
Abstract
The effective entry of retroviruses into target cells depends on the presence of viral envelope (Env) proteins and cognate cellular receptors, such as the murine cationic amino acid transporter-1 (mCAT-1) for the ecotropic murine leukemia virus (MLV-E). Here, we examined whether human cells internalize MLV-E or other retroviral pseudotypes irrespective of the presence of a specific receptor. Using fluorescently tagged Gag to monitor viral internalization, and treating cells with chloroquine or bafilomycin A1, we show that endocytosis is the main pathway for productive transduction with ecotropic particles, but endocytosis of retroviral particles itself does not depend on a suitable receptor or Env. Nonspecific endosomal uptake and lysosomal degradation occurred with all “illegitimate” envelope–receptor combinations tested: MLV particles pseudotyped with the ecotropic envelope or measles virus H and F proteins as well as “ecotropic” or “bald” HIV-1 particles. Kinetic studies in cell lines and primary human T lymphocytes showed the persistence of Gag–GFP signals for more than 10 days after exposure to retroviral vector particles, even in the absence of a suitable receptor. Further studies testing the Gag-mediated transfer of protein or retroviral mRNA revealed that nonspecific endocytosis prevented the release of functional particle-associated proteins and nucleic acids into the cytosol. We conclude that receptor-targeted retroviral particles are unlikely to escape nonspecific cellular uptake unless appropriate protective principles are discovered. Conversely, as lysosomal degradation was found to inactivate mRNA and proteins embedded into retroviral particles, receptor targeting is a useful strategy for both transient and permanent cell modification by retrovirus-like particles.
Introduction
However, Env is not necessary for the initial binding of viral particles to cells: MLV-E and “bald” MLV particles lacking Env bind to human cells with similar frequency as MLV with the “human-infectious” amphotropic Env (Pizzato et al., 1999). Nonspecific binding also occurs with HIV-1 (Mondor et al., 1998; Sharma et al., 2000). Even incorporation of a single-chain antibody into ecotropic or amphotropic Env does not result in enhanced particle binding to cells expressing the corresponding antigen (Pizzato et al., 2001). Glycosaminoglycans play a role in nonspecific virus binding, possibly by accumulating viral particles at the cell surface (Jinno-Oue et al., 2001; Walker et al., 2002). Env may thus be less important in the initial binding, but functions mainly in the induction of fusion between cellular and viral membranes.
There are two major routes for internalization of viral particles: pH-independent direct fusion with the cellular plasma membrane or pH-dependent fusion of the viral membrane with an endosomal membrane after endocytosis and exposure to low pH. The latter can be blocked with drugs that prevent endosomal acidification, such as the weak base chloroquine (CQ) (Ginsburg, 1990) or the specific inhibitor of the vacuolar H+-ATPase, bafilomycin A1 (BFLA-1) (Yoshimori et al., 1991; Mercer et al., 2010). Generally well accepted examples of viruses using the pH-independent entry mechanism are herpes simplex virus 1 (Maurer et al., 2008) and measles virus (Yanagi et al., 2006), whereas vesicular stomatitis virus (VSV) (Superti et al., 1987) and influenza virus (Lakadamyali et al., 2004) are pH dependent and typically enter cells by endocytosis. The cell entry pathway of MLV-E is controversial. Some studies suggest a pH-dependent endocytic pathway (Andersen and Nexo, 1983; McClure et al., 1990; Kizhatil and Albritton, 1997; Katen et al., 2001), and immunoelectron microscopy showed vesicular transport (Risco et al., 1995). Others reported the fusion of ecotropic Env to occur independently of low pH at the plasma membrane (Ragheb and Anderson, 1994; Kizhatil and Albritton, 1997; Koch et al., 2009).
In the case of HIV-1, particles bound to the cell surface do not exclusively use their canonical pathway of pH-independent fusion with the plasma membrane (Stein et al., 1987; McClure et al., 1988, 1990) but can also be endocytosed. Although this alternative route was long supposed to be noninfectious, treatment of the target cells with substances that prevent acidification of endosomes/lysosomes significantly increased the infectivity of HIV-1 (Fredericksen et al., 2002; Wei et al., 2005). Others provided direct evidence for productive infection by HIV-1 by pH-independent endocytosis (Daecke et al., 2005; Miyauchi et al., 2009), indicating that HIV-1 particles can escape endosomes before damage by strong acidification. Endosomal uptake itself was shown to be independent of Env and its receptor (Marechal et al., 1998; Schaeffer et al., 2001).
Elucidating the underlying mechanism of retroviral entry is of particular importance because a variety of gammaretroviral or lentiviral pseudotypes are used in nonclinical research, clinical gene therapy, and vaccination (Boztug et al., 2010; Dalba et al., 2007; Cappelli and Aiuti, 2010; Hacein-Bey-Abina et al., 2010; Kang et al., 2010). Pseudotyping alters tropism by substituting the natural Env proteins with those from other enveloped viruses. The significance of the pseudotype for genetic cell modification has been emphasized by work showing that lentiviral particles pseudotyped with measles virus (MV) glycoproteins efficiently transduce resting B and T lymphocytes, which are largely refractory to transduction with lentiviral vectors equipped with VSV glycoprotein (VSV-G) (Frecha et al., 2008, 2009; Funke et al., 2008).
Detargeting (avoiding transduction of undesired target cells) also represents an important rationale for pseudotyping. For example, gammaretroviral and lentiviral particles pseudotyped with the murine ecotropic envelope are generally considered not to be capable of entering human cells. These particles thus represent a preferred choice for nonclinical research regarding biosafety aspects, especially when transmitting oncogenic sequences (Schambach et al., 2006b).
In the present study we used fluorescently labeled Gag (comprising the viral structural proteins) of MLV and HIV-1 to tag viral particles and tested whether ecotropic, VSV-G, or MV pseudotypes, or bald particles lacking Env, would be internalized by human cells. Viral particles were detected in the endosomal/lysosomal compartment irrespective of their pseudotype and the potential availability of suitable receptors. All subsequent steps of the life cycle, including those promoting the endosomal/lysosomal release of functional retroviral proteins and mRNA, required the presence of Env and its receptor. Our findings support the notion that fusion, and not binding or uptake, represents the critical step in the selective targeting of retroviral vector particles.
Materials and Methods
Cell lines and primary cells
Human fibroblast cell lines HT1080 and HT1080mCAT (HT1080 cells expressing the murine ecotropic receptor mCAT-1), human embryonic kidney cell line 293T, murine fibroblast cell line SC1, and Flp indicator cells (HT1080 or SC1 cells expressing the Flp reporter cassette; Voelkel et al., 2010) were cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented with glutamine (Biochrom, Berlin, Germany), 1 mM sodium pyruvate, 10% fetal calf serum (FCS), streptomycin (100 μg/ml), and penicillin (100 U/ml) (all from PAA, Marburg, Germany). Primary human leukocytes were purified from whole blood by Ficoll gradient centrifugation (Histopaque 1083; Sigma-Aldrich, Munich, Germany) and subsequent cultivated in lymphocyte medium (RPMI 1640 supplemented with 10% FCS, streptomycin [100 μg/ml], penicillin [100 U/ml], 1% glutamine, and human interleukin-2 [hIL-2, 100 U/ml; Proleukin]) and Meropenem supplemented with anti-hCD3 (clone OKT3; eBioscience, San Diego, CA) and anti-hCD28 (clone CD28.2; eBioscience)-coated beads (Dynabeads M-450 Epoxy; Invitrogen Dynal Biotech, Carlsbad, CA). The purity of cultured T lymphocytes was confirmed by fluorescence-activated cell-sorting (FACS) analysis for CD3.
