
Abstract
Gene therapy for motor neuron diseases requires efficient gene delivery to motor neurons (MNs) throughout the spinal cord and brainstem. The present study compared adeno-associated viral (AAV) vector serotypes 1, 6, 8, and 9 for spinal cord delivery in adult mice, by the intraparenchymal or intrathecal route of administration. Whereas intraparenchymal injections resulted in local transduction of the lumbar segment of the spinal cord, intrathecal injections led to a broader distribution, transducing cells along the sacral, lumbar, and lower thoracic spinal cord. Overall, AAV6 and AAV9 performed better than the other serotypes. Dramatic differences in cell-specific expression patterns could be observed when constructs bearing the chicken β-actin (Cba) versus cytomegalovirus (CMV) promoter were compared. In summary, intrathecal delivery of AAV6 or AAV9 vectors containing the CMV promoter yielded the strongest levels of biodistribution and MN transduction in the spinal cord.
Introduction
The characterization of multiple serotypes of AAV has further strengthened its use for gene therapy. Serotypes vary by few amino acids, slightly changing the make-up of the capsid and affecting binding and intracellular trafficking (Wu et al., 2006a,b). Therefore, each serotype has a natural tropism for a particular tissue and/or cell type, through a given route of administration. For example, AAV1 has been demonstrated to be superior to AAV2 and AAV5 for skeletal muscle transduction (Wu et al., 2006b), whereas AAV6 may be optimal for retrograde transport to motor neurons (MNs) after intramuscular injection (Towne et al., 2009, 2010). Several groups have investigated the use of AAV to target motor neurons in the spinal cord after intramuscular, intraneural, intraparenchymal, or intrathecal injections. Hollis and colleagues compared multiple serotypes to determine the best approach to target the spinal cord and motor system after intramuscular and intraneural injections, and concluded that AAV6 was most amenable to this approach (Hollis et al., 2008). By intraparenchymal injections, only AAV1, AAV2, and AAV5 have been directly compared, with AAV1 showing robust transduction of neurons (Burger et al., 2004). Although intrathecal administration of AAV serotypes has been demonstrated by several groups (Storek et al., 2006; Towne et al., 2009; Kao et al., 2010; Vulchanova et al., 2010), it has focused mainly on dorsal root ganglia (DRG), with limited information regarding MN transduction. In fact, the DRG is anatomically distinct as a small structure suspended in the cerebrospinal fluid (CSF), whereas motor neurons lie deep within the spinal cord. Although pain and neuropathy may benefit from DRG gene delivery, motor neuron diseases require cord penetration across the pia by either direct injection or transpial movement from the CSF space.
The present study attempted to determine the best combination of AAV serotype and route of administration for gene delivery to the spinal cord and particularly to motor neurons. We compared AAV1, AAV6, AAV8, and AAV9 by intraparenchymal and intrathecal administration to determine which serotype resulted in the highest transduction of the spinal cord. Vectors packaging green fluorescent protein (GFP) under the control of either the chicken β-actin (Cba) promoter or the cytomegalovirus (CMV) promoter were tested in parallel, to exclude promoter bias in the assessment of biodistribution and cell tropism. In conclusion, intraparenchymal delivery of AAV9 followed by AAV6 provided robust but localized transduction of cells in the spinal cord, whereas intrathecal delivery of either AAV6 or AAV9 allowed for the widest distribution throughout the spinal cord.
