
Abstract
Interleukin-15 (IL-15) is known to stimulate the proliferation of CD8+ T-cells and natural killer cells, and also to help to maintain memory CD8+ T cells, suggesting that it may be of value in cytokine treatment of bladder cancer. In this experiment, we tested the efficiency of intravesical liposomal IL-15 gene delivery and its antitumor effect in a mouse orthotopic bladder cancer model. We established an orthotopic bladder cancer model by implanting 5×105 MBT-2 cells into female C3H/HeN mice through the urethra. The mice received repeated intravesical gene delivery injected with liposome-mediated plasmids (5 μg) transurethrally. On day 23, the bladder weights in the group receiving medium alone, the beta-galactosidase gene delivery control group, and the IL-15 gene therapy group were 196±36 mg, 201±35 mg, and 96±29 mg, respectively (p<0.05), demonstrating the antitumor effect of intravesical IL-15 gene therapy in this model. In the bladders treated with IL-15 gene plasmid instillation, histological analysis revealed that many inflammatory cells were induced around the tumors. Immunohistochemical analysis confirmed that there was predominant infiltration of CD8+ T cells around the tumor nest. After the intravesical IL-15 gene therapy, the growth of rechallenged subcutaneous MBT-2 cells in surviving mice was inhibited again via tumor-specific cytotoxic T lymphocytes, although newly implanted FM3A cells in the same mice were not rejected. The present findings indicate that IL-15 gene therapy may be a promising new adjuvant therapy for bladder cancer.
Introduction
To improve the efficiency of immunotherapy and to diminish its side effects, gene therapy that sufficiently activates only the targeted cytokine would be an attractive strategy for the management of non–muscle-invasive bladder cancer. Interleukin (IL)-15 is a novel 15-kDa cytokine that uses the β- and γ-chains of IL-2 receptors for signal transduction and shares many of the biological activities of IL-2 (Bamford et al., 1994; Giri et al., 1994; Grabstein et al., 1994; Waldmann et al., 2001), including stimulation of the proliferation of T and B lymphocytes and natural killer (NK) cells (Armitage et al., 1995; Carson et al., 1995; Mori et al., 1996; Colucci et al., 2003; Waldmann, 2006), and the facilitation of cytotoxic T-lymphocyte (CTL) and lymphokine-activated killer cells (Gamero et al., 1995; Lewko et al., 1995; Munger et al., 1995). These properties suggest that IL-15, like IL-2, may be of value in cancer control. Several reports have suggested IL-15 has antitumor effects against various cancers other than bladder tumors. A previous report demonstrated that IL-15 induced killer activity in peripheral blood mononuclear cells against lung cancer cell lines (Takeuchi et al., 1996). Another report showed that daily administration of IL-15 prolonged the period of remission induced by cyclophosphamide in mice bearing rhabdomyosarcoma (Evans et al., 1997). In addition, IL-15 was reported to play an important role in inducing the homeostatic proliferation and maintenance of memory CD8+ T cells, while IL-2 can promote apoptosis and limit memory CD8+ T-cell survival (Zhang et al., 1998; Kennedy et al., 2000; Nishimura et al., 2000; Fehniger et al., 2001; Yajima et al., 2002; Schluns et al., 2002; Becker et al., 2002). Thus, IL-15 may exert more long-lasting antitumor effects and might be suitable for immunotherapy of non–muscle-invasive bladder cancer.
It is important to understand that tumor cells outside their natural milieu behave quite differently than in their organ of origin (Fidler, 1990; Connor et al., 1993). Based on this understanding, we used an orthotopic bladder tumor model to reproduce the environment that exists in human bladder cancer, which is an appropriate tumor model for evaluating new therapeutic modalities of human bladder cancer. MBT-2 cells, derived from a carcinogen-induced bladder tumor in C3H/He mice, are poorly differentiated transitional cell carcinoma and resemble its human counterpart both grossly and histologically (Horiguchi et al., 2008). IL-2 and IL-12 intravesical gene therapies were previously reported using this novel MBT-2 non–muscle-invasive bladder cancer model (Horiguchi et al., 2000; Horinaga et al., 2005). In the present study, we evaluated the therapeutic effect of in situ IL-15 gene therapy in an orthotopic bladder cancer model and the long-lasting tumor-specific antitumor effect of an in situ IL-15 gene delivery system.
Materials and Methods
Tumor cell line and animals
MBT-2, which was established from N-[4-(5-nitro-2-furyl)-2-thiazolyl]-formamide–induced urothelial carcinoma of the bladder of a female C3H/He mouse, was maintained in Roswell Park Memorial Institute (RPMI)-1640 medium supplemented with 10% heat-inactivated fetal bovine serum, 2 m
Preparation of plasmids and liposome
Eukaryotic expression plasmids encoding murine IL-15 cDNA and beta-galactosidase (β-gal) cDNA were obtained from InvivoGen. β-gal gene plasmid was used as a control and a reporter gene. Liposome GenePORTER reagent (Genlantis) consists of the cationic lipid 1,2-dimyristyloxypropyl-3-dimethyl-hydroxyethyl ammonium bromide (DMRIE) and the neutral lipid dioleoylphosphatidylethanolamine (DOPE) at a 1:1 ratio (both in moles). For our gene therapy protocols, we used a 1:5 ratio of plasmid DNA to GenePORTER reagent, in accordance with a previous report (Horinaga et al., 2005).