Plasmids and retroviral vectors
The integrating retroviral vector expressing DsRed-Express (SF91.DsRed) was derived from SF91.GFP (Hildinger et al., 1999; Schambach et al., 2000). The integrating vector encoding human truncated CD34 (SF11.tCD34) has been described (Fehse et al., 2000). To construct a lentiviral DsRed-Express expression vector (pRRL.PPT.SF.DsRed.pre), we excised the DsRed cassette from pCMV.DsRed-Express as an AgeI/blunt fragment (NotI site treated with Klenow fragment) and cloned it into the AgeI/blunt (BsrGI site treated with Klenow fragment) sites of pRRL.PPT.SF.GFPpre (Schambach et al., 2006b). To design a lentiviral Gag-Pol construct with a green fluorescent protein (GFP) label at the C terminus of the matrix protein (pcDNA3.HIV.MA.GFP) we introduced the GFP protein coding sequence into the MA protein coding sequence of the lentiviral Gag-Pol expression construct pcDNA3.GP.4xCTE (Schambach et al., 2006a). To avoid interference with MA function, the GFP sequence was flanked by two glycine residues on both sides and introduced just upstream of the protease cleavage site (VSQNY//) separating MA and CA (viral capsid). This localization was earlier determined to be tolerant for GFP insertions in the full-length HIV-1 genome (Muller et al., 2004). The vector SF91.aPBS.GFP.pre for mRNA transfer of GFP was described (Galla et al., 2008), as was the construction of the modified MLV Gag-Pol carrying either GFP or Flp recombinase fused to the C terminus of MA (MA.GFP and MA.Prot.Flpo) (Voelkel et al., 2010).
Production of viral particles
Retroviral supernatants were produced with a three-plasmid split gammaretroviral packaging system (Schambach et al., 2006a, c) by transient transfection of 293T cells with 7 μg of MA.GFP (Gag-Pol construct), 5 μg of SF91.DsRed or SF11.tCD34 (vector), and 2 μg of ecotropic envelope (kindly provided by T. Kitamura, Tokyo, Japan) (Morita et al., 2000). For VSV-G-pseudotyped particles 1.5 μg of VSV-G was cotransfected instead of ecotropic Env; for particles pseudotyped with measles virus envelope 2 μg of H (HcΔ24+4A) and 2 μg of F (FcΔ30) (Funke et al., 2008) (kindly provided by S. Kneissl and C. Buchholz, Langen, Germany) were cotransfected. For production of retroviral particles that are capable of transferring Flp recombinase the following plasmids were cotransfected into 293T cells: 3.5 μg of MA.Prot.Flpo, 3.5 μg of wild-type Gag-Pol, 5 μg of SF11.tCD34, and 2 μg of ecotropic Env. Particles for retroviral mRNA transfer were produced as described (Galla et al., 2004, 2008) by cotransfection of 5 μg of SF91aPBS.GFP.pre, 7 μg of wild-type Gag-Pol, and 2 μg of ecotropic Env. HIV-1 vector particles were produced in a four-plasmid split packaging system (Schambach et al., 2006b) by cotransfecting 5 μg of pRRL.PPT.SF.DsRed.pre (vector), 12 μg of pcDNA3.HIV.MA.GFP (Gag-Pol), 5 μg of Rev, and 2 μg of ecotropic Env. For all “bald” particles, the transfection mix excluded Env. All supernatants (besides MA.Prot.Flpo particles) were concentrated by ultracentrifugation (Optima LE-80K; Beckman Coulter, Krefeld, Germany) at 12,300×g and 4°C overnight.
Cell transduction
The day before transduction 5×104 HT1080 or HT1080mCAT cells per well were seeded in 24-well plates. For Western blot analysis, 7×105 cells per well were seeded in 6-well plates. Medium was replaced with fresh medium and concentrated viral particles were added. Cells were incubated at 37°C until harvest and analysis. For experiments in Figs. 1C, 3B, 4A and B, and 7A transduction was enhanced with protamine sulfate (4 μg/ml) and centrifugation for 1 hr at 400×g and 37°C. To inhibit lysosomal functions, cells were preincubated in medium containing 15 μM chloroquine (25 μM CQ in the case of Western blots) (Sigma-Aldrich) or 1 μM BFLA-1 (Santa Cruz Biotechnology, Heidelberg, Germany) 1 hr before transduction, maintained during transduction until harvest. If indicated, viral particles bound to the surface of cells were inactivated before analysis as described previously (Katen et al., 2001). Medium±CQ or BFLA-1 was removed and cells were washed with ice-cold phosphate-buffered saline (PBS). Afterward, cells were incubated for 30 sec in ice-cold citric acid buffer (pH 3.0) consisting of 40 mM citric acid, 10 mM KCl, and 135 mM NaCl. The buffer was removed and cells were washed with warm PBS, harvested, and analyzed by flow cytometry. For the kinetic analysis of MA.GFP uptake in HT1080 and HT1080mCAT cells (Fig. 2A–C), cells were kept in medium containing protamine sulfate (4 μg/ml). Concentrated viral particles consisting of SF11.tCD34, MA.GFP, and ecotropic Env were added (multiplicity of infection [MOI] of 2, based on tCD34 expression). At the indicated time points, cells were washed twice with ice-cold PBS and harvested in cold PBS containing 2 mM EDTA. Afterward, the cells were analyzed by FACS. Expanded human T lymphocytes (2×105) were transduced with bald or VSV-G-pseudotyped MA.GFP MLV particles on RetroNectin-coated culture plates, harvested, and washed as described for HT1080 cells and analyzed by FACS at various time points. Before transduction, the purity of T cells was confirmed by allophycocyanin-conjugated human CD3 (CD3-APC) (BD Biosciences, Heidelberg, Germany) antibody staining.

Endocytosis of MLV-E in human cells is independent of mCAT-1 receptor expression.