Materials and Methods
Vector production
Recombinant AAV vectors were produced by the University of North Carolina Vector Core (Chapel Hill, NC), using a triple-transfection method in HEK293 cells as previously described (Grieger et al., 2006). All recombinant vectors packaged self-complementary genomes with enhanced GFP. The Cba-GFP vector included the CMV enhancer, miniature-Cba promoter, truncated simian virus 40 (SV40) 16S intron, and bovine growth hormone poly(A). The CMV-GFP vector included the CMV enhancer, CMV promoter, and SV40 poly(A). Highly pure recombinant virus containing self-complementary genomes was recovered using two sequential CsCl gradients, and then the peak fractions were dialyzed in phosphate-buffered saline containing 5%
Animals
Mice were handled according to Emory University (Atlanta, GA) Institutional Animal Care and Use Committee-approved protocols. To conduct intraparenchymal injections, an incision was made directly above the lumbar spinal cord through the skin and overlying muscles. Lidocaine (1%) was injected into the surrounding muscles. Paraspinous muscles were retracted and a laminectomy was performed at lumbar vertebra 2 (L2) to expose the underlying dura. The dura was slit and a glass nanopipette connected to a Nanoject oocyte injector (Drummond Scientific, Broomall, PA) was used to deliver the vector. A total of 6.25 × 108 vector genomes (VG) of each serotype in a 2-μl volume was delivered over an 8-min period (n = 5 per group). Removal of the pipette was delayed for 5 min to minimize reflux. The overlying muscle and skin were sutured. For intrathecal injections, the same procedure was performed. A laminectomy was performed, exposing the underlying dural membrane. A Hamilton syringe was slipped under the dura and the vector was slowly released into the CSF. A total of 2.5 × 109 VG of each vector was delivered in a 10-μl volume (n = 5 per group). An operating microscope was used to ensure that the needle was under the dura, but did not penetrate the spinal cord. The syringe was removed after 1 min and Gelfoam was used to cover the puncture in the dura to minimize CSF and vector leakage. The overlying skin and muscles were sutured. For all surgeries, mice were anesthetized with 2% isoflurane and posttreated with subcutaneous buprenorphine (0.05 mg/kg) for pain.
Vectors could have been titer-matched for either intraparenchymal or intrathecal deliveries. Because these routes of administration allow for different volumes, we decided to use the maximal volume and titer possible for each route of administration.
Tissue processing
Mice were killed and perfused 4 weeks postoperatively with 0.9% saline followed by 4% paraformaldehyde. Spinal cords, brains, and brainstems were dissected and postfixed overnight in 4% paraformaldehyde (PFA). Tissue was then cryoprotected in 30% sucrose and segments of lumbar, thoracic, and cervical spinal cords were frozen in Tissue-Tek O.C.T. compound (Sakura Finetek USA, Torrance, CA) at −80°C. Brains and brainstems were flash-frozen in 2-methylbutane and then stored at −80°C.
Histology
Repeated series of six 40-μm sections were cut with a cryostat (Jung Frigocut 2800N; Cambridge Instruments, Germany) and mounted on slides. For GFP analysis, tissue was mounted and coverslipped with VECTASHIELD with 4′,6 diamidino-2-phenylindole (DAPI) (Vector Laboratories, Burlingame, CA). For motor neuron staining, adjacent sections were mounted, blocked, and then incubated with either fluorescent Nissl (diluted 1:100; Invitrogen, Carlsbad, CA) for 2 hr at room temperature or anti-choline acetyltransferase (ChAT) (diluted 1:100; Millipore, Bedford, MA) polyclonal antibody overnight at 37°C. For glial cell staining, an anti-glial fibrillary acidic protein (GFAP) (diluted 1:1000; Dako, Carpinteria, CA) polyclonal antibody was used. The next day, slides were rinsed and incubated with a Texas red secondary antibody (diluted 1:100; Jackson ImmunoResearch Laboratories, West Grove, PA) for 2 hr at room temperature. Last, slides were rinsed and coverslipped with VECTASHIELD (Vector Laboratories). Control immunohistochemical analysis of untreated slides and with the secondary antibodies alone was also performed. Last, for assessment of inflammatory response, hematoxylin and eosin staining was performed in adjacent sections. Images were obtained with a Nikon DS-Fil color digital camera on a Nikon E400 microscope and quantified with NIS-Elements software (Nikon Instruments, Melville, NY).
