
Abstract
Adenovirus (Ad)-based antiangiogenesis gene therapy is a promising approach for cancer treatment. Downregulation or loss of coxsackievirus and adenovirus receptor (CAR) is often detected in various human cancers, which hampers adenoviral gene therapy approaches. Cationic liposome-complexed adenoviral vectors have been proven useful in CAR-deficient cells to enhance therapeutic gene transfer in vivo. Here, we investigated the antitumor effects of recombinant adenovirus encoding endostatin (Ad-hE) encapsulated in cationic liposome (Ad-hE/Lipo) on CAR-deficient CT26 colon carcinoma murine models. In vitro, Ad-hE/Lipo enhanced adenovirus transfection in CAR-deficient cells (CT26), and endostatin gene expression was measured by both qualitative and quantitative detection. In addition, an antibody neutralizing assay indicated that neutralizing serum inhibited naked adenovirus 5 (Ad5) at rather higher dilution than the complexes of Ad5 and cationic liposomes (Ad5-CL), which demonstrated that Ad5-CL was more capable of protecting Ad5 from neutralization. In vivo, Ad-hE/Lipo treatment in the murine CT26 tumor model by intratumoral injection resulted in marked suppression of tumor growth and prolonged survival time, which was associated with a decreased number of microvessels and increased apoptosis of tumor cells. In conclusion, recombinant endostatin adenovirus encapsulated with cationic liposome effectively inhibited CAR-deficient tumor growth through an antiangiogenic mechanism in murine models without marked toxicity, thus showing a feasible strategy for clinical applications.
Introduction
Among the endogenous proteins with selective effects on vascular endothelial cells, endostatin, a 20-kDa COOH-terminal proteolytic fragment of the basement membrane component collagen XVIII, was originally discovered in the supernatant of a murine hemangioendothelioma cell line as an inhibitor of proliferation of endothelial cells in vitro and a strong inhibitor of angiogenesis in vivo (O'Reilly et al., 1997). Although endostatin is one of the first antiangiogenesis compounds to be evaluated for the treatment of patients with colorectal cancer, there are still some obstacles in clinical applications with endostatin owing to the relatively short half-life and in vivo instability, with daily serum levels only briefly reaching that which is associated with antiangiogenic activity in vitro (Mancuso and Sternberg, 2005). In this regard, gene therapy by which endostatin is expressed in vivo has the potential to produce the therapeutic agent at high concentration in a local area for a sustained period. It has been reported that high circulating levels of biologically active endostatin could be achieved through adenovirus-mediated gene transfer (Feldman et al., 2000; Sauter et al., 2000).
Adenoviral (Ad) vectors are efficient gene delivery systems and are one of the obvious choices for cancer gene therapy (Sharma et al., 2009). However, their uptake by cells in many cases requires the coxsackievirus and adenovirus receptor (CAR), a transmembrane protein initially described as the high-affinity attachment receptor for the group C adenovirus fiber protein (Bergelson et al., 1997). Thus, lack of CAR expression, which is the situation in CT26 mouse colon cancer cells (Lee et al., 2004), is a potential cause of intrinsic resistance of tumor cells to adenovirus-based therapies. Unfortunately, CAR expression is frequently altered in gastrointestinal malignancy, as well as under hypoxic conditions most frequently demonstrating loss or reduction of CAR expression (Korn et al., 2006; Kuster et al., 2010). Furthermore, loss of CAR expression may be associated with malignant progression and increased tumor aggressiveness (Matsumoto et al., 2005; Anders et al., 2009). This may account for some of the limited success in past clinical trials using adenoviral vectors (Relph et al., 2005). More recent studies suggest that limitations of gene therapy approaches involving the use of adenoviral vectors, due to the CAR deficiency of target cells, could be overcome by complexing the adenovirus with cationic liposomes (Okegawa et al., 2001; Han et al., 2008). By facilitating adenovirus binding to the cell surface via an electrostatic interaction, conjugation of adenovirus with cationic liposome has been shown to be effective in increasing adenoviral transgene expression on CAR-deficient cells (Kerr, 2003).
Cationic liposomes based on dioleyloxypropyl trimethylammonium chloride are potent nanocarriers to deliver various drug molecules to tumors and have demonstrated success both in in vitro and in vivo gene delivery (Korn et al., 2006). In a previous study, we prepared a cationic liposome as a nanocarrier to encapsulate recombinant adenovirus encoding human endostatin (Ad-hE/Lipo), which showed good cell viability and high transfection efficiency in CAR-deficient SKOV-3 cells. Furthermore, systemic administration of Ad-hE/Lipo was well tolerated and resulted in marked suppression of tumor growth and reduced immunogenicity of adenovirus in an established ovarian cancer model (Yang et al., 2010). This liposome system is efficient and safer for gene delivery, and may promote the potential of recombinant adenovirus encoding endostatin in the treatment of CAR-deficient tumors. In the current research, we evaluated the efficacy of the antitumor activities of intratumorally administered cationic liposome-complexed recombinant human endostatin adenovirus in CAR-deficient CT26 colon carcinoma murine models. Our study indicated that the complex can arrest tumor growth through the inhibition of angiogenesis in vivo, and is well tolerated by recipient mice. The current study helps to improve the efficiency and safety of adenoviral gene therapy for future clinical application.