In vitro gene transfection
For in vitro transfection, MBT-2 cells were plated at a density of 5×105 cells in 60-mm culture dishes and allowed to incubate for 24 hr at 37°C. DMRIE/DOPE was provided as a dried powder formulation and was resuspended in sterile water to a concentration of 1 mg/ml. For the liposomal transduction, various amounts of the plasmid DNA of IL-15 or β-gal gene (2.5, 5, and 10 μg) were diluted in serum-free medium to a total volume of 500 μl. The corresponding amounts of DMRIE/DOPE (12.5, 25, and 50 μg) were also separately diluted to a total volume of 500 μl and then mixed rapidly together in polystyrene tubes. This 1.0-ml solution was allowed to form plasmid DNA/DMRIE/DOPE lipoplex at room temperature for 30 min, and was then added to the cellular monolayers. After 30 min of incubation at 37°C, the medium containing the lipoplex was aspirated, and the cells were rinsed twice with phosphate-buffered saline (PBS) and then covered with 2.0 ml of normal culture media.
X-gal staining assay and measuring the IL-15 level in vitro
Various amounts of β-gal reporter plasmid (2.5, 5, and 10 μg) were transfected into MBT-2 cells plated at a density of 5×105 cells in 60-mm culture dishes in the same manner as described in vitro gene transfection, and the efficiencies of the gene transfections were determined and calculated in triplicate at 24, 48, and 72 hr after transfection using a X-gal Staining Kit (OZ Biosciences). After fixing the cells with 2% formaldehyde, 5-bromo-4-chloro-3-indolyl-β-
Cytotoxic assay in vitro
To investigate the cytotoxicity of the gene transfections, 24, 48, and 72 hr after in vitro transfection of IL-15 or β-gal gene in 96 wells in a volume of 1×104 cells/100 μl/well culture medium, the number of cells in each well were counted in triplicate using a Premix water-soluble tetrazolium (WST)-1 Cell Proliferation Assay System (Takara Bio). Gene transfection was performed using the same concentration of lipoplex described for in vitro gene transfection. Premix WST-1 (10 μl/well) was added to each well and the plates were incubated for 30 min. Next, the absorbance was measured with a microplate reader at 450 nm with a reference wavelength of 650 nm. Data are expressed as a percentage of survival compared with control.
Intravesical implantation of MBT-2 cells to establish an orthotopic bladder cancer model
All animal experiments were reviewed and approved by our Institutional Animal Care Committees. An orthotopic bladder cancer model was established by intravesical implantation of MBT-2 cells as previously described (Horiguchi et al., 2008). Briefly, the mice were anesthetized with an intraperitoneal injection of 1.5 mg/200 μl pentobarbital sodium. Urine was evacuated from the bladder by mild pressure on the abdomen. A 24-gauge Teflon-coated catheter was introduced into the lumen of the bladder through the urethra. MBT-2 cells, 5×105 in a 50 μl suspension of serum-free RPMI-1640 medium, were then injected into the bladder. The catheter was removed, and to prevent voiding of the MBT-2 cells, the urethra was tied with 4-0 silk thread for 2 hours. Previously our group has evaluated the pathological findings of bladder tumors in mice sacrificed 3 days after the instillation of MBT-2 cell suspension, and found the cells formed tiny superficial tumors, which gradually grew as apparent visible masses (Horiguchi et al., 2008). All the procedures were well tolerated and no mice died postoperatively.
X-gal staining of the whole bladders after in situ gene delivery
At 5 days after the intravesical implantation of the MBT-2 cells, either PBS or lipoplex containing 5 μg of β-gal gene plasmid and 25 μg of GenePORTER was instilled transurethrally into the bladder to a total volume of 50 μl (n=3 in each group). X-gal staining of the whole bladder was performed to confirm the gene delivery in situ. Mice were killed 24 hr after this in situ transfection on day 6. Whole bladders were dissected and harvested into fixative (2% formaldehyde) for 1 hr. The bladders were opened longitudinally and rinsed with PBS, and the entire bladder was then stained for 6 hr in X-gal solution and washed.
Intravesical liposome-mediated IL-15 gene therapy in an orthotopic bladder cancer model
The mice were separated into three groups according to the agent for intravesical liposome-mediated gene therapy (PBS, β-gal, and IL-15; n=10 in each group). For our gene therapy protocols, we used a 1:5 ratio of plasmid DNA (5 μg) to GenePORTER reagent (25 μg) added to serum-free RPMI-1640 medium (50-μl total injection volume per animal), by referring to a previous report that conducted similar experiments (Horinaga et al., 2005). This was close to the maximum dose of plasmid DNA possible due to the bladder capacity limit of the mice. Fifty microliters of PBS was administered as a control. Transurethral injections were performed under anesthesia with an intraperitoneal injection of pentobarbital sodium. Urinary IL-15 levels in each group were monitored with an ELISA kit. We also checked the serum concentrations of IL-15 on day 12 and day 21 in another set of mice (n=3 in each group). Additionally, to examine the potential toxicity of intravesical IL-15 lipofection, we checked the serum concentrations of albumin, total bilirubin, alanine aminotransferase, and creatinine. Each treatment was repeated 5 times at 2-day intervals starting from day 5 after tumor implantation (on day 5, 8, 11, 14, 17, and 20). On day 23, all mice were sacrificed and underwent necropsy to measure bladder wet weight and a histological examination.