Quantification and statistical analysis
For biodistribution, pictures were taken at low magnification. The longitudinal diffusion of GFP was determined by counting the number of sections (in one of each series of six 40-μm sections) that had GFP expression and then multiplying them by 240 μm (i.e., 6 sections × 40 μm per section). This estimate was used to compare the serotype with the greatest diffusion and it was calculated for each serotype, route of administration, and spinal cord segment (lumbar, thoracic, and cervical).
Motor neuron counts were assessed by counting the total number of ChAT+/GFP+ cells in the ventral horn in every sixth section. The total longitudinal biodistribution and number of transduced MNs was averaged for each group (n = 5 per group). One-way or two-way analysis of variance and post hoc Tukey tests were used to assess for significant differences in each category and among groups.
Results
Spinal cord biodistribution
Intraparenchymal injections
Lumbar injections of AAV serotypes 1, 6, 8, and 9 containing the Cba promoter resulted in GFP expression in the lumbar spinal cord 4 weeks after injection. Gene expression was localized predominantly at the injection site with limited diffusion, mostly through the ventral white matter. AAV9.Cba followed by AAV6.Cba resulted in the highest diffusion from the injection site compared with AAV1 and AAV8 (*p < 0.05) (Fig. 1). Although the mean spread of AAV9.Cba-mediated gene expression was greater than that of AAV6.Cba, the values were not statistically significant.

Spinal cord GFP biodistribution after intraparenchymal injection of AAV serotypes 1, 6, 8, and 9 containing the chicken β-actin (Cba) promoter. Vectors were injected into the lumbar spinal cord. Tissue was collected and analyzed by fluorescence microscopy 4 weeks after injection. Longitudinal diffusion of GFP expression was assessed. AAV9.Cba resulted in better diffusion away from the injection site than AAV1, AAV6, and AAV8 (*p < 0.05).
Intrathecal injections: Cba promoter
Although intraparenchymal injections result in robust transduction of targeted segments and may provide a means to treat symptoms specific to limited critical spinal cord segments such as diaphragm weakness, gait, or arm function, it is not surgically possible to treat the whole spinal cord in this fashion. To investigate diffuse application of AAV across a larger portion of the spinal cord, intrathecal injections of AAV9.Cba and AAV6.Cba were performed next. Although injected into the CSF, AAV.Cba gene expression remained restricted to the sacral, lumbar, and lower thoracic regions of the spinal cord. AAV6.Cba resulted in the highest levels of GFP expression, but limited penetration into the ventral horn. AAV6.Cba also resulted in robust transduction of ependymal cells of the central canal (Supplementary Fig. S1A; supplementary data are available online at
Intrathecal injections: CMV promoter
To exclude possible promoter bias in characterizing vector diffusion, AAV6 and AAV9 vectors packaging GFP under CMV promoter control were tested in parallel by intrathecal injections at the same doses. Both AAV6.CMV and AAV9.CMV resulted in high gene expression throughout the spinal cord. AAV6.CMV yielded robust gene expression in both gray and white matter in the lower thoracic, lumbar, and sacral spinal cord. GFP expression of AAV6.CMV in the cervical cord was limited to axons, not labeling any cell bodies. In particular, white matter staining was seen in Lissauer's tract (Supplementary Fig. S1B), which contains fibers projecting from dorsal root ganglion sensory neurons into the dorsal horn suggesting that gene delivery to sensory systems occurs over a broader distribution than to motor systems. AAV9.CMV resulted in robust gene expression in the lower thoracic, lumbar, and sacral spinal cord. GFP expression of AAV9.CMV in the cervical cord was also limited to axons, once again predominantly in Lissauer's tracts. Overall, intrathecal injection of AAV6.CMV resulted in the best diffusion throughout the cord, with GFP expression reaching the upper thoracic and cervical regions as opposed to the Cba promoter (*p < 0.05) (Fig. 2).