Materials and Methods
Cell lines and animals
Mouse colorectal carcinoma cell line CT26 and human embryonic kidney (HEK293) cells were obtained from the American Type Culture Collection (ATCC, Manassas, VA). They were maintained in monolayer cultures in RPMI 1640 or Dulbecco's modified Eagle's medium (DMEM; Gibco/Invitrogen, Carlsbad, CA), supplemented with 10% heat-inactivated fetal bovine serum (FBS), at 37°C in a humidified atmosphere containing 5% CO2. Human umbilical vein endothelial cells (HUVECs) were collected and cultured as described (Lasic, 1998; Yao et al., 2005). Female BALB/c mice (6–8 weeks old) were purchased from the Laboratory Animal Center of Sichuan University (Chengdu, Sichuan, China).
Recombinant adenoviral vector construction
Recombinant human endostatin adenovirus was constructed in our laboratory as described previously (Wu et al., 2005). Briefly, total RNA was extracted from human liver tissue (TRIzol; Invitrogen, Carlsbad, CA). The full-length human endostatin cDNA (about 570 bp) was amplified by RT-PCR, using primers 5′-CGGGATCCACAGCCACCGCGACTTCCAGCC-3′ and 5′-GCGGATCCTACTTGGAGGCAGTCATGAAGC-3′, with a BamHI restriction site in both primers (Titan one tube RT-PCR kit; Roche, Mannheim, Germany). After sequence confirmation, the cDNA was cloned into a shuttle vector for the rescue of the recombinant E1/E3-deleted adenovirus (AdEasy system) as described earlier (He et al., 1998). The viral particles were amplified in 293 cells, purified by two-step cesium chloride gradient ultracentrifugation, and measured by absorption (A 260). The viral titer was quantified in a standard TCID50 assay. The recombinant green fluorescent protein adenoviral vector (Ad-GFP; replication-defective adenovirus type 5) was obtained.
Preparation of adenovirus vector complexed with cationic liposome
Cationic liposomes were prepared in our laboratory and are described briefly as follows: mixtures of DOPE/DOTMA/CHOL at various weight ratios were dissolved in chloroform–methanol (3:1, v/v) and evaporated to dryness under reduced pressure in a rotary evaporator. The resulting lipid film was hydrated in 5% dextrose (2 mg/ml) and sonicated until solubilized completely in a sonicator. The liposomes were stored at 4°C until use. Particle size was measured by dynamic laser scattering, using a Nano ZS particle sizer (Malvern Instruments, Malvern, UK). The ζ potential of the cationic liposomes was measured with a Zetasizer Nano ZS90 (Malvern Instruments). Before the measurements, the samples were diluted 1:5 in distilled water. Each sample was analyzed in triplicate, and the ζ potential was derived from mobility of particles in electric field by applying the Smoluchowski relationship.
To prepare complexes of Ad-GFP or Ad-hE and cationic liposomes (Ad-GTP/Lipo or Ad-hE/Lipo), blank cationic liposomes with various lipid concentrations according to the needs of the following gene transfer studies were added to an equal volume of Ad-GFP or Ad-hE and incubated at room temperature for 15 to 30 min before use.
Adenovirus infection
CT26 cells were transfected with complexes representing various ratios of cationic liposome to Ad-GFP. In brief, cells were seeded into 6-well plates at a density of 2×105 cells per well and used at 70–80% confluence. Various amounts of liposome (1, 5, 10, 15, 20, 50, 100, 500, and 1000 μg, respectively) were incubated with a fixed amount of Ad-GFP for 30 min at room temperature, and then added to CT26 cells. Forty-eight hours after infection, the percentage of GFP expression was determined by fluorescence-activated cell-sorting (FACS) analysis.
Effect of neutralizing antibodies in vitro for CT26 cells
Rabbits were immunized with 109 plaque-forming units (PFU) of Ad-null by subcutaneous injection once per week for 4 weeks. Eight days after the fourth adenovirus injection the rabbits were killed, and the neutralizing activity in blood from rabbits treated with adenovirus was assayed with CT26 cells. CT26 cells (2×105) were seeded in 6-well plates. The next day, serum was diluted 10- to 50-fold with RPMI 1640. According to the optimized ratio, complexes of Ad-GFP and cationic liposome were used to infect CT26 cells with or without serum containing a neutralizing antibody at 37°C for 30 min. In brief, cells were washed with phosphate-buffered saline (PBS), and the mixture was added to CT26 cells. After a 4-hr incubation in a 5% CO2 incubator at 37°C, cells were washed with PBS to remove the complexes and 2 ml of fresh medium with 10% FBS was added. Cells were then incubated for an additional 48 hr before assessing GFP expression. Expression of GFP in infected cell samples was visualized with an Olympus microscope. Furthermore, we used Ad-hE/Lipo that optimized ratio of 2.50×109 viral particles [VP]/μg, with Ad-hE as the control group, to infect CT26 cells at a multiplicity of infection (M.O.I.) of 50 in the presence of various concentrations of Ad-neutralizing antibody. The level of endostatin production in the supernatants was determined by ELISA.