Another set of 40 animals were prepared, consisting of 10 mice treated with PBS, 10 mice that underwent β-gal transfection, and 20 mice that underwent IL-15 transfection in situ for the evaluation of survival. Intravesical instillation of PBS or lipoplex containing 5 μg of β-gal or IL-15 gene plasmid was repeated five times at 2-day intervals starting from day 5 after tumor implantation according to the same protocol outlined for gene therapy. Mice were checked daily and the objective findings (i.e., general conditions, palpable masses, and gross hematuria) were recorded. Mice with severe cachexia were euthanized so as to prevent them from suffering, and they were counted as having died with bladder cancer. The mice that died before the end-point were necropsied immediately, and the cause of the death was determined. We confirmed that all mice died due to bladder cancer.
Histological examination and immunohistochemical staining
The dissected bladders were fixed in 10% neutral-buffered formalin and embedded in paraffin. Sections (4-μm) were then stained with hematoxylin and eosin. For CD8 immunohistochemical staining, we used a Biotin-free Catalysed Signal Amplification staining system (K1497) from Dako according to the manufacturer's protocol. Formalin-fixed and paraffin-embedded tissues were deparaffinized three times; in xylene for 1 min, for 1 min each in serial dilutions of ethanol, followed by three changes of water. For antigen retrieval, the sections were heated at 120°C for 10 min in an autoclave. Endogenous peroxidase activity was blocked with 3% hydrogen peroxide for 5 min and nonspecific binding was blocked with skim milk. The sections were then incubated overnight at 4°C with anti-mouse CD8 antibody (ab52854, Abcam) at a 1:50 dilution. Polyclonal Goat Anti-Rabbit Immunoglobulin/HRP (K1497, Dako) was applied to the slides at a 1:100 dilution for 15 min at room temperature. This was followed by incubation with fluorescyl-conjugated phenol (FITC-tyramide, Dako) for 15 min at room temperature and then incubation with anti-FITC antibody conjugated to HRP (Dako) for 15 min at room temperature. Finally, the reaction products were visualized in 20 mg/dl 3,3′-diaminobenzidine tetrahydrochloride (Dako) solution containing 0.006% hydrogen peroxide. Negative control studies were conducted without applying the primary antibody, and mouse spleen sections known to be stained with CD8 antibody were included with each run as a positive control.
Rechallenge with new subcutaneous MBT-2 cells in surviving mice after IL-15 gene therapy
Eight mice in the group treated with in situ IL-15 gene delivery on day 50 survived after the experiment of survival analysis of the gene therapy. These animals were rechallenged with 5×105 MBT-2 cells/200 μl in their right lower abdominal quadrant, and 1×106 FM3A cells into the opposite side at the same time on day 50 to evaluate the tumor-specific immunologic memory against MBT-2 cells. Eight new age-matched control mice were also implanted with the same cells and the same dose. The rechallenged mice and the new control mice received no further treatment. Tumor volumes were measured in cubic millimeters with a vernier caliper and determined according to the following formula: a×b 2/2, where a is the larger and b the smaller of the two dimensions, and continued until day 66 after the initial MBT-2 cell implantation, or day 16 after rechallenge with new MBT-2 cells.
Cytotoxic T-lymphocyte assay
The Cyto Tox 96 nonradioactive cytotoxicity assay (Promega) was performed to determine the cytotoxic activity of CTL in mice treated with IL-15 gene therapy. We used another set of mice treated with in situ IL-15 gene therapy that survived until day 50 and new age-matched control mice. Splenocytes from these mice were harvested and depleted of red blood cells with ammonium chloride Tris buffer. The cells were restimulated with MBT-2 cells treated with mitomycin C at a ratio of 20:1 for 3 days in the presence of human IL-2 (10 U/ml). CD8+ T cells were used as effector cells, which were isolated from the splenocytes by negative selection using CD8+ T-cell enrichment columns (Cedarlane) according to the manufacturer's protocol. MBT-2 cells inactivated with mitomycin C were used as target cells. The effector cells and the target cells were incubated for 8 hr in a total volume of 100 μl in 96-well round-bottom micro-titer plates at various cell densities in order to achieve effector-to-target ratios of 30:1, 10:1, 3:1, and 1:1. FM3A cells inactivated with mitomycin C were used as control target cells. Target cell lysis was calculated as (OD490 nm of sample – OD490 nm with spontaneous release of lactate dehydrogenase (LDH) from target cells – OD490 nm with spontaneous release of LDH from effector cells)×100/(OD 490 nm with maximum release of LDH from target cells –; OD490 nm with spontaneous release of LDH from target cells).
