; Hauck et al., 2009; Li et al., 2009a; Mays and Wilson, 2009; Mays et al., 2009; Pien et al., 2009; Vandenberghe et al., 2009a,b). Many strategies, such as altering the helper sequences to minimize homology between the AAV ITR and the rep and cap gene sequences, can significantly reduce rcAAV generation through homologous recombination events. Nevertheless, nonhomologous recombination between the AAV helper gene and vector sequences can not be controlled, because all these DNA sequences have to be expressed in the nucleus of the host cells. Large quantities of AAV vector DNA from transfection coupled with AAV DNA replication create a large pool of DNA templates, which increases the chance of nonhomologous recombination (Ferrari et al., 1997; Liu et al., 2000; Qiao et al., 2002). Due to the possibility of nonhomologous recombination, it is currently impossible to completely eradicate rcAAV formation from the rAAV production system.
There is no doubt that rcAAV particles are undesirable contaminants in vector preparation, especially those to be used clinically in human beings. Several studies based on the outcome of a recent hemophilia B clinical trial using AAV vector targeting the liver suggested that AAV capsid proteins, from either the capsid itself or rcAAV particles, may have caused the destructive immune response/inflammation that eventually eliminated the therapeutic gene expression (Li et al., 2009a,b; Mays and Wilson, 2009; Pien et al., 2009). Therefore, development of new technology is necessary to control residual de novo capsid expression from rcAAV particles. Here we attempted a novel approach by targeting the rcAAV particles that arise from unavoidable nonhomologous recombination. The rationale is that rcAAV particles would not be harmful if the capsid gene remains silent. Therefore, we attempted to include hematopoietic and hepatic miRNA target sequences in the helper expression cassette. As it is located immediately after the stop codon of the capsid proteins and before the poly(A) site, it is an integral part of the AAV helper gene, which could exert its function in rcAAV particles that express gene products for the capsid proteins.
The selection of mir122 and mir142 is based on their specificity in two important tissues. Liver cells are our transgene expression target tissues. Cells of hematopoietic origin are likely regulators for immune responses. The choice of miRNA target sequences to be included in the AAV helper would depend on the ultimate target tissue. For example, an miRNA targeting sequence specific to the muscle should be used so that rcAAV in the muscle tissue would stay inactive.
The miRNA target sequences are potent regulators of their underlying (specific) gene expression. Nevertheless, inclusion of miRNA target sequences mir122 and mir142 in the AAV helper did not affect its functionality for rAAV production in 293 cell lines. As demonstrated in Figs. 2 –4, various aspects of the rAAV vector produced remained unaltered, including physical parameters such as buoyant density and capsid protein composition. This suggests that the miRNA target sequence is in an inert state during vector production. Supporting this conclusion, the vector produced behaved normally in the transduction of tissue culture cells and in the muscle and liver of mice.
The critical function of the miRNA target sequences is to suppress potential viral gene expression from rcAAV particles. Using an “exaggerated” model with high-level capsid expression, we demonstrated that capsid expression could be suppressed 3 to 4 logs (Figs. 5 and 6). Currently, rcAAV was generated at a frequency of 1 out of 1 × 109 or 1 × 1010 genomes titer. The additional suppression would allow an improved rcAAV level to an equivalent of 1 × 1013 or 1 × 1014. For a human subject receiving 1 × 1014 vg, the effects of rcAAV would become negligible.
The miRNA targeting sequences can be readily adopted by any of the current rAAV vector production systems by simply adding them to the end of the AAV helper expression cassette (Mohiuddin et al., 2005; Chen, 2008; Clément et al., 2009; Virag et al., 2009). For example, the rAAV production cell line, baculovirus, or herpes-based rAAV production system can conveniently add appropriate miRNA target sequencing. It will not incur additional production costs, but it reduces the potential hazardous effect of rcAAV particles. The overall effects of miRNA targeting sequences in the AAV helper expression cassette can be analogous to a “free” insurance policy. If there is no rcAAV particle contamination, they remain inactive. However, in the case of an rcAAV contamination crisis, they exert their effects and suppress would-be-disastrous AAV viral protein expression in vivo. The same principle and concept may be applied toward other DNA viral vector productions.
Footnotes
Acknowledgments
This work was supported by the National Institutes of Health (HL080789 and HL084381 to W.X.).
Author Disclosure Statement
All authors have nothing to disclose.