Western blot analysis
CT26 cells were transfected with the complexes Ad-hE/Lipo, Ad-hE alone, and control adenovirus (Ad-null), respectively. Cells were conditioned at 37°C for 48 hr, and the supernatant was harvested and concentrated by ultrafiltration (Centricon YM-3; Millipore, Bedford, MA) and analyzed by Western blot. After loading equal volumes (10 μl) per lane, the supernatants were separated on a 12% sodium dodecyl sulfate (SDS)–polyacrylamide gel and transblotted onto a polyvinylidene difluoride (PVDF) membrane. The membrane was blocked with TTBS (Tris-buffered saline including 0.1% Tween 20) with 5% nonfat milk, and later it was probed at 4°C overnight with rabbit polyclonal anti-endostatin antibody (Santa Cruz Biotechnology, Santa Cruz, CA) diluted 1:500. Blots were then incubated with horseradish peroxidase-conjugated anti-rabbit immunoglobulin (diluted 1:5000; Sigma-Aldrich, St. Louis, MO). Protein bands were detected with an enhanced chemiluminescence (ECL) detection kit (GE Healthcare Life Sciences, Piscataway, NJ).
ELISA determination of recombinant endostatin
Using a protocol similar to one described previously (Yao et al., 2005), levels of endostatin from the cloned cells were measured in triplicate, using standard ELISA kits 48 hr after infection according to the manufacturer's instructions (Chemicon/Millipore, Temecula, CA).
In vitro bioactivity assay of endostatin
A proliferation inhibition assay using different fold dilution series of conditioned supernatant (Ad-hE/Lipo) on HUVECs was performed as previously described (Yao et al., 2005). Briefly, 2×105 CT26 cells, at 70–80% confluence in 35-mm dishes, were infected with Ad-Null or Ad-hE/Lipo at an M.O.I. of 50 or left untreated (NS). After 48 hr, serum-free supernatant was harvested and sequentially concentrated over Centriplus spin columns (Millipore). HUVECs were seeded in 96-well plates (2×104/ml) and treated with a 20-μl volume of various fold dilutions of conditioned supernatant. Survival of HUVECs after treatment was quantified by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) colorimetric assay (EZ4U kit; Biomedica, Vienna, Austria). Each well was supplemented with 50 μl of a 2-mg/ml solution of MTT in complete medium and incubated at 37°C for 4 hr. The medium and MTT solution were then removed, and 150 μl of dimethyl sulfoxide was added to each well. Absorbance was read at 540 nm with a microplate reader. Data represent the average of six wells, and the experiment was repeated three times.
Animal models and in vivo effects of Ad-hE/Lipo
All studies involving mice were approved by the institute's animal care and use committee. CT26 cells (1×106) were injected into the flanks of BALB/c mice, 6–8 weeks of age. To explore the therapeutic efficacy of Ad-hE/Lipo in a subcutaneous tumor model, we treated the mice on day 7 after implantation of tumor cells, when the tumors were 3–5 mm in diameter. They were randomly assigned to 5 groups (10 mice per group). Tumors were injected with 1×109 PFU of Ad-hE, Ad-hE/Lipo, Ad-N (Ad-null), or Ad-N/Lipo (Ad-null/Liposome) suspended in 100 μl of carrier (PBS), or left untreated (NS). Treatments were given once every 3 days, for a total of four times. Injections were distributed equally into each of four tumor quadrants. Tumors were measured with calipers in two dimensions and the tumor volume was determined by the following formula: tumor volume (mm3)=π/6×length (mm)×width (mm)×width (mm). Gross measures such as weight loss, ruffling of fur, behavior, and feeding were investigated To observe survival time, mice were inoculated and treated in the same way as previously (n=10 per group); residual mice in all groups were killed 7 weeks after tumor establishment.
Immunohistochemical analysis and TUNEL assay
Paraffin-embedded tissue sections from treated mice were processed and stained with an antibody reactive to CD31 according to standard immunohistochemical techniques. Sections were then stained with labeled streptavidin–biotin reagents (LSAB kit, peroxidase; Dako, Carpinteria, CA). Vessel density was determined by counting the number of microvessels per high-power field in the sections. The fluorescence in situ terminal deoxynucleotidyltransferase-mediated nick end labeling (TUNEL) assay was performed with an in situ apoptotic cell detection kit (Boehringer Mannheim, Indianapolis, IN) in accordance with the manufacturer's protocol. It is based on the enzymatic addition of digoxigenin nucleotide to the nicked DNA by terminal deoxynucleotidyltransferase. For the observations of potential side effects in the treated mice, the tissues of heart, liver, spleen, lung, kidney, and so on, were also fixed in 10% neutral buffered formalin solution and embedded in paraffin. Sections of 4–5 μm were stained with hematoxylin and eosin (H&E). Sections subjected to H&E, immunohistochemical staining, and TUNEL assay were observed by two pathologists in a blinded manner.
Data analysis and statistics
The results of statistical analyses are presented as means±SD. For comparison of individual time points, analysis of variance (one-way analysis of variance) and an unpaired Student t test were used. Survival was analyzed by the Kaplan–Meier method and compared by log-rank test. p<0.05 was considered statistically significant.