Statistical analysis
Values are presented as the mean±standard error, and analysis of variance for unpaired observations was used. Statistical significance was determined at p<0.05. Survival curves were generated using the Kaplan-Meier method and compared using the log-rank test.
Results
In vitro efficiency of β-gal gene transfection and the secretion of IL-15
The efficiency of gene transfection of each amount of β-gal reporter plasmid is shown in Fig. 1A. The efficiency values were calculated to be 31.1±3.6%, 14.9±1.6%, and 2.1±0.6% with 10μg, 5μg, and 2.5μg of β-gal reporter plasmid at 24 hr, respectively.
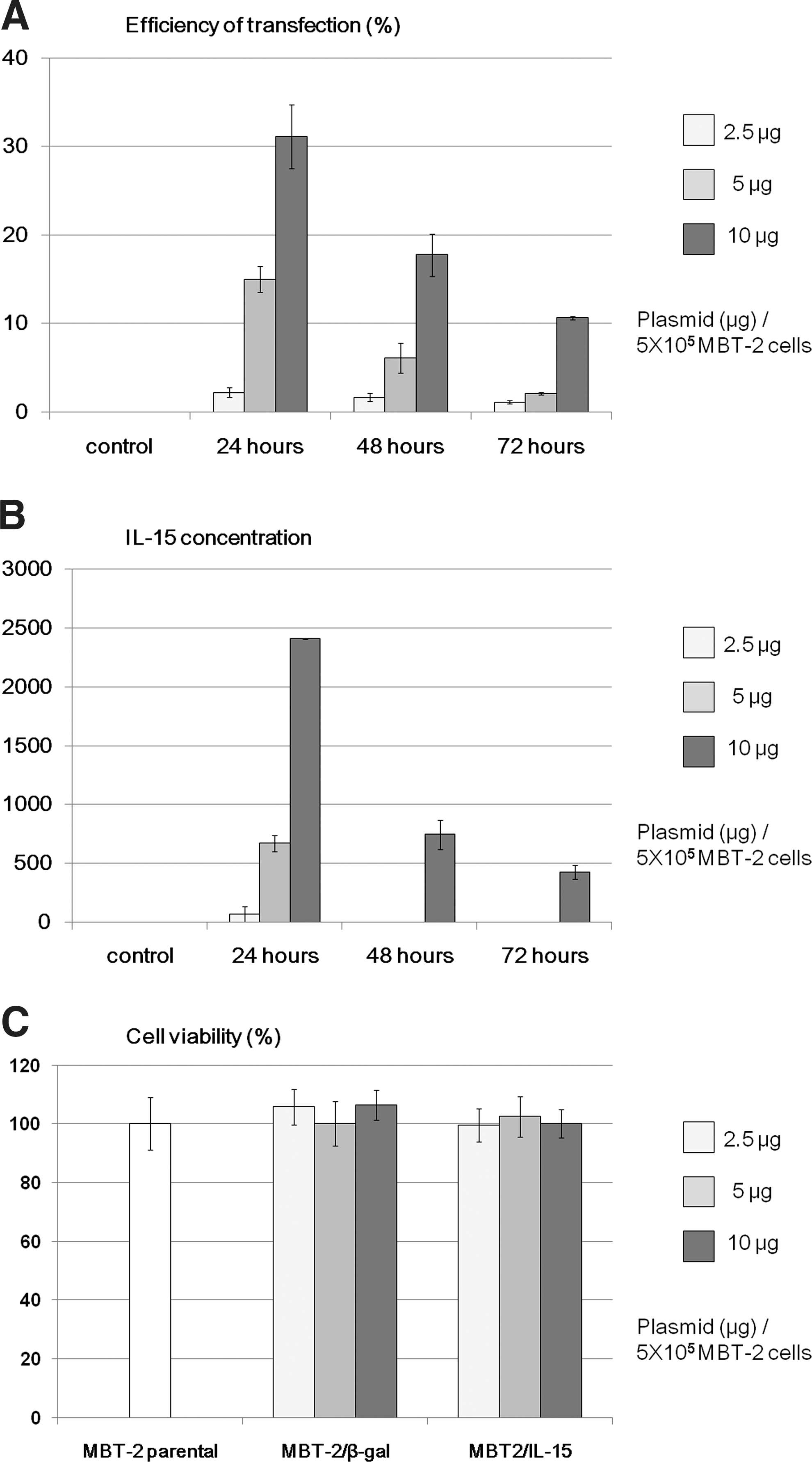
In proportion to the transfection efficiency, the IL-15 concentrations in the culture supernatant were dose dependent after the IL-15 gene plasmid transfection (Fig. 1B). The peak IL-15 level of 2410±5 pg/ml was observed 24 hr after the transfection of 10 μg of IL-15 plasmids. These results showed that the lipoplex composed of 10 μg plasmids had the most efficient transfection in vitro for 5×105 MBT-2 cells.