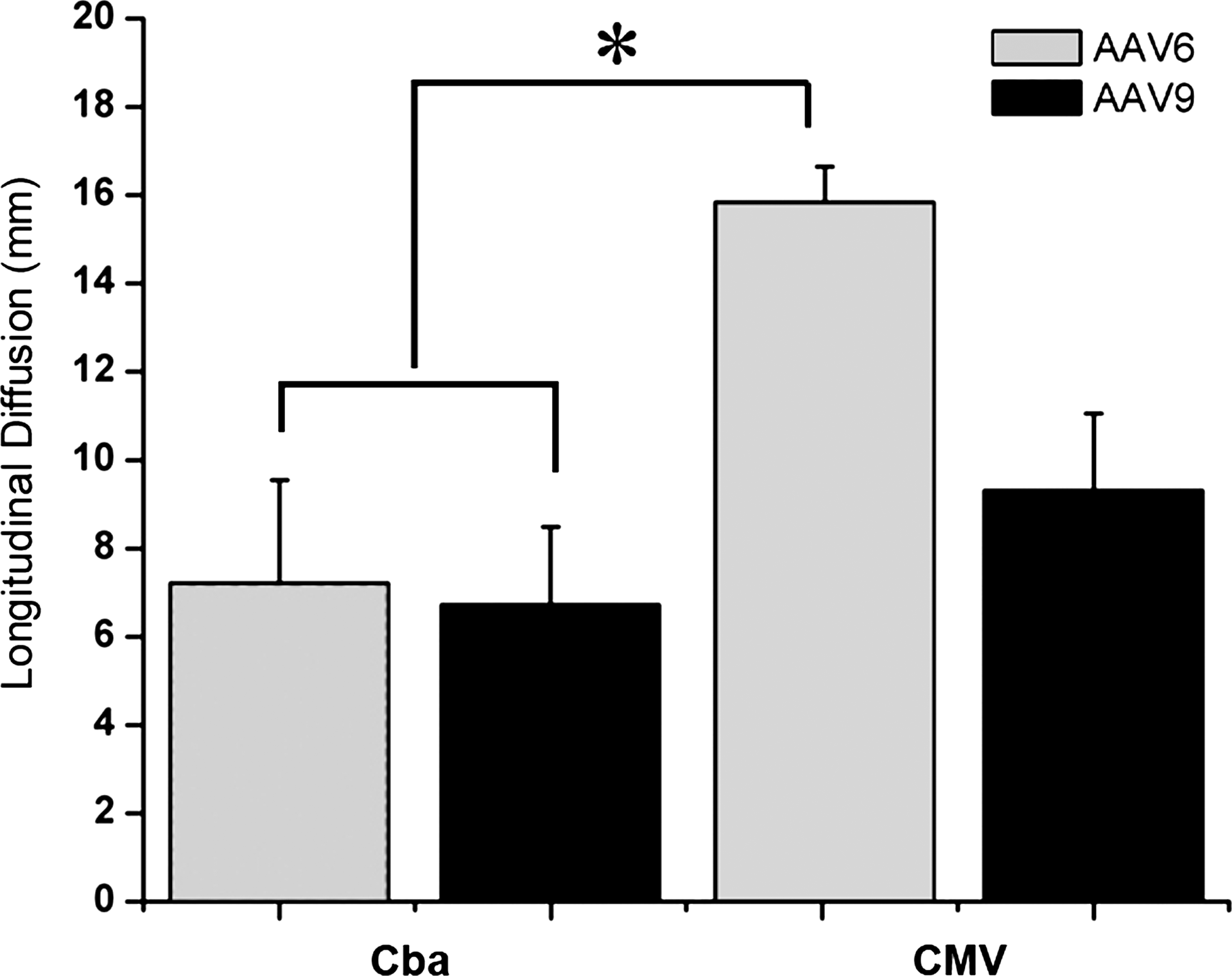
Spinal cord GFP biodistribution after intrathecal injection of AAV serotype 6 or 9 containing either the Cba or CMV promoter. Vectors were injected into the CSF at the level of the lumbar spinal cord. Tissue was collected and analyzed by fluorescence microscopy 4 weeks after injection. Longitudinal diffusion of GFP expression was assessed. AAV6.CMV and AAV9.CMV yielded robust and widespread GFP expression throughout the spinal cord in comparison with vectors containing the Cba promoter. The two-way ANOVA test revealed a significant effect of the promoter (*p < 0.05).