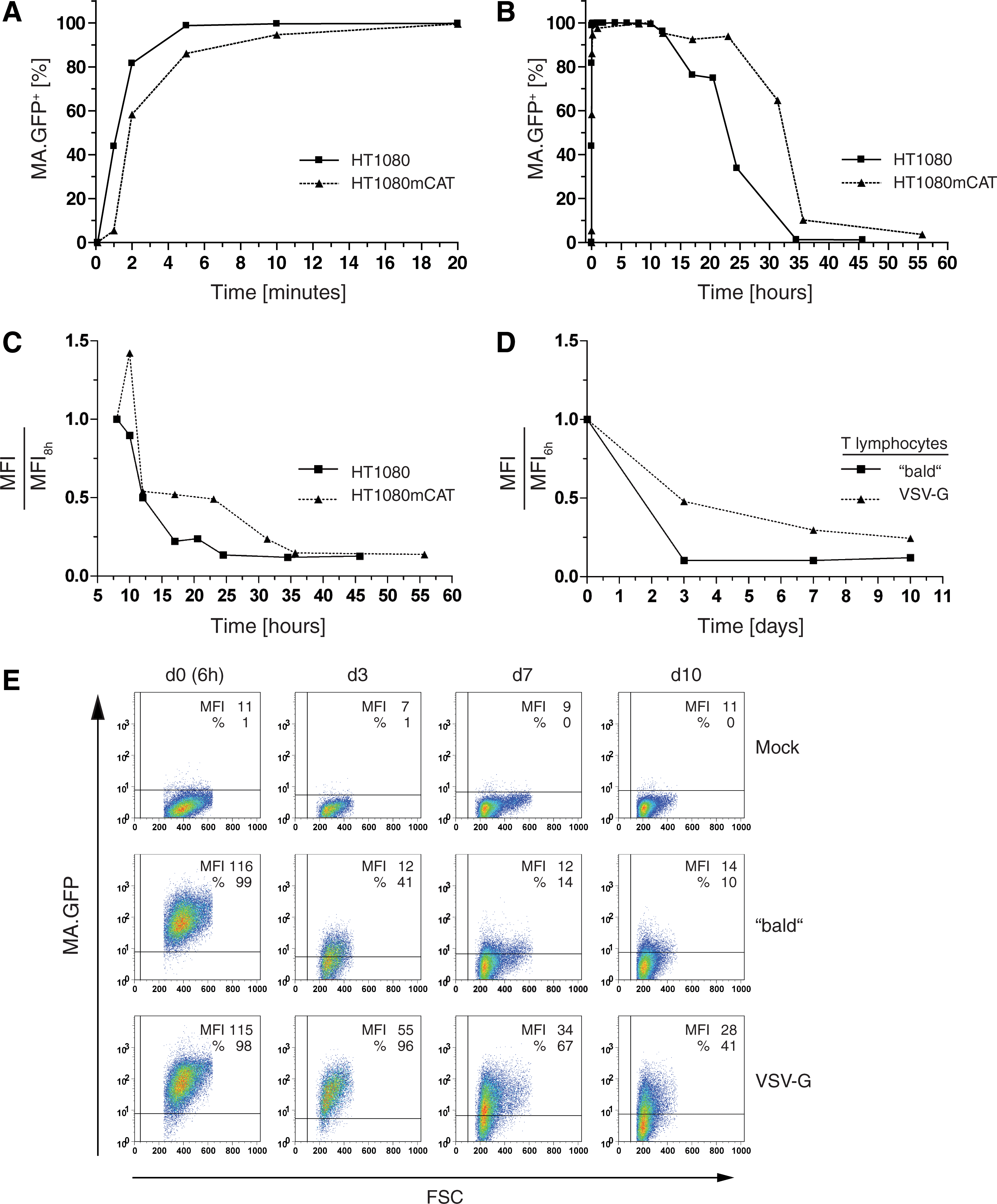
Kinetics of MA.GFP MLV particles in cell lines and primary human T lymphocytes.
Flow cytometry
Cells were harvested and measured on a FACSCalibur (BD Biosciences). GFP, DsRed, or dTomato fluorescence was directly measured. Truncated human CD34 was in some cases (Figs. 1C, 2A and B, and 3B) used for titer determination on SC1 cells and detected via an APC-conjugated antibody (anti-human CD34; BD Biosciences). After measurement of ≥10,000 events, data were analyzed with CellQuest (BD Biosciences) or FlowJo software (Tree Star, Ashland, OR).
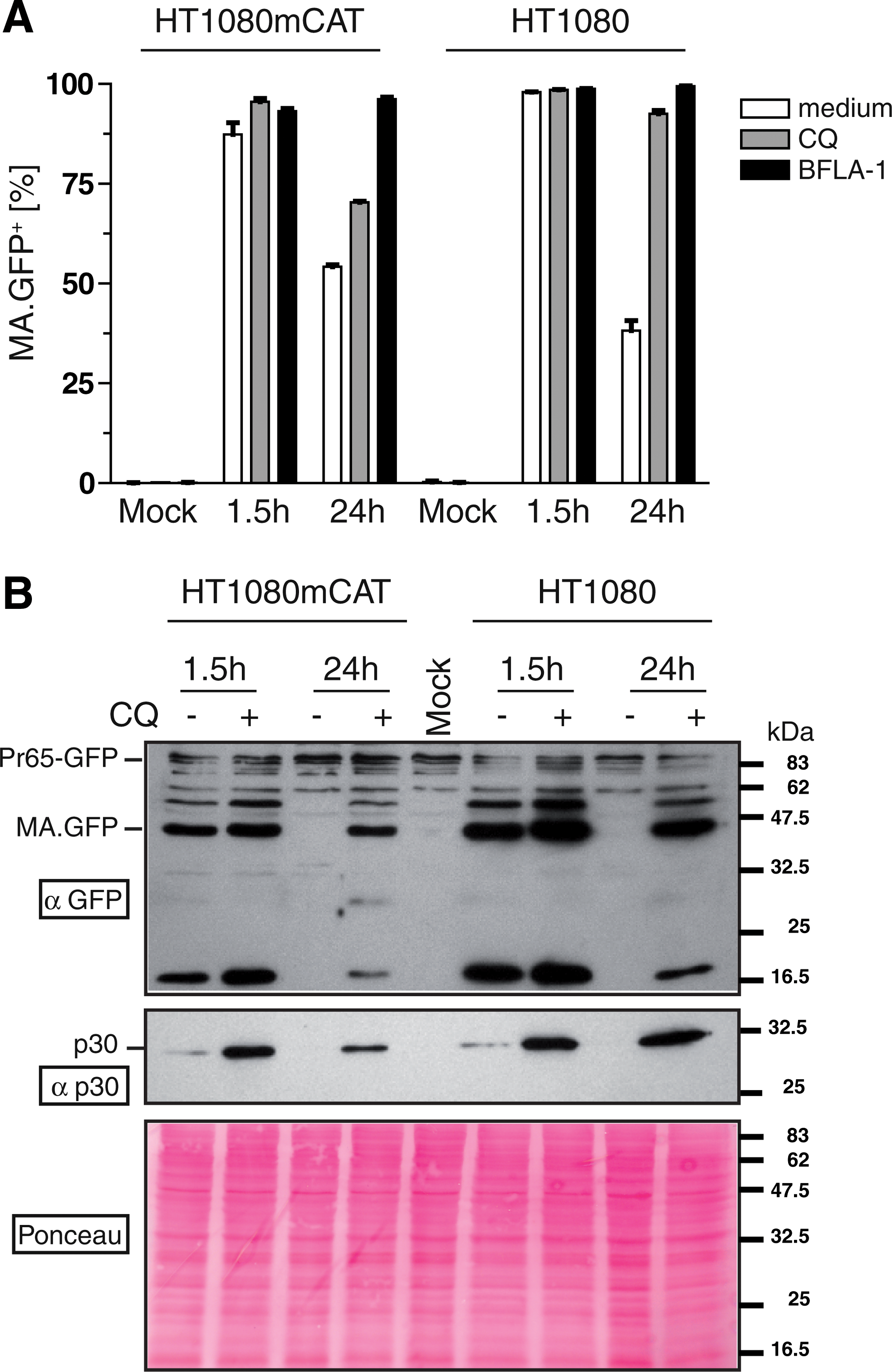
Human cells internalize “bald” MLV particles.
Immunoblots
Immunoblots were performed as described previously (Schambach et al., 2006a). In brief, transduced cells were washed twice in PBS and lysed in the presence of protease inhibitors. Fifty micrograms of denatured protein was separated by sodium dodecyl sulfate (SDS)–polyacrylamide gel electrophoresis and blotted to a nitrocellulose membrane (Whatman, Dassel, Germany). The membrane was stained with Ponceau S and afterward incubated with primary polyclonal antibody detecting GFP (Santa Cruz Biotechnology) in TBS (150 mM NaCl, 10 mM Tris-HCl; pH 8.0) −0.05% Tween. Detection by enhanced chemiluminescence (Thermo Fisher Scientific, Karlsruhe, Germany) was performed after incubation with a secondary peroxidase-conjugated antibody (Santa Cruz Biotechnology). The membranes were stripped in 62.5 mM Tris-HCl (pH 6.8), 2% SDS, 100 mM 2-mercaptoethanol and reprobed with the primary polyclonal antibody detecting p30 (anti-RLV p30 serum; kindly provided by S.K. Ruscetti, National Cancer Institute, Frederick, MD).