Cytotoxicity with IL-15 or β-gal gene transfection in vitro
Cytotoxic assay using Premix WST-1 Cell Proliferation Assay System was performed to investigate the cytotoxicity of gene transfection in vitro. As shown in Fig. 1C, MBT-2/IL-15 cells can proliferate in a similar manner as MBT-2 cells and MBT-2/β-gal 24 hr after gene transfection, and similar results were obtained at 48 and 72 hr (data not shown), indicating that liposomal transduction itself at the dose used in this study did not affect cell viability in vitro.
Efficiency of intravesical gene delivery of reporter plasmid for an orthotopic bladder cancer model (X-gal staining)
As shown in Fig. 2A, the gross appearance revealed that, after intravesical instillation of lipoplex containing β-gal reporter gene on day 5, MBT-2 tumors exclusively expressed β-gal on day 6, while the surrounding normal bladder urothelium was not apparently stained by X-gal. Meanwhile, no X-gal staining was observed in the bladders treated with intravesical instillation of PBS alone.

Therapeutic effect of intravesical IL-15 gene delivery in an orthotopic bladder cancer model
Five days after intravesical implantation of MBT-2 cells, lipoplex containing 5 μg of IL-15 gene plasmid and 25 μg of GenePORTER was instilled transurethrally into mice bladders on days 5, 8, 11, 14, 17, and 20, and their urinary IL-15 concentrations were then measured. As shown in Fig. 2B, the first peak urinary IL-15 level was 2606±298 pg/ml 24 hr after the lipofection, following which it gradually decreased and then increased again after the next instillation. Meanwhile in the controls, urine IL-15 concentrations were below the limit of detection at all times. Therefore, gene delivery at 2-day intervals for a total of six times using intravesical instillation resulted in the successful maintenance of urine IL-15 concentrations. We also checked the serum concentrations of IL-15 on day 12 and day 21 (n=3 in each group). However, these concentrations of IL-15 were all below detectable levels, and we did not observe a significant increase in serum IL-15 levels in the IL-15 lipoplex groups compared to controls. Serum levels of albumin, total bilirubin, alanine aminotransferase, and creatinine on day 12 and day 21 were all in the normal ranges in these groups, indicating that intravesical IL-15 lipofection did not cause any systemic side effect.
All mice were still alive after six instillations of PBS, β-gal lipoplex, or IL-15 lipoplex. The mice were sacrificed on day 23 and the bladder weights were measured to examine the growth of the bladder tumors (n=10 in each group). As shown in Fig. 2C, intravesical IL-15 gene therapy in situ resulted in a significant reduction of bladder weight (96±29 mg) compared with those of the control group (196±36 mg) and β-gal group (201±35 mg; for both p<0.05). However, there was no significant difference in bladder weight between the PBS group and β-gal lipoplex group.
Survival analysis of IL-15 gene delivery in situ
In the second set of experiments, the survival rate on day 40 in the IL-15 treatment group was 45% (9 of 20 mice), while in the groups treated with PBS or β-gal lipoplex all of the mice had died by day 40 (n=10 in each group). A significant survival advantage for the IL-15 gene therapy group was found (p<0.05), but there was no significant difference in survival between the PBS group and β-gal treatment group (Fig. 2D). There was no significant difference in the cause of death among the three groups. At the end of the experiment, the tumor eventually occupied the whole bladder lumen, and most of the mice died due to urinary obstruction caused by locally extended bladder tumors. Metastases to the liver and/or lung were observed in only four mice (2 of 10 mice treated with PBS, 1 of 10 mice treated with β-gal, and 1 of 11 dead mice treated with IL-15).
Histological examination and immunohistochemical staining
Histological evaluation revealed cancer nests of various sizes in all 30 bladders treated intravesically with either PBS, β-gal lipoplex, or IL-15 lipoplex. Muscle invasion of tumors (equal to or greater than pathological stage T2) treated with IL-15 was found in 7 of 10 mice, compared with 9 of 10 mice treated with PBS and 8 of 10 mice treated with β-gal (p=0.26 and p=0.61, respectively). Nuclear grades were equally high in all tumors. Lymphovascular invasion was identified in two mice treated with IL-15, compared with four mice treated with PBS and two mice treated with β-gal (n=10 in each group), and there was no significant difference. Figure 3A–C shows the representation of tumors in bladders in each treatment. Huge tumors in bladders treated with PBS alone (Fig. 3A) or β-gal lipoplex (Fig. 3B) were observed. In contrast, tumors treated with IL-15 lipoplex (Fig. 3C) were smaller, and predominant infiltration of inflammatory cells surrounding the tumor was observed in 4 of 10 bladders in the IL-15 group. These immunological responses were not seen at all in the other treated groups. Immunohistochemistry showed that these inflammatory cells were composed of CD8+ cells (Fig. 3D) and no bladders in the PBS only or β-gal groups were CD8+ (figures not shown).