Motor Neuron Transduction
Intrathecal injections: Cba promoter
Interestingly, our assessment of MN transduction was quite different depending on the promoter used. Whereas AAV6.Cba resulted in a relatively small amount of MN transduction, independent of the route of administration, AAV9.Cba failed to transduce MNs (*p < 0.001) (Fig. 3).

Motor neuron (MN) counts after intrathecal injection of AAV serotype 6 or 9 containing either the Cba or CMV promoter. GFP-positive motor neurons were counted in the ventral horn in every sixth section. The total longitudinal number of transduced MNs was then averaged for each group. AAV9.CMV and AAV6.CMV yielded robust transduction of MNs in the ventral horn of the spinal cord. AAV9.Cba did not transduce MNs. The two-way ANOVA test revealed a significant effect of the promoter (*p < 0.05).
Intrathecal injections: CMV promoter
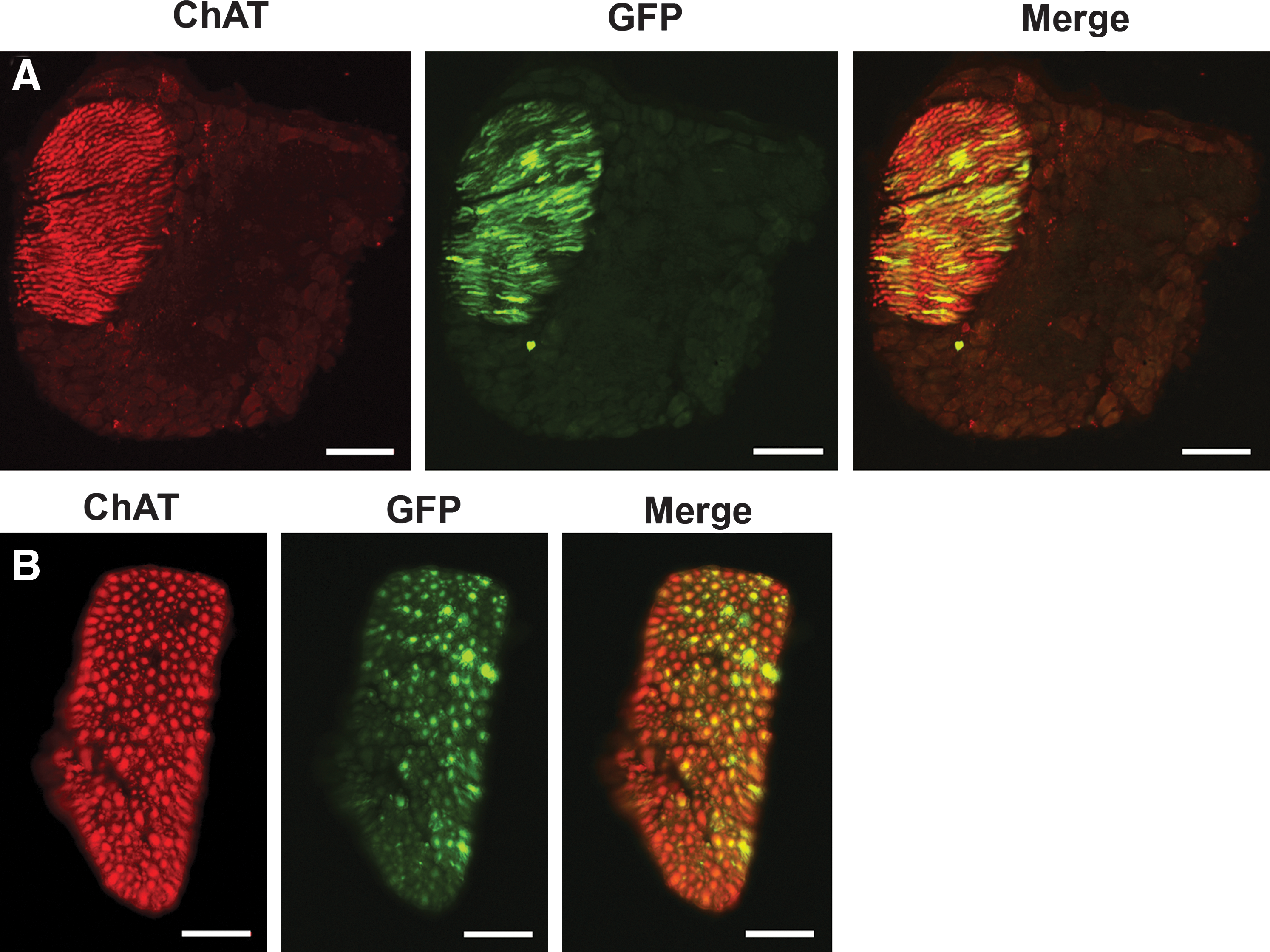
The CMV promoter revealed extensive and highly efficient transduction of MNs with either AAV6 or AAV9 when compared with the Cba promoter (*p < 0.05) (Fig. 3). AAV9.CMV resulted in the most efficient transduction of MNs, with up to 1200 transduced MNs observed in the spinal cord of a single animal. GFP expression colabeled with ChAT+ cells in the ventral horn (Fig. 4). No transduced cell bodies were found in the brain or brainstem, suggesting that AAV6 and AAV9 did not undergo retrograde transport to the brain. However, both AAV6.CMV and AAV9.CMV transduced cell bodies in the spinal cord, whose projections terminate in the brain, particularly in the spinocerebellar tract (Supplementary Fig. S1C). Last, both vectors but mostly AAV9.CMV yielded robust gene expression in motor axons in the lumbar spinal cord (Fig. 5).