Fluorescence microscopy
To detect early endosome antigen-1 (EEA-1) and γ-adaptin (AP-1), 5×104 HT1080 and HT1080mCAT cells were grown on glass coverslips in 24-well plates. Viral particles containing SF11.tCD34, MA.GFP, and ecotropic Env (MOI of 2, based on tCD34 expression) were allowed to bind to the cells at 4°C for 1.5 hr. Medium was replaced with fresh medium at 37°C to start virus internalization. Protamine sulfate was present to enhance transduction. For the time points 4 min (EEA-1) and 3 min (AP-1), cells were incubated at 37°C for 4 or 3 min, respectively. For the time point 1.5 hr, cells were centrifuged for 1 hr at 400×g and 37°C and incubated for 30 min at 37°C. After incubation cells were washed three times in cold PBS, fixed with 4% paraformaldehyde in PBS, permeabilized with 0.1% Triton-X in TBS, and incubated in PBS with 3% bovine serum albumin (BSA) and primary antibody detecting either EEA-1 (mouse monoclonal anti-EEA-1 antibody; BD Biosciences) or AP-1 (20D6 hybridoma; Ahle et al., 1988). After washing with PBS, cells were incubated with rhodamine-labeled anti-mouse antibody (Invitrogen, Karlsruhe, Germany). Coverslips were mounted in HardSet mounting medium (Vector Laboratories, Burlingame, CA). Imaging was performed with a Zeiss LSM 510 meta confocal laser scanning microscope (Carl Zeiss, Oberkochen, Germany) equipped with a Plan-Apochromat 63× (numerical aperture, 1.4) oil immersion objective and the appropriate filter sets for GFP and rhodamine. Image analysis was performed with software Zeiss LSM Image Browser version 4.2.0.121 (Carl Zeiss). For colocalization experiments of bald and ecotropic MLV particles with LysoTracker red, 2×105 cells were grown in 35-mm glass bottom dishes (Fluorodish; World Precision Instruments, Berlin, Germany). After transduction with viral particles (14 μl of ecotropic or 23 μl of bald in 1 ml of medium without protamine sulfate) for the indicated times, medium was replaced with fresh medium containing 50 nM LysoTracker red DND-99 (Invitrogen) and cells were incubated for 1 hr at 37°C. Cells were washed with PBS and live cell imaging was performed with an Olympus FluoView 1000 confocal laser scanning microscope (Olympus, Hamburg, Germany) with a UPLSAPO 60× (numerical aperture, 1.35) oil immersion objective and the appropriate filter sets for GFP and LysoTracker red DND-99. Image analysis was performed with software Olympus FluoView version 2.0b (Olympus).
Results
Human cells endocytose MLV-E particles independently of mCAT-1
To test whether MLV-E can be internalized by human cells, we used fluorescently labeled Gag (Voelkel et al., 2010) to monitor retroviral uptake. Fusing GFP to the C terminus of the viral structural protein MA results in functional fluorescent retroviral particles (construct MA.GFP with GFP embedded into an intact gag-pol reading frame; Voelkel et al., 2010). Using a three-plasmid split gammaretroviral packaging system (Schambach et al., 2006a, c), we cotransfected plasmids encoding MA.GFP (Voelkel et al., 2010) and optionally the ecotropic envelope with an integrating retroviral vector expressing the red fluorescent protein DsRed (SF91.DsRed). The latter was used to monitor vector-mediated transgene expression indicating effective nuclear transfer with subsequent genomic integration of the vector genome. Concentrated viral supernatants were prepared to transduce human HT1080 fibroblasts. Because MLV-E can infect human cells expressing the ecotropic receptor mCAT-1 (Albritton et al., 1989), we used HT1080 cells engineered to express mCAT-1 (HT1080mCAT) as a positive control. Mock-treated cells served as a negative control. As expected, 24 hr after transduction, only HT1080mCAT cells expressed DsRed as detected by flow cytometry (Fig. 1A). When the cells were preincubated for 1 hr with CQ (15 μM) or BFLA-1 (1 μM) and transduced in drug-containing medium until harvest, integration was reduced (Fig. 1A), as also reported by others (Katen et al., 2001). These data suggest that an endocytic pathway was involved in infectious entry of MLV-E, which depended on the presence of Env and its receptor.
However, when examining MA.GFP fluorescence, we found positive cells 1.5 hr posttransduction irrespective of the presence of mCAT-1 (Fig. 1B). Whereas in HT1080mCAT cells the MA.GFP-derived fluorescence signal remained stable when examined after 24 hr, the signal in HT1080 cells lacking mCAT-1 declined to 50% unless CQ or BFLA-1 was present (Fig. 1B). On the basis of the mean fluorescence intensity of MA.GFP per cell, BFLA-1 seemed to be more efficient than CQ in preventing MA.GFP from degradation (data not shown). To exclude effects resulting from enhanced viral binding mediated by polycationic substances, the cells were transduced in the absence of protamine sulfate, and ice-cold citric acid buffer (pH 3) was used to remove surface-bound viral particles before flow cytometry (Katen et al., 2001).
In a similar experiment we confirmed these results by immunoblotting, detecting either GFP or viral capsid (CA, p30) protein in lysates of transduced HT1080mCAT or HT1080 cells harvested 1.5 and 24 hr after transduction (Fig. 1C). Results were consistent with flow cytometry because viral particles, detected as signals of both MA.GFP and p30, were present in the transduced cells early after transduction (1.5 hr) regardless of the presence of the receptor mCAT-1 (Fig. 1C). Again, the signals declined over time whereas they were recovered by addition of CQ.
These data suggest efficient endocytosis of MLV-E in human cells and furthermore provide evidence for a nonspecific internalization pathway functioning independently of the cognate receptor.
Kinetics of retroviral internalization in cell lines as well as primary human T lymphocytes
To further analyze the differences in particle uptake in the presence or absence of mCAT-1, we measured the kinetics in HT1080 as well as HT1080mCAT cells transduced with ecotropic MLV particles containing MA.GFP. We found the uptake to occur within minutes, having already plateaued 5–10 min after transduction and being slightly faster in the absence of mCAT-1 (Fig. 2A). Loss of the MA.GFP signal was also faster in the absence of mCAT-1, beginning to decline 12 hr posttransduction (Fig. 2B and C). These data indicate that there might be different pathways for uptake of MLV-E in cells depending on the availability of a suitable receptor.
To analyze the kinetics of MA.GFP MLV particles in primary human T lymphocytes in the presence or absence of a specific receptor, we generated viral supernatants as described previously, but omitting Env. The resulting “bald” MLV particles as well as VSV-G-pseudotyped MA.GFP MLV particles were concentrated and used to transduce primary human T lymphocytes. At early time points posttransduction, lymphocytes were washed in ice-cold citric acid buffer (pH 3) to remove surface-bound particles and analyzed by flow cytometry. Although the MA.GFP signal was lost faster when delivered from bald particles instead of from VSV-G-pseudotyped particles, almost all human T lymphocytes were positive for MA.GFP 6 hr posttransduction (Fig. 2D and E). These data not only confirm our findings in HT1080 and HT1080mCAT cells, but also demonstrate that the receptor-independent uptake of viral particles occurs in primary and therapeutically relevant human cells.