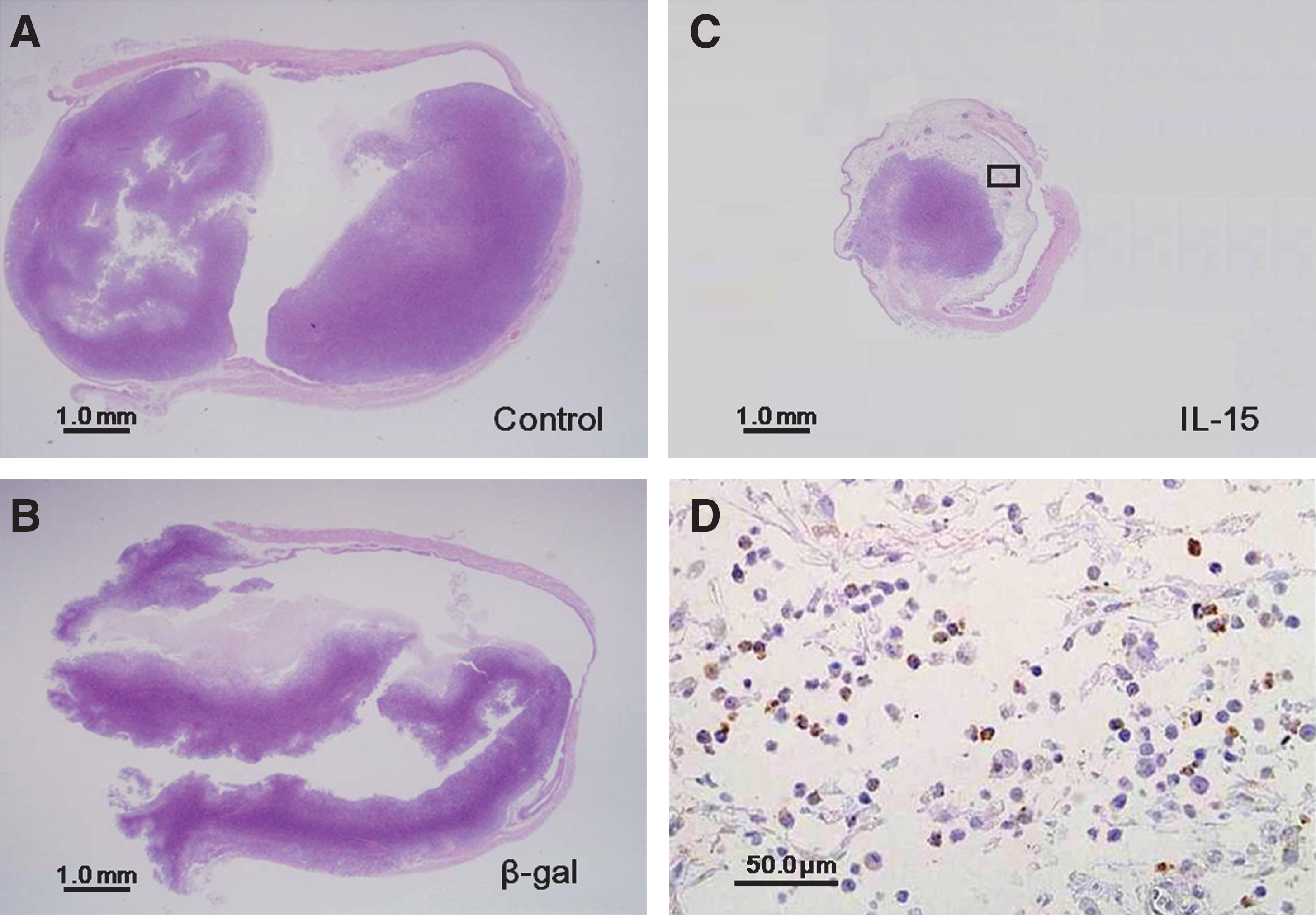
Histological examination of bladder.
Rechallenge experiment
Eight mice in the group treated with in situ IL-15 gene delivery on day 50 survived in the above survival analysis experiment of the gene therapy. These animals received subcutaneous injections of 5×105 MBT-2 cells into the left lower abdominal quadrant, and 1×106 FM3A cells into the opposite side at the same time. The growth of the rechallenged MBT-2 tumors was inhibited again to a significant degree (214±74 mm3) compared with eight new age-matched control mice that underwent implantation with the same cells in the same manner (601±47 mm3; p<0.05) on day 16 after new tumor cell inoculation. However, the growth rate of subcutaneous FM3A tumors (754±153 mm3) was equal to that of control mice (777±204 mm3; p=0.91) as shown in Fig. 4A. This rechallenge experiment was repeated and the result was confirmed.