Results
Optimization of cationic liposome and adenoviral vector ratio
Varying the DOPE/DOTMA/CHOL ratio resulted in complexes of different sizes and different net surface charge. The particle size and ζ potential distribution of the cationic liposomes are summarized in Table 1. The mixtures of DOPE/DOTMA/CHOL in 1:1:0.1 weight ratio were ratios at which the complexes formed small particles and there was a net cationic charge. On the basis of the preceding data, we chose cationic liposomes with a DOPE/DOTMA/CHOL weight ratio of 1:1:0.1, characterized as optimal in terms of both size distribution and ζ potential, for further assessment of the effect of adenovirus on liposome-complexed adenoviral gene transfer in CT26 cells. Fixed amounts (5×1010 VP/μl) of Ad-GFP were incubated with various concentrations of cationic liposome (1, 5, 10, 15, 20, 50, 100, 500, and 1000 μg) as used for CT26 cell infections and adenoviral GFP expression was measured by FACS. We found that adenoviral GFP expression in CT26 cells increased (67.79±7.7%) with the concentration at 2.50×109 VP/μg cationic liposome (Fig. 1). Next, we used cationic liposome-encapsulated Ad-GFP at a ratio of 2.50×109 VP/μg to infect CT26 cells in the presence of Ad-neutralizing antibody. However, the preexisting neutralizing antibody failed to influence the susceptibility of CT26 cells to infection by the cationic liposome-encapsulated Ad-GFP (Fig. 1). This indicated that cationic liposome-encapsulated Ad-GFP can infect CAR-negative cells in a CAR-independent way.
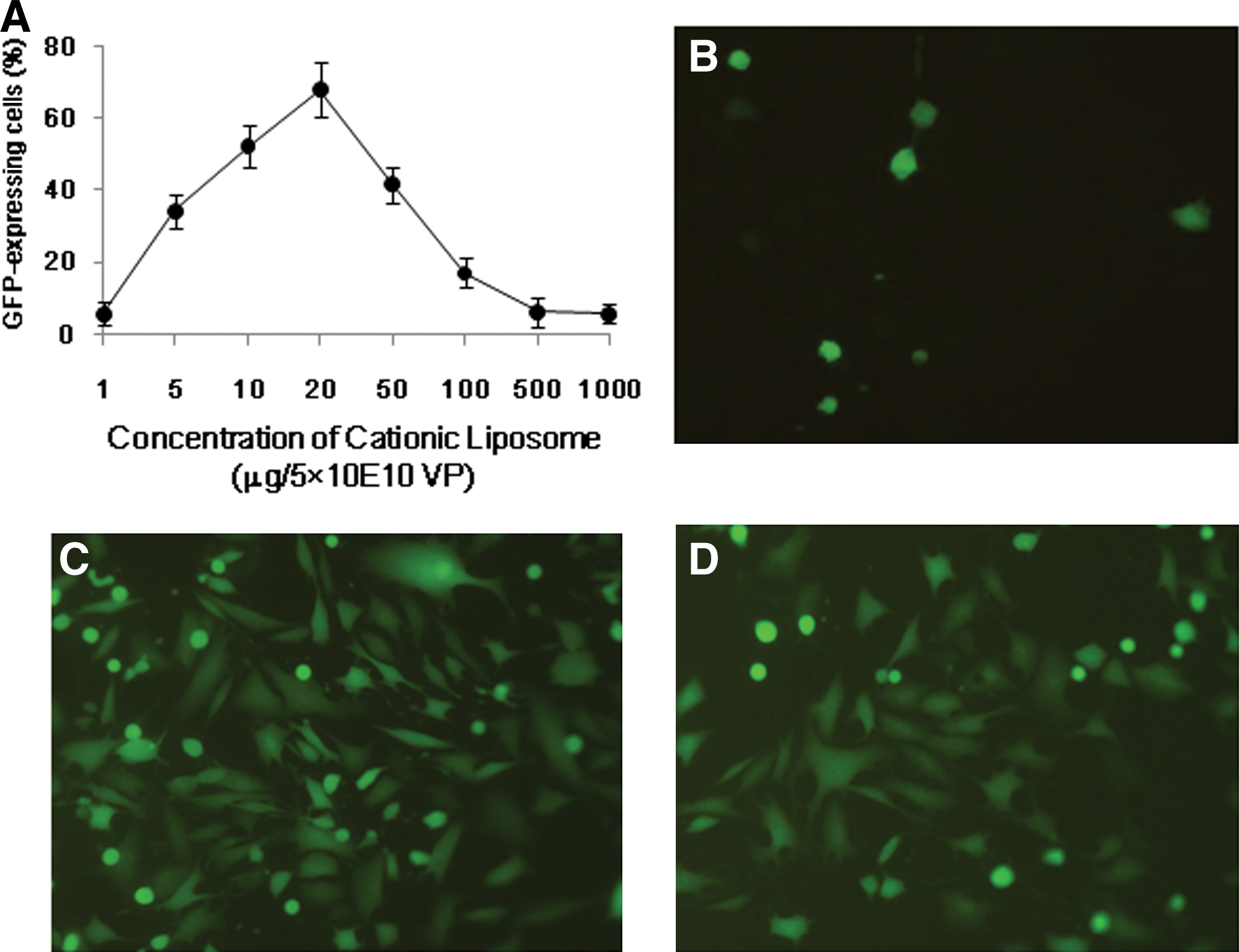
The Ad–cationic liposome complex shows enhanced transfection capacity in CT26 cells. CT26 cells were transfected at various cationic liposome (CL) and Ad-GFP ratios. Forty-eight hours after infection, the percentage of GFP expression was determined by FACS analysis
Results are expressed as means±SD of at least three measurements.
CL, cationic liposomes.