MN transduction after intrathecal injection of AAV serotype 6 or 9 containing either the Cba or CMV promoter. Fluorescence microscopy shows GFP-positive motor neurons (choline acetyltransferase, ChAT) or astrocytes (glial fibrillary acidic protein, GFAP). Merged images show colocalization of GFP and ChAT in motor neurons or GFP and GFAP in astrocytes. (
Robust GFP expression in motor axons of the lumbar spinal cord after AAV9.CMV intrathecal injections. (
In summary, AAV6 and AAV9 bearing the CMV promoter proved highly conducive to spinal cord diffusion by intrathecal injection, with AAV9.CMV showing the most efficient MN transduction. Hematoxylin and eosin staining did not reveal any inflammatory reaction (data not shown), corroborating the advantageous features of AAV use in the CNS.
Discussion
The goal of this study was to determine the best combination of AAV serotype and route of administration for gene delivery to the spinal cord and motor neurons. We first compared AAV serotypes 1, 6, 8, and 9 after intraparenchymal injections. In summary, although restricted to the lumbar cord, AAV9.Cba and AAV6.Cba resulted in the best diffusion (Fig. 1). Because intrathecal injections are less invasive and may allow for diffuse transduction, in the next step we compared intrathecal injections, using the vectors that performed best after intraparenchymal injections (AAV6 and AAV9). In this series, we also compared vectors containing the Cba versus the CMV promoter. Intrathecal injections can potentially be performed in an outpatient setting and have substantially lower morbidity in comparison with intraparenchymal injections. Intrathecal injections of vectors containing the CMV promoter resulted in significantly better gene expression and higher transduction of motor neurons than those with the Cba promoter (Fig. 2). Overall, AAV9.CMV yielded the highest number of GFP-positive MNs (Figs. 3 and 4) with some overlap in the pattern of transduction of AAV6.CMV.