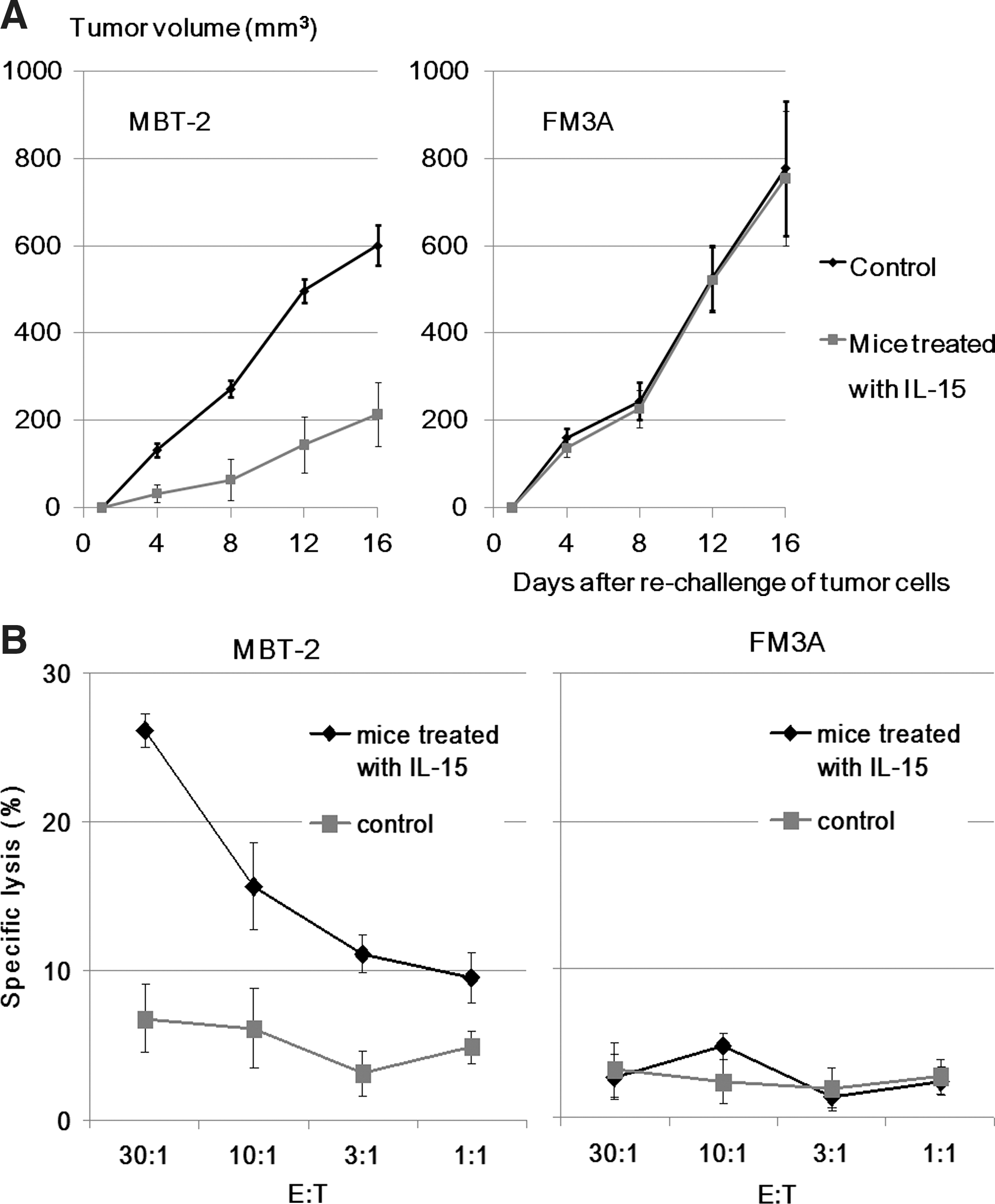
MBT-2 tumor-specific cytotoxic T-lymphocyte responses
We prepared other mice that were treated with in situ IL-15 gene delivery and survived until day 50 and new age-matched mice. CD8+ T cells isolated from the mouse splenocytes were used as effector cells, and MBT-2 cells were used as target cells. We evaluated the CTL response to parental MBT- 2 cells. As shown in Fig. 4B, intravesical IL-15 gene therapy was capable of eliciting a CTL response in a tumor-specific manner, and FM3A used as control target cells showed no specific lysis.
Discussion
This is the first paper to describe the antitumor effect of IL-15 gene transfection in situ in an orthotopic bladder cancer model. In this study involving intravesical administration of murine IL-15 lipoplex, we confirmed IL-15 in mouse urine, which was believed to be secreted from IL-15 gene-transfected tumor cells. Thereafter significant bladder tumor suppression was observed with the infiltration of CD8+ T cells, and about 40% of the mice survived after day 40, while none of the mice treated with PBS or the same dose of β-gal plasmid and DMRIE/DOPE lipoplex survived. These results seem to indicate that our intravesical gene therapy was only effective when it resulted in attracting a host immune response against parental tumor cells by local IL-15 secretion from in situ transfected tumor cells themselves, but not by the effect of inflammatory cytokine production derived by artificial trauma or liposome instillation itself. The inflammation caused by the intravesical IL-15 therapy seemed not to be systemic but rather localized because the urine concentration was maintained throughout the follow-up period, although the serum IL-15 level did not increase and was the same level as in the controls. Furthermore, all mice that initially survived the intravesical IL-15 gene therapy rejected the rechallenged MBT-2 cells on day 50 without any additional treatments, suggesting that intravesical IL-15 gene therapy may be capable of inducing long-lasting tumor-specific immunologic memory. Previously, we evaluated the efficacy of intravesical BCG therapy in the same orthotopic bladder cancer model (Horinaga et al., 2005). A significant survival advantage was observed in mice treated with seven instillations of 400 μg of BCG compared with the vehicle control group. However, all surviving mice that were treated with the BCG therapy died due to a new bladder tumor after being rechallenged with MBT-2 cells on day 60, a finding that was the same as that in new age-matched controls. These results suggest that intravesical BCG instillation could induce an inflammatory reaction in the bladder, although it seems to lack long-lasting and tumor-specific protective immunity, unlike the IL-15 gene therapy described in the present study.