Transfection efficiency of Ad-hE/Lipo and expression of biologically active endostatin in vitro
We then investigated whether this system could offer more efficient in vitro gene transfer to CT26 cells than Ad-hE alone. We also used Ad-hE and Ad-hE/Lipo to transfect CT26 cells. As shown in Fig. 2A, adenoviral endostatin expression significantly increased when adenovirus was complexed with cationic liposome (468.87±46.7 at 50 M.O.I., and 964.59±65.8 at 500 M.O.I.). Thus, it is clear that complexation with cationic liposome leads to adenoviral gene expression in CAR-deficient cancer cell lines when the M.O.I. is 50–500. Moreover, preexisting neutralizing antibody failed to block the Ad-hE/Lipo infection of CT26 cells or to decrease the level of endostatin production (Fig. 2B). The biological activity of secreted endostatin was tested against primary HUVECs. Conditioned medium from Ad-hE/Lipo-transfected CT26 cells apparently inhibited HUVEC growth, compared with conditioned medium from CT26 cells transfected with Ad-hE or nontransfected cells (Fig. 2C). Secreted endostatin protein was also identified by Western blot (Fig. 2D).
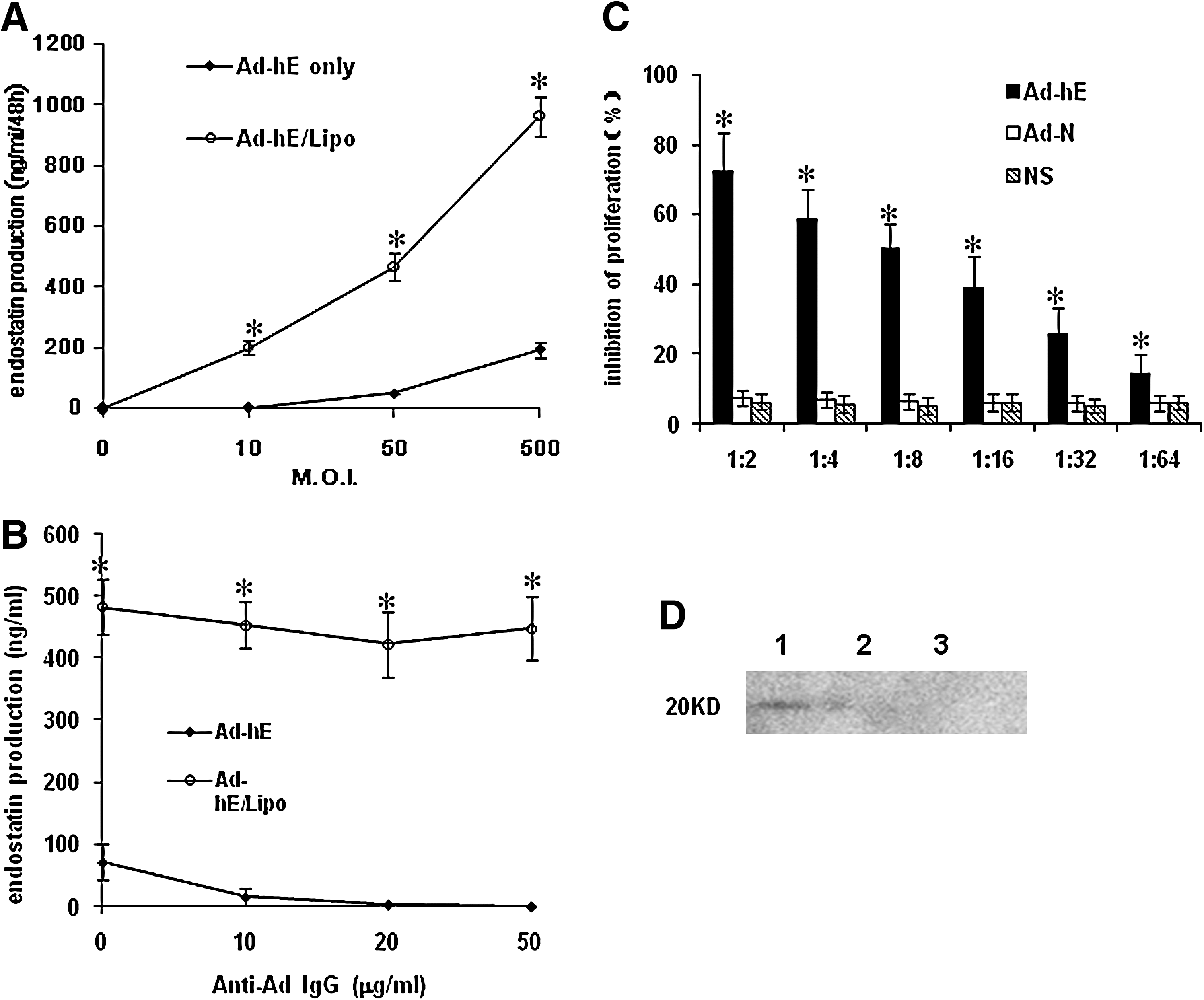
Enhancement of endostatin production after Ad-hE infection in combination with cationic liposomes (CL) in vitro. CT26 cells were infected with or without CL (10 ml) at various viral titers (0–500 M.O.I.)