Endocytosis in human cells is independent of ecotropic Env
To further examine the role of ecotropic Env in cellular uptake, we used concentrated bald MA.GFP MLV particles for transduction of HT1080 and HT1080mCAT cells, as in Fig. 1A and B. Again, flow cytometry showed the majority of cells being positive for MA.GFP at 1.5 hr posttransduction (Fig. 3A). The MA.GFP signal mediated by bald particles declined by about half in both cells types (HT1080mCAT and HT1080) at 24 hr posttransduction (Fig. 3A), as for MLV-E in HT1080 cells lacking the cognate receptor (Fig. 1B). Again, endosome inhibitors prevented degradation of GFP fluorescence, with BFLA-1 being more efficient than CQ (Fig. 3A). In addition, the absence of DsRed-expressing cells (data not shown) suggested failure of viral fusion, reverse transcription, nuclear transport, or integration.
The immunoblot and flow cytometry data also correlated for bald particles (Fig. 3B): MA.GFP or p30 was detected in HT1080 or HT1080mCAT cells 1.5 hr after transduction and degraded within 24 hr, and the signal was rescued by CQ (Fig. 3B). These data show that ecotropic Env is dispensable for the endocytic uptake of MLV particles by human cells. However, nonspecific uptake efficiently inhibits the delivery of retroviral DNA.
MLV particles with different pseudotypes end up in lysosomes of human cells
We next investigated which cellular compartments may be involved in nonspecific/noninfectious endocytosis. On the basis of our kinetic studies (Fig. 2) we fixed the cells at 4 min or 1.5 hr posttransduction. Using antibodies against early endosome antigen-1 (EEA-1), a marker for early endosomes (Mu et al., 1995), and confocal microscopy, we detected no colocalization of MA.GFP with EEA-1 (which would be indicated by yellow overlay signals) at early (4 min; data not shown) and late (1.5 hr; Supplementary Fig. S1A [supplementary data are available online at
We repeated the experiment and labeled the cells at 3 min or 1.5 hr after transduction with an antibody directed against the clathrin adaptor complex AP-1, a marker for both the trans-Golgi network and endosomal subpopulations (Ghosh and Kornfeld, 2004). Similar to the previous experiment, we found no colocalization with MA.GFP at 3 min (data not shown) and 1.5 hr (Supplementary Fig. S1B).
After nonspecific endosomal uptake, the lysosome would be the final destination regardless of which type of vesicle was involved previously. Hence, we examined HT1080mCAT cells transduced with ecotropic or bald MLV particles carrying MA.GFP by confocal live-cell microscopy 4 and 7.5 hr posttransduction, using LysoTracker red DND-99 to stain acidic organelles such as late endosomes and lysosomes. The images were selected from z-stacks by the criterion of maximal colocalization (Fig. 4). Both particle types partially colocalized with the lysosomal compartment, where the bald particles were more enriched than the ecotropic particles (Fig. 4). In contrast, ecotropic particles apparently escaped this destiny in HT1080mCAT cells, consistent with the prolonged MA.GFP signal obtained in human cells expressing a cognate receptor (Fig. 2). It should be noted that appropriate mock-treated controls were used to rule out that photoconversion of LysoTracker red to LysoTracker green contributed to the observed colocalization signals (Freundt et al., 2007).
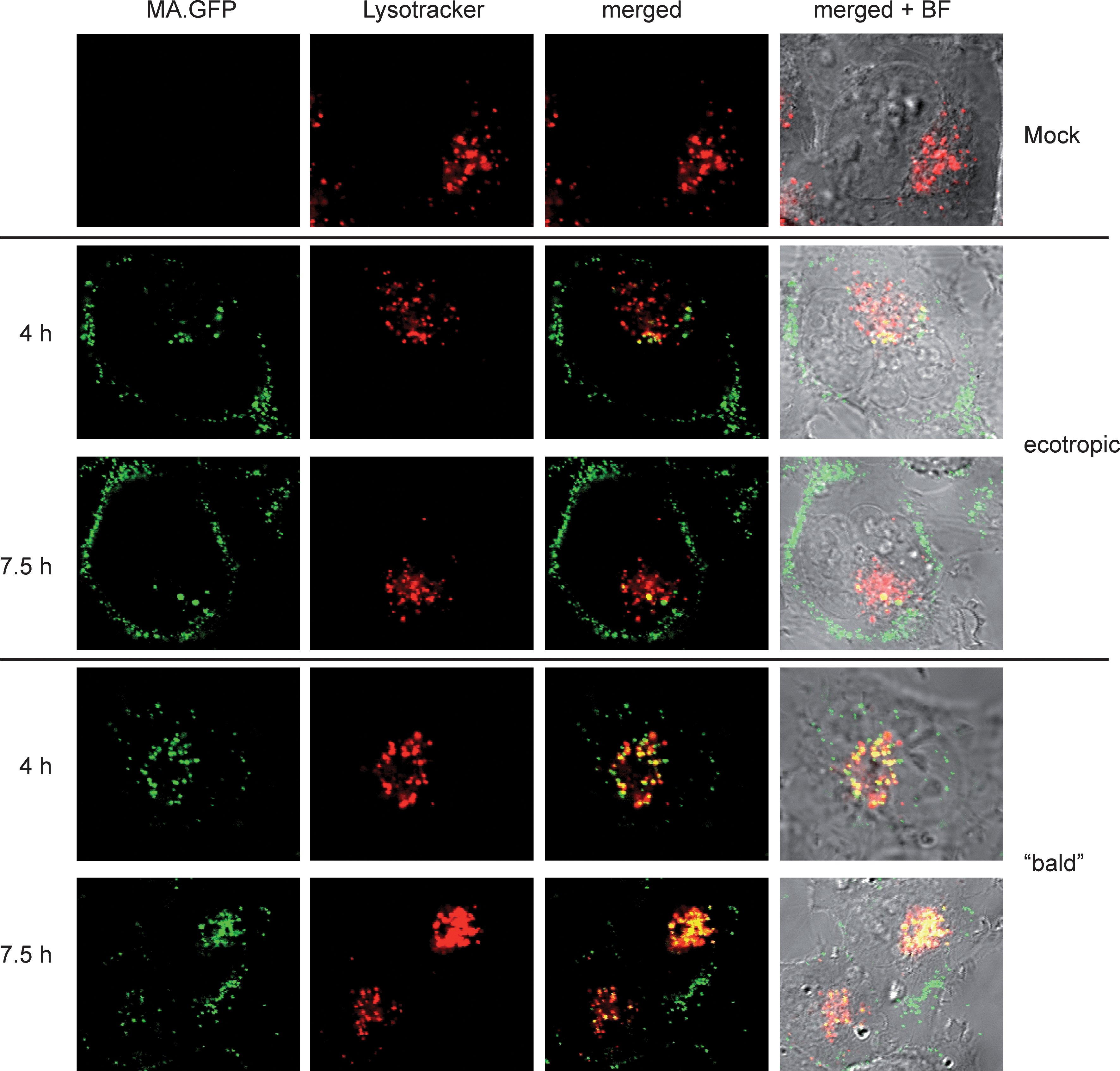
Ecotropic and “bald” MLV particles end up in lysosomes. HT1080mCAT cells were transduced with concentrated particles consisting of SF91.DsRed, MA.GFP, and either ecotropic (14 μl) or no envelope (bald, 23 μl) for 4 or 7.5 hr. Cells were stained for 1 hr with LysoTracker red DND-99 and live cell imaging was performed by confocal microscopy. Mock-treated cells served as controls. Green signals correspond to MA.GFP of viral particles, red signals to LysoTracker red DND-99, yellow signals to a colocalization of green MA.GFP and LysoTracker in merged images. BF, bright-field images. Images were selected from z-stacks by the criterion of maximal colocalization.