Although intrathecal injections were used successfully in this study, it is unlikely that we bathed the entire spinal cord and brain with AAV. In this study, a small, single dose of vector was injected into the CSF at the level of the lumbar spinal cord. Because of the small size of the space and the increased risk of reflux with increased volumes, only 10 μl of vector could be injected. It has been demonstrated that even in large animals, large amounts of drug infused slowly into the CSF, resulting in peak drug levels at the level of the injection that tapered off with distance from the injection site (Flack and Bernards, 2010). Therefore, it is likely that the vectors did not diffuse and circulate throughout the CSF; this is consistent with our data. In the best cases, we found GFP expression only in cell bodies up to the lower thoracic cord. Similarly, the distribution of gene expression was not always axially symmetric, suggesting that vectors penetrated at specific pial entry points that are not necessarily evenly distributed. We did see axonal GFP in the cervical cord and limited brain regions (data not shown). However, the absence of GFP in cell bodies in these regions suggests that the transgene arrived by anterograde transport from DRGs (Lissauer's tract) and transduced spinal cord neurons at lower segments, including the spinocerebellar neurons. In all cases, there was a break in GFP expression in the thoracic cord. As with the drug in the referenced pig study (Flack and Bernards, 2010), it is likely that the vector follows the natural flow of the CSF, which is dependent on the position of the animal. That would make the vector settle to the ventral side of the canal, explaining the apparent diffusion into the ventral spinal cord in most cases. Because of the larger volume of CSF in larger animals, we can predict that intrathecal delivery may result in better biodistribution throughout the thoracic and cervical cords. A larger canal may also allow for the use of catheters in larger animals, facilitating the injection of multiple boluses at different segments and improving the overall distribution.
AAV9 was demonstrated by two distinct groups to penetrate the blood–brain barrier (BBB) after intravenous administration, suggesting that this route of administration might be used for transduction of the CNS (Duque et al., 2009; Foust et al., 2009). Whereas one of these studies demonstrated restricted transduction of astrocytes in adult rodents (Foust et al., 2009), the other study also reported transduction of neurons (Duque et al., 2009). Of note, two different promoters were used in these studies: the Cba promoter was used in the first study (Foust et al., 2009) and the CMV promoter was used in the latter (Duque et al., 2009). In our study, the CMV promoter dramatically increased the expression of AAV9-transduced MNs, corroborating the results of these intravenous studies. Intrathecal injections may be more invasive than intravenous injections. However, intrathecal delivery would not only require far less vector, with substantially reduced off-target expression, but would also reduce side effects and improve the feasibility of biomanufacturing. Moreover, intrathecal injection may reduce the exposure of the reticuloendothelial system (spleen and liver), hence reducing the potential for a deleterious inflammatory response.
When considering the translation of these approaches, the two promoters used in our study (Cba and CMV) have distinct limitations. Cba promoters led to less intense gene expression. On the other hand, despite its robust and targeted expression, the CMV promoter has been shown to lose expression in the CNS at longer time points (McCown et al., 1996; Klein et al., 1998; Paterna et al., 2000; Tenenbaum et al., 2004). Although the assessment of both promoters in parallel facilitated the goals of this study (i.e., to assess the biodistribution of different AAV serotypes), neither would be ideal for translation of a therapeutic approach. The results of this study highlight the importance of developing novel promoters for use with self-complementary AAV genomes, specifically small promoters capable of stable and robust expression in MNs.
Footnotes
Acknowledgment
This work was supported by Hannah's Hope Fund.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.