Encapsulation of cationic liposome enhances antitumor effects of Ad-hE in vivo
In this study, CT26 cells and a subcutaneous BALB/c mouse tumor model were used to analyze the therapeutic potential of Ad-hE/Lipo. We first investigated whether cationic liposome can synergize with endostatin to improve the antitumor effect by intratumoral injection. To explore the feasibility of the approach, when small palpable tumor nodules could be easily seen under the skin, the first injection of Ad-hE/Lipo complexes (1×109 PFU of Ad-hE plus 100 μg of liposome) was administered intratumorally on day 7 after tumor implantation. Treatments were given once every 3 days for a total of four times. The mice had to be killed, and tumor samples taken, in week 7 after tumor cell implantation because of excessive tumor volumes. Results showed that treatment with Ad-hE/Lipo resulted in a significant arrest of tumor growth compared with nontreatment (NS) or treatment with Ad-N, Ad-N/Lipo, or Ad-hE (p<0.05; Fig. 3A).
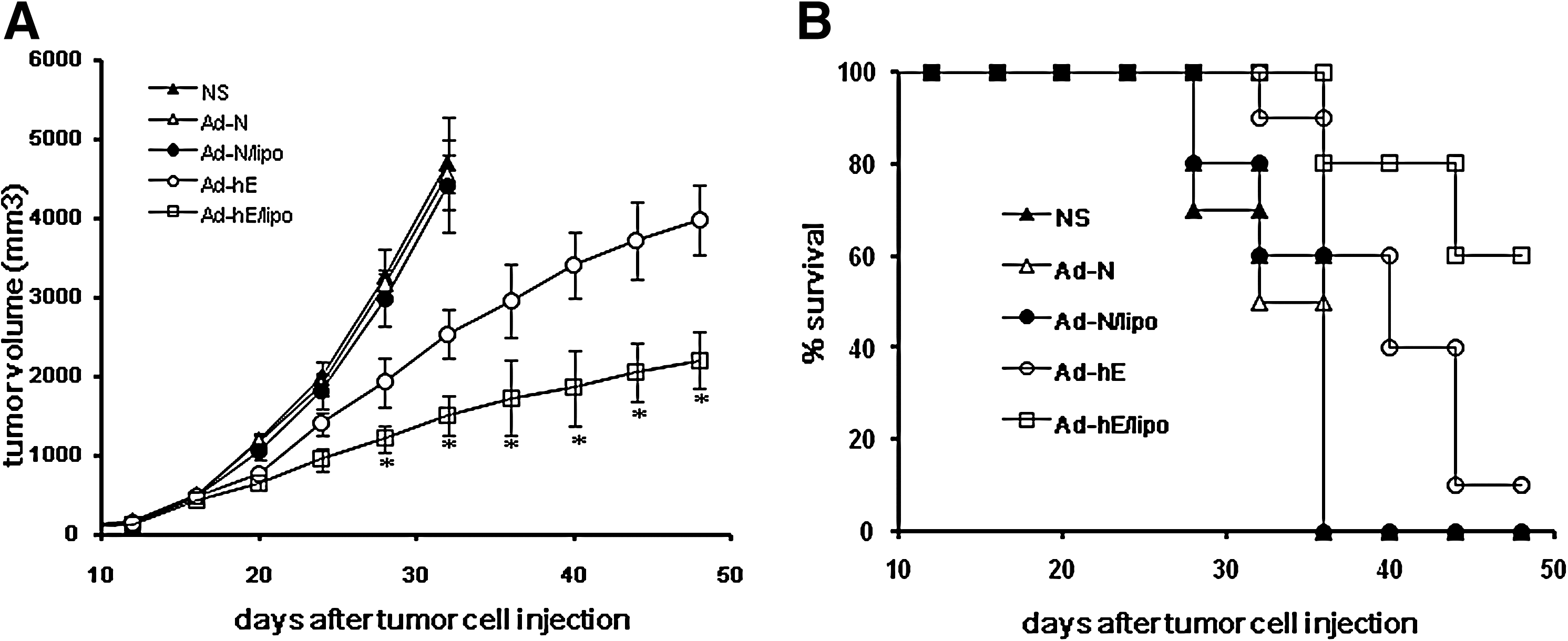
Cationic liposome-encapsulated Ad-hE showed enhanced antitumor effects through intratumoral injection. Ad-hE/Lipo therapy significantly decreased tumor volume in mice
In all cases, the cause of death was tumor progression or cachexia because of the tumor burden. The survival time of the Ad-hE/Lipo treatment group was significantly longer than that of the other groups (i.e., the Ad-N, Ad-N/Lipo, NS, and Ad-hE groups) (p<0.05).
Effective angiogenesis inhibition and increased apoptosis through Ad-hE/Lipo
To evaluate the consequence of antiangiogenesis therapy, paraffin-embedded tumor tissue sections were stained with an endothelial-specific antibody against CD31. Microvessel density was then counted. Decreased microvessel densities were found in Ad-hE- and Ad-hE/Lipo-treated tumors, and it was lower in the Ad-hE/Lipo-treated group than in the Ad-hE-treated group (p<0.05; Fig. 4).