To characterize potential differences between the productive receptor-specific and nonspecific entry pathway, we performed a cell fractionation assay. After transduction of HT1080mCAT cells with either bald or ecotropic pseudotyped MA.GFP particles, we washed the cells with ice-cold citric acid buffer (pH 3) to remove surface-bound viral particles. Seven hours posttransduction we performed a sequential subcellular protein extraction and analyzed cytosolic as well as membrane/organelle protein fractions by immunoblot (Supplementary Fig. S2). Whereas we detected MA.GFP from ecotropic particles in the cytosol as well as in the organelle fraction, MA.GFP from bald particles was restricted to the organelle fraction. These data confirm that particles entering the cell in the absence of a receptor fail to be released into the cytosol.
We went on to examine the uptake of particles pseudotyped with measles virus hemagglutinin (H) and fusion (F) proteins, mediating pH-independent fusion with the plasma membrane, and particles pseudotyped with VSV-G for receptor-mediated endocytosis. First, we verified the specificity of the particles with H and F glycoproteins (vaccinia strain). Their receptors are CD46, a complement regulatory protein expressed on all nucleated human cells, and SLAM (CD150), expressed only on human leukocyte subsets (Yanagi et al., 2006). Therefore, H- and F-pseudotyped MLV would properly infect human HT1080 fibroblasts but not murine SC1 fibroblasts lacking human CD46 (Yanagi et al., 2006). We generated viral supernatants as described previously, by packaging MA.GFP and the integrating retroviral vector expressing DsRed but pseudotyped with H and F (HF). As expected, DsRed was detected only in transduced HT1080 cells, not in SC1 cells (Fig. 5A). In contrast, MA.GFP signals were also found in the murine SC1 cells (Fig. 5B). These data indicated that nonspecific and nonproductive endocytosis occurs also for HF pseudotypes.

MLV particles are nonspecifically internalized independent of pseudotype and are trapped in lysosomes.
Next, we analyzed whether particles containing MA.GFP pseudotyped with HF or VSV-G also ended up in lysosomes. As expected, HT1080mCAT cells transduced with VSV-G-pseudotyped particles colocalized with the late endosome/lysosomal compartment (7.5 hr posttransduction; Fig. 5C) and always showed some “agglutination” of MA.GFP signals. Interestingly, also for HF pseudotypes, significant colocalization was found (Fig. 5C).
Nonspecific endocytosis thus likely presents an additional route to the productive entry pathway, irrespective of the Env glycoprotein (ecotropic, VSV-G, or HF), for example, in the situation of receptor saturation or for degradation of particles lacking sufficient amounts of functional Env.
Endosomal trapping inhibits the release of functional retroviral mRNA or proteins
To define the stage at which the viral life cycle was blocked, we made use of our established system of modified Gag precursors (Voelkel et al., 2010). We produced MLV-E as well as bald particles containing a modified Gag-Pol in which Flp recombinase was fused to MA. Both proteins were separated by an additional protease site so that Flp should be cleaved from its viral fusion partner MA by the viral protease during particle maturation (construct MA.Prot.Flpo; Voelkel et al., 2010). In case of endosomal trapping, the Flp recombinase packaged within these retroviral particles would neither reach the cytosol nor the nucleoplasm, and thus would be unable to switch a chromosomally established reporter construct from GFP to dTomato expression (Voelkel et al., 2010). Indeed, when we transduced HT1080 cells containing the reporter construct with the ecotropic and bald Flp-transferring particles, there was no induction of dTomato expression (Fig. 6A). This occurred only in SC1 indicator cells in the presence of ecotropic Env and its receptor mCAT-1. In this case, treatment with CQ decreased Flp recombination (Fig. 6A).
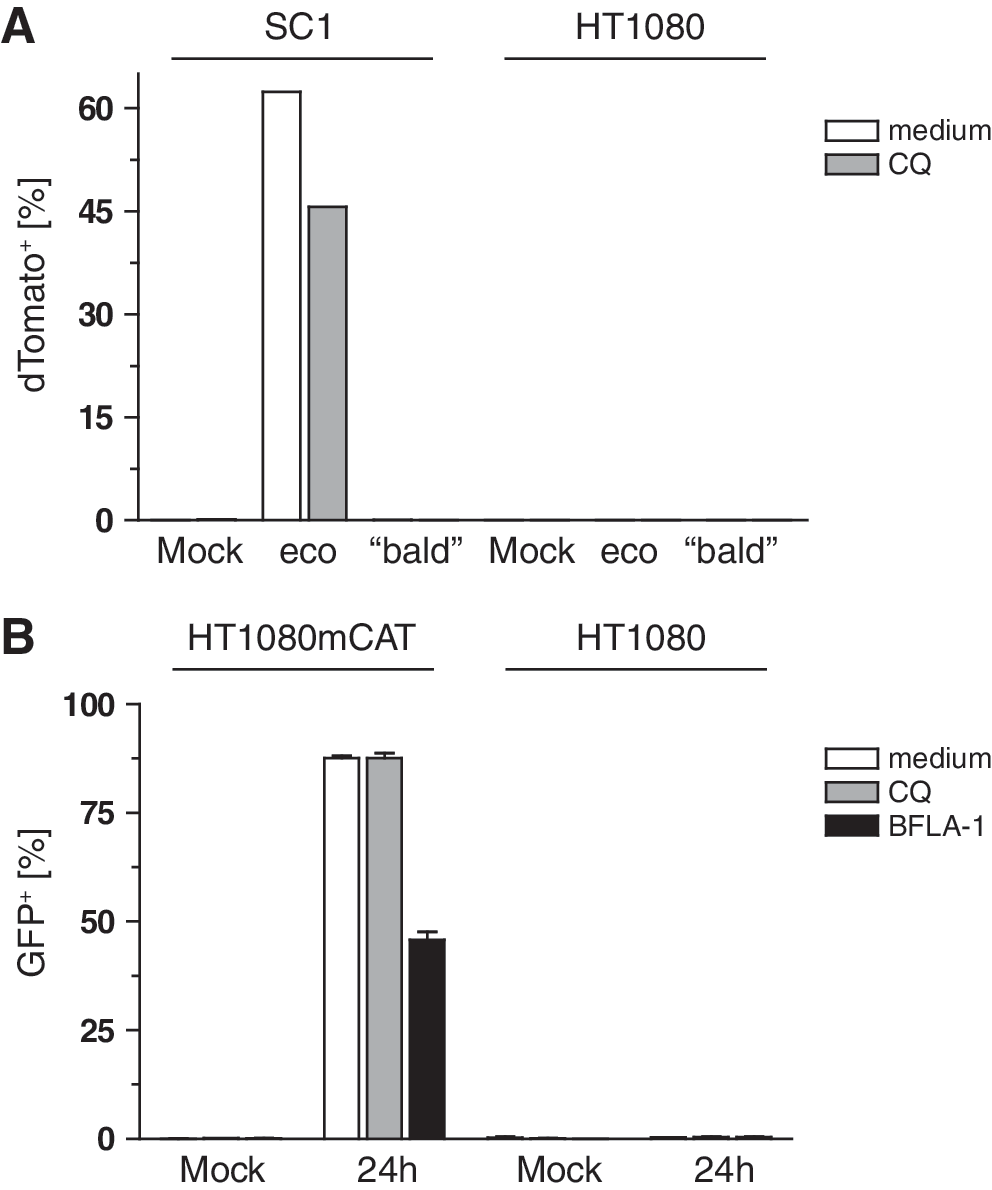
Flp-transferring MLV particles and retroviral mRNA transfer reveal the trapping within endosomes.