Several ex vivo studies have used IL-15 gene immunotherapy to treat cancer. A previous report demonstrated the suppression of tumorigenesis in human prostate cancer cells (PC-3) transfected with a chimeric IL-15 gene in nude mice. The researchers hypothesized that NK cells mediate the antitumor effect of IL-15 on PC-3 tumor cell growth (Suzuki et al., 2001). However, the exact immunologic aspects induced by the therapy must be evaluated with a syngeneic wild-type mouse cancer model. Nude mouse and SCID mouse models cannot be used to evaluate the immunologic events of antitumor effects because they lack T cells. Our orthotopic bladder cancer model most closely mimics the clinical situation, a prerequisite for drawing meaningful conclusions. Another report showed that genetically modified MethA fibrosarcoma cells producing IL-15 underwent complete rejection, in a response characterized by massive infiltration of CD4+ T cells and neutrophils (Hazama et al., 1999). Similar data that subcutaneous murine colon adenocarcinoma tumor cells transfected with IL-15 regressed spontaneously in contrast with the tumors of wild-type cells were also reported (Tasaki et al., 2000). However, these vaccine strategies were difficult to apply to clinical trials due to their subcutaneous cancer model and to the cumbersome ex vivo gene transfer procedures. Returning tumor cells back into a patient is potentially hazardous, even if ex vivo gene-modified tumor cells are irradiated. The bladder represents an ideal target for gene therapy in situ based on the relatively noninvasive access provided through catheterization, eliminating the need for systemic administration. Intravesical administration of vectors allows for direct local tumor contact, circumventing the difficulties associated with tumor targeting via systemic administration.
Although previously used retroviral vectors have demonstrated highly efficient gene delivery in a bladder cancer model and long-term persistence of transduction in an orthotopic murine bladder cancer model (Siemens et al., 2003; Kikuchi et al. 2007), they require active cell division for efficient gene transfer (Jaffee et al., 1993). In addition, the generation of retroviral vectors is complex and associated with safety issues for human trials that include the production of replication-competent recombinant virus (Miller et al., 1990). Due to these limitations of the retroviral vector system as a transfection vehicle, we chose cationic liposomes for our intravesical gene delivery of IL-15 due to their safety, low cost, and ease of preparation (Anderson, 1998). However, liposome methods have disadvantages, including low gene transfer efficiency and transient gene expression. Therefore, we adopted multiple intravesical protocols (a total of six instillations) to overcome these disadvantages. In addition, immunological gene therapy does not necessarily require whole tumor cells to be transfected because certain amounts of cytokines may derive sufficient host immunity. Furthermore, intravesical instillation of lipoplex was reported to potentially deliver the gene that was specific to bladder tumors (Horiguchi et al., 2000). Similarly, another study found that β-gal expression detected by X-gal staining was limited only to tumor cells in their in vivo model for peritoneal dissemination of ovarian cancer in which they used cationic liposomes for the gene delivery (Lee et al., 2002). The protective glycosaminoglycan (GAG) layer of the normal bladder urothelium may act as a physiological barrier to the lipoplex, while absence of the GAG complex allows bladder tumor transfection to occur. In contrast, the biodistribution of adenoviral vector gene delivery after intravesical instillation exhibited nonspecific effects in the bladder (Wood et al., 1999). Thus, we speculate that the tumor-specific biodistribution of intravesical liposomal gene delivery is another advantage of this technique.
Our results in vivo showed that using liposome-mediated in situ gene transfer via simple intravesical instillation, intravesical IL-15 plasmid transfection in situ exhibits significant tumor suppression in an orthotopic bladder cancer model, and that CD8+ T cells seemed to be associated with its antitumor effect. In mice that survived the intravesical IL-15 gene therapy, rechallenged MBT-2 cells were suppressed and they had the potential to acquire tumor-specific immunologic memory against MBT-2 cells because inoculated FM3A cells were not rejected after the intravesical IL-15 gene therapy. In conclusion, the present data indicate that IL-15 gene therapy may be a promising new adjuvant therapy for non–muscle-invasive bladder cancer and a useful substitute for BCG treatment.
Footnotes
Acknowledgments
This work was supported in part by Grants-in-Aid for Young Scientists (B) from The Ministry of Education, Culture, Sports, Science and Technology of Japan.
Author Disclosure Statement
The authors declare no conflict of interest.