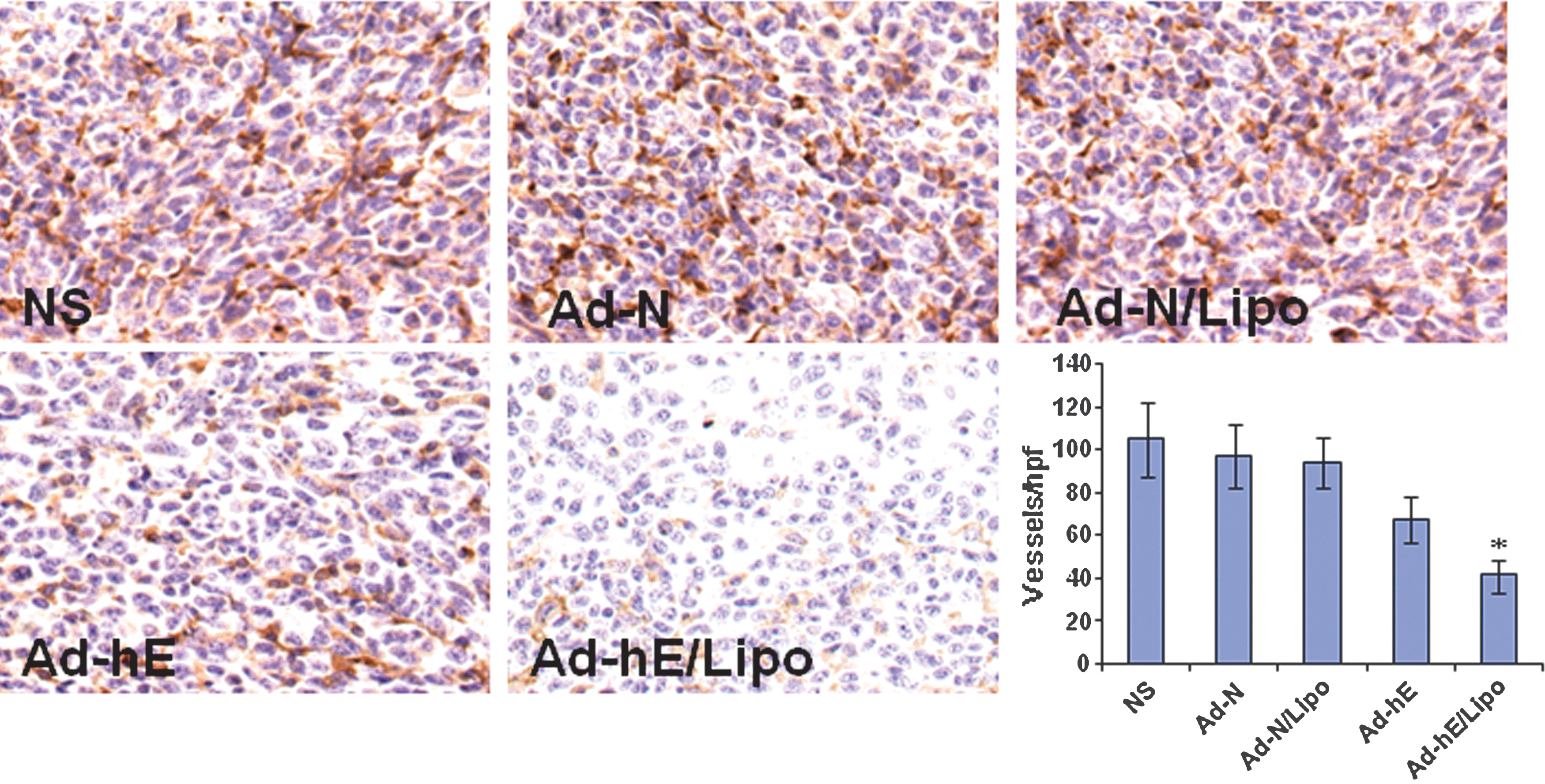
Inhibition of tumor angiogenesis by Ad-hE/Lipo intratumoral injection. Tumors were harvested from mice that were not treated (NS) or treated with Ad-N, Ad-N/Lipo, Ad-hE, or Ad-hE/Lipo. Paraffin-embedded tissue sections were fixed and treated with anti-CD31 antibody. The stained sections were visualized. Significantly decreased microvessels were observed in the Ad-hE/Lipo-treated group compared with the control groups, including Ad-hE (*p<0.05). Original magnification: ×400. hpf, high-power field.
Apoptotic cells in tumors were determined by TUNEL assay. The liposome control group was found to induce low levels of apoptotic cells in the tumor region, whereas the number of apoptotic cells in the Ad-hE/Lipo-treated group was significantly higher than in the other groups, including Ad-hE alone (p<0.05; Fig. 5).
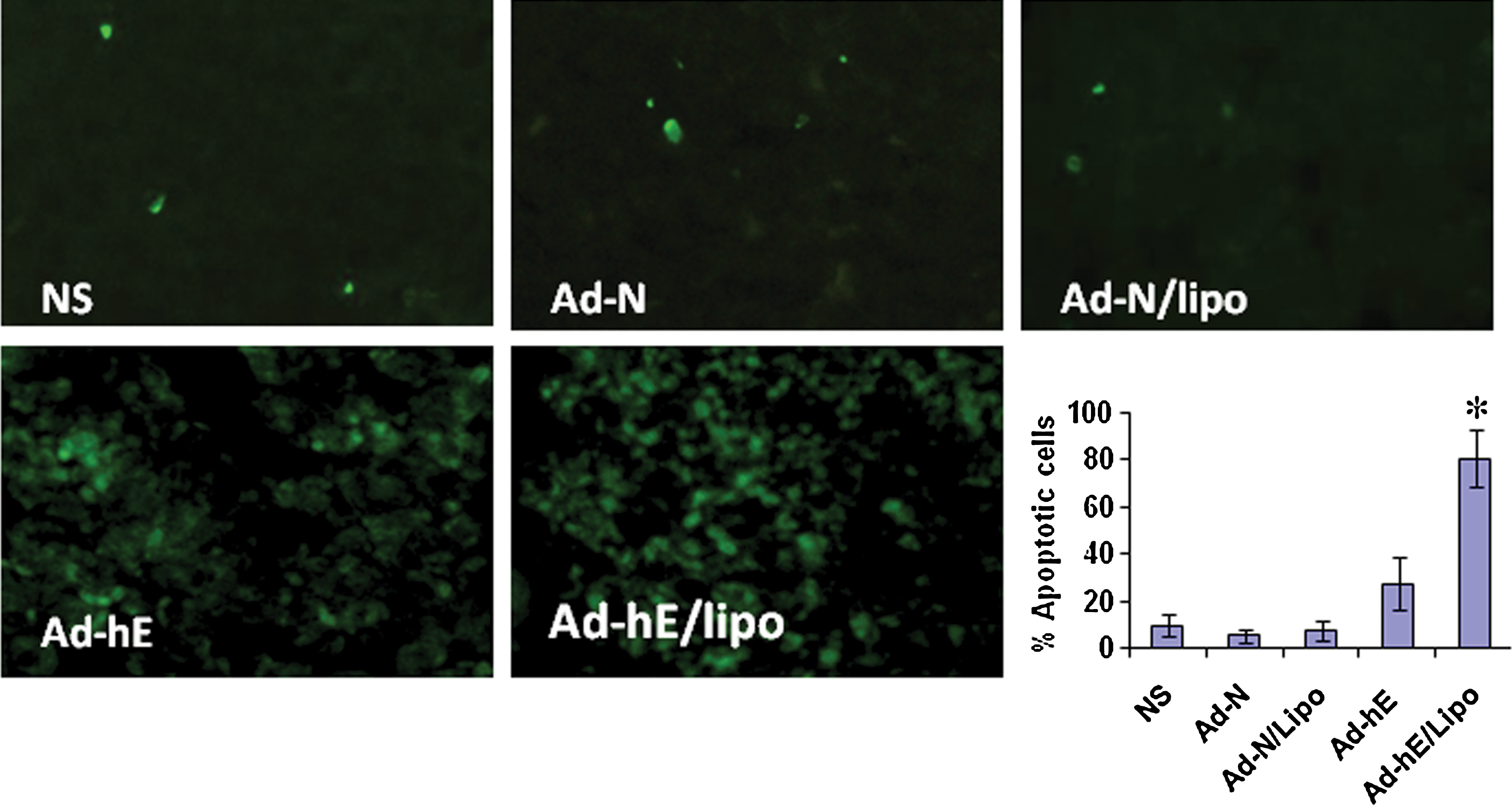
Ad-hE/Lipo treatment resulted in increased apoptosis in CAR-deficient CT26 colon carcinoma murine models. Frozen histological sections of tumors were stained with a TUNEL kit and examined by immunofluorescence microscopy. Compared with the NS, Ad-N, Ad-N/Lipo, and Ad-hE groups, locally increased apoptosis was found in tumors from the Ad-hE/Lipo-treated group (p<0.05). *p<0.05, means±SD. Original magnification: ×200. Color images available online at
Toxicity observation
To evaluate the health status of mice treated by Ad-hE/Lipo injection, the weight of mice was monitored once every 3 days throughout the whole experiment and considered a parameter for evaluation of systemic well-being, anorexia, or cachexia. No significant differences in weights were found among any of the groups. No adverse effects in other gross measures, such as skin ulcerations or death, were observed in the Ad-hE/Lipo group. Furthermore, toxic pathological changes in the liver, lungs, kidneys, spleen, brain, and heart were not detected by microscopic examination.
Discussion
Gene therapy is a promising approach for therapeutic intervention and has great therapeutic potential for patients with advanced colorectal cancer, incurable by conventional treatments. Several features of the natural history of CRC render it a good candidate for gene therapy such as gene replacement, virus-directed enzyme–prodrug therapy, immune manipulation, and virotherapy, all of which have entered clinical trials but have not yet demonstrated significant therapeutic benefit (Kerr, 2003). Many of these gene transfer vectors are based on adenovirus; to refine the approach, continuing efforts should be made to improve the antitumor potency, efficiency of gene delivery, and accuracy of gene targeting. Angiogenesis and its fine balance are essential for tumor growth and invasion in colorectal cancer. Antiangiogenesis gene therapy strategies leading to increased production of endogenous angiogenesis inhibitors would seem perfectly suited to support the approach by tipping the balance toward a more antiangiogenic state (Tandle et al., 2004). Moreover, that CRC is often confined to specific organs or compartments offers an advantage through regional delivery of a genetic vector to maximize tumor exposure and minimize systemic toxicity (Kerr, 2003).
Several observations have been made in the present study concerning gene therapy treatment based on recombinant human endostatin-expressing adenoviruses, encapsulated with and without cationic liposome in vivo and used in