To monitor the fate of retroviral nucleic acids, we used our system of retroviral mRNA transfer (Galla et al., 2004): A mutated primer-binding site in a retroviral GFP vector blocks reverse transcription, generating a viral mRNA that serves as a translation template. GFP expression thus indicates cytosolic delivery of the viral mRNA. We packaged the vector expressing GFP (SF91.aPBS.GFP.pre) together with wild-type MLV Gag-Pol with or without the ecotropic Env and used these two particle types to transduce HT1080mCAT and HT1080 cells±CQ or BFLA-1 (analogous to the experiment in Fig. 1A and B). After 24 hr GFP was expressed only in HT1080mCAT cells (Fig. 6B), but not in HT1080 cells transduced with ecotropic particles (Fig. 6B) or in HT1080 and HT1080mCAT cells transduced with bald particles (data not shown). In HT1080mCAT cells transduced with the ecotropic particles, GFP expression was reduced by BFLA-1 treatment, indicating endosomal trafficking (Fig. 6B). On the basis of the mean fluorescence intensity of GFP per cell, a similar effect of CQ was detected (data not shown). These data indicate that endosomal/lysosomal trapping completely degrades retrovirus-like particles, including their codelivered proteins and nucleic acids.
Nonspecific endocytosis as an additional route used by lentiviral particles
Last, we were interested in whether nonspecific internalization may also occur for other retroviruses. We therefore investigated the uptake of ecotropic pseudotyped as well as bald HIV-1-based lentiviral particles in HT1080 and HT1080mCAT cells. We packaged an integrating lentiviral vector expressing DsRed (RRL.PPT.SF.DsRed.pre) together with lentiviral Gag-Pol carrying GFP fused to MA, Rev, and the ecotropic envelope. To produce bald lentiviral particles, we omitted Env. Transductions were performed as described for ecotropic and bald MLV (Fig. 1A and B, and Fig. 3A). Although fewer ecotropic lentiviral particles were taken up within 1.5 hr (Fig. 7A, MA.GFP+ cells: ∼70% in HT1080mCAT, ∼35% in HT1080) compared with ecotropic MLV (Fig. 1B, MA.GFP+ cells: ∼100% in both cell types), we observed the same principle: that is, uptake of ecotropic lentivirus occurred independently of the presence of mCAT-1, the MA.GFP signal declined over time, and CQ or BFLA-1 prevented degradation. The same was applicable to the bald HIV-1 particles (Fig. 7B). Again, integration of the vector genome with expression of DsRed depended on the presence of mCAT-1 (data not shown).
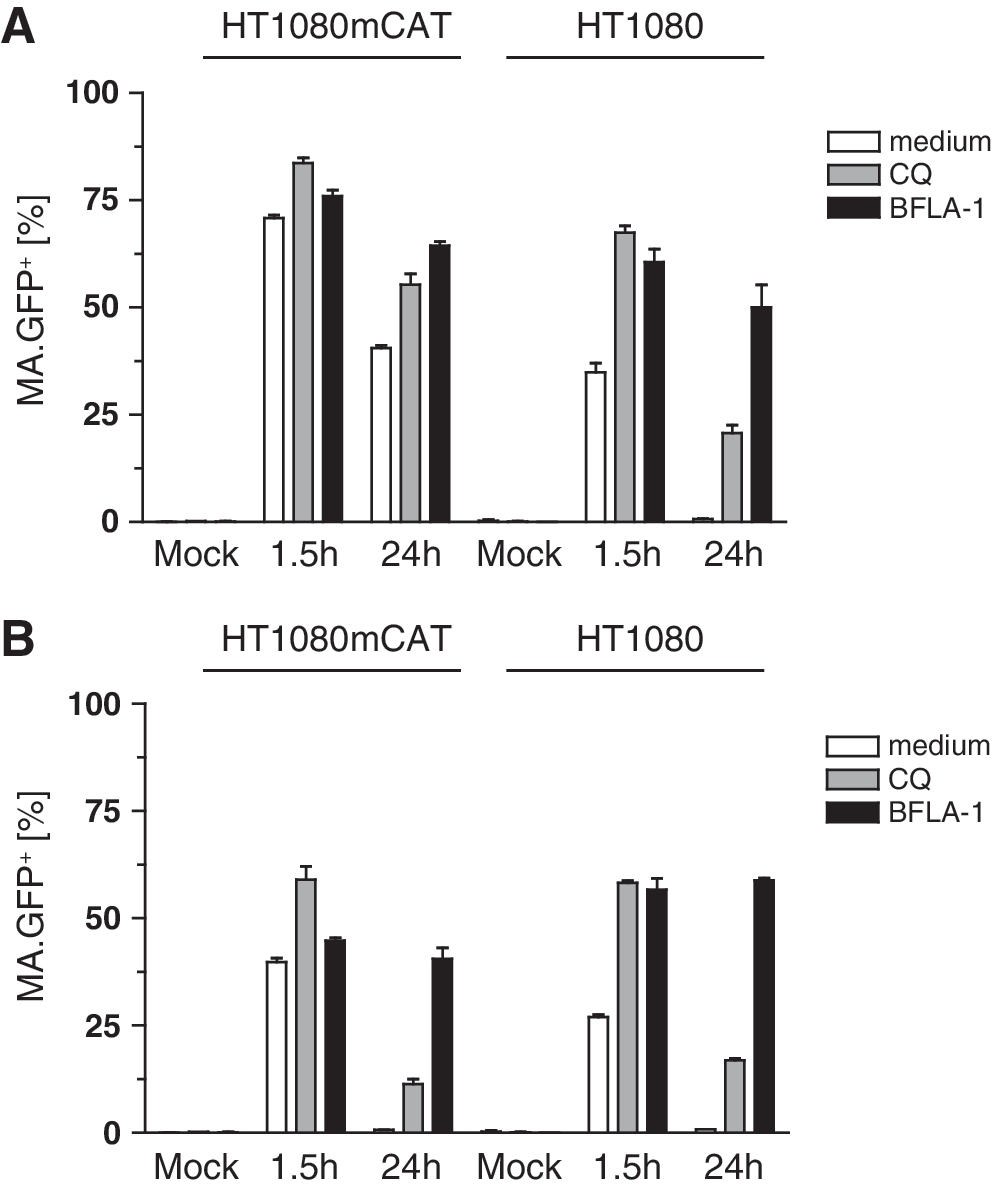
Nonspecific endocytosis of HIV-1 particles in human cells.
Discussion
It is commonly assumed that ecotropic MLV particles, and other gammaretroviral or lentiviral pseudotypes lacking suitable envelope proteins, cannot enter human cells. However, by using fluorescent MLV particles labeled with GFP fused to the viral MA protein we showed that human cells internalize MLV-E particles despite the absence of the receptor mCAT-1 or the authentic Env. The lysosomotropic weak base CQ as well as the vacuolar H+-ATPase inhibitor BFLA-1, which both block endocytic acidification, prevented degradation of MA.GFP-modified Gag, suggesting endocytosis as the underlying mechanism of particle uptake. However, despite delayed degradation in the presence of these drugs, endocytosed particles lacking an appropriate Env were trapped within vesicles. Furthermore, the integration of MLV-E vector genomes in human cells expressing mCAT-1 was inhibited by CQ or BFLA-1, suggesting that endocytosis with pH-dependent uptake, as proposed by others (Andersen and Nexo, 1983; McClure et al., 1990; Risco et al., 1995; Kizhatil and Albritton, 1997; Katen et al., 2001), rather than fusion with the plasma membrane (Ragheb and Anderson, 1994; Kizhatil and Albritton, 1997) is required for MLV-E cell entry.
Classical assays determining the susceptibility of human cells to MLV infection measure the integration of the viral DNA, typically using reporter genes (Albritton et al., 1993; Sharma et al., 1997). Our approach based on fluorescent MLV particles, made by fusing GFP to the viral MA protein combined with an integrating retroviral vector expressing another fluorescent marker, provides the opportunity to monitor early entry events independent of viral integration. Instead of GFP, the fusion of a bioactive nuclear protein such as Flp recombinase to MA (Voelkel et al., 2010) allowed us to draw conclusions on endosomal escape, as the delivered Flp recombinase is capable of catalyzing DNA recombination only if it escapes from endosomes. Endosomal escape was also detected by means of our established platform of retroviral mRNA transfer (Galla et al., 2004). We found the nonspecific endosomal trapping to inactivate mRNA and proteins that are codelivered with the retroviral particles. Receptor targeting thus allows stringent discrimination of target cell effects not only for the conventional use of retroviral vectors to deliver integrating DNA, but also for the use of retrovirus-like particles as carriers of foreign proteins (Voelkel et al., 2010) or mRNA (Galla et al., 2004). This may be of interest for applications in transient cell modification (Schott et al., 2011) or vaccination (Dalba et al., 2007).
By confocal laser scanning microscopy we were unable to reveal the localization of retroviral particles in early endosomes or the trans-Golgi network. However, as the contribution of early endosomes is only transient, the viral particles might have left these organelles at the analyzed time points, or the amounts of measurable MA.GFP might have been below the detection level. Fluid-phase markers or other markers for early endosomes such as Rab5 might provide further information on the involvement of early endosomes.
Caveola-mediated endocytosis could be considered to be involved in early trafficking events. Caveolin-1 (Cav-1), a marker protein of caveolae, was shown to colocalize with the N-terminus of MLV MA during viral assembly (Yu et al., 2006), and mCAT-1 was also found to be associated with Cav-1 (Lu and Silver, 2000). Although studies have provided evidence that caveolae might interact with late endosomes and lysosomes (Botos et al., 2008; Kiss and Botos, 2009), the physiological role of caveosomes has been questioned (Hayer et al., 2010; Mercer et al., 2010). Another potential mechanism for viral entry might be macropinocytosis, an endocytic mechanism used by many viruses (Mercer and Helenius, 2009). Interestingly, macropinosomes were found to become acidified (Hewlett et al., 1994), making macropinocytosis a candidate pathway for nonspecific retroviral uptake.
Whatever upstream route is taken, we found that nonspecifically ingested retroviral particles accumulated in late acidic endosomes/lysosomes.
Although nonspecific endocytosis does not support subsequent steps of the retroviral life cycle such as membrane fusion, release of particle-associated proteins or mRNA, reverse transcription, nuclear translocation, or chromosomal integration, it will nevertheless contribute to the outcome of experiments using various forms of retroviral vectors in basic research and clinical gene therapy. As we also found nonspecific internalization for MLV enveloped with measles virus H and F glycoproteins and “bald” lentiviral particles, nonspecific endocytosis seemed to occur independently of the particle pseudotype and capsid. Indeed, nonproductive internalization of HIV-1 carrying its natural Env was shown to occur in human renal cells (Mikulak and Singhal, 2010) and neural cells (Alvarez Losada et al., 2002) lacking the HIV-1 receptor CD4. Furthermore, nonproductive endocytosis of HIV-1 pseudotyped with natural Env seems to occur in parallel to the main route of HIV-1 infectious entry by plasma membrane fusion (Marechal et al., 1998; Schaeffer et al., 2001). When using vectors with targeting domains in the envelope proteins, such as the emerging measles virus pseudotypes and targeted envelopes derived thereof (Buchholz et al., 2009), nonspecific uptake may reduce the bioavailability of targeted particles and even trigger potential off-target effects, if the particles or their degradation products induce an innate or acquired immune response. This concern is corroborated by our observation that retrovirally delivered proteins, and potentially even infectious particles (Skinner et al., 2009) may persist for several days in cells, even after nonspecific uptake (Fig. 2).
Although many efforts have been made to produce targetable retroviral vectors by insertion of a targeting domain in Env, these attempts often failed to achieve high transduction levels (Benedict et al., 1999; Zhao et al., 1999; Lorimer and Lavictoire, 2000; Pizzato et al., 2001; Viejo-Borbolla et al., 2005). Various underlying reasons were discussed, including nonspecific binding to other cells (Pizzato et al., 2001), failure to induce membrane fusion (Benedict et al., 1999; Zhao et al., 1999; Viejo-Borbolla et al., 2005), and redirection to a noninfectious endosomal pathway (Viejo-Borbolla et al., 2005). Our study supports the latter mechanism as a relevant limitation.
To improve cell targeting, strategies preventing nonspecific ingestion should thus receive more attention. Furthermore, it might be important to determine whether there is a connection between envelope-independent uptake and nonspecific adsorption of viral particles to cells, reported previously (Mondor et al., 1998; Pizzato et al., 1999, 2001; Sharma et al., 2000; Walker et al., 2002). If nonspecific endocytosis depended on nonspecific adsorption, vectors lacking undesired adhesion properties might be developed, as exemplified for chemically modified adenoviral vectors (Kreppel et al., 2005). If the focus is to improve transduction efficiency rather than cell specificity, endosomolytic agents may be of interest, such as protein VI of adenovirus (Wiethoff et al., 2005) or the INF7 peptide of influenza hemagglutinin (Maeda et al., 1981).
In conclusion, our study reports nonspecific ingestion and lysosomal trapping as a major pathway for gammaretroviral and lentiviral particle uptake, largely independent of pseudotype. We showed that lysosomal trapping not only abrogates the default pathway of transgene integration but also inactivates proteins and mRNA embedded into retrovirus-like particles. Although receptor targeting holds great promise to exclude productive transduction of undesired cell types with retrovirus-embedded protein, mRNA, or reverse-transcribed DNA, prevention of nonspecific uptake should receive more attention to avoid residual off-target effects and nonspecific trapping, especially for direct administration in vivo.
Footnotes
Acknowledgments
The authors thank Ernst J. Ungewickell for useful discussions and for reading the manuscript. The authors are grateful to Rudolf Bauerfeind (Confocal Microscopy Facility, Hannover Medical School) for support with the confocal laser scanning microscopes. In addition, the authors thank Ivonne Fernandez, Diana Knetsch, and Valerie Mordhorst for technical assistance, and are grateful to Sabrina Kneissl and Christian Buchholz for providing the measles virus envelope proteins H and F. This work was supported by grants from the Deutsche Forschungsgemeinschaft (SFB738 and cluster of excellence REBIRTH [EXC 62/1]), the Bundesministerium für Bildung und Forschung (BMBF Joint Research Project IFB-Tx), and the European Union (FP7 project PERSIST, HEALTH-F5-2009-222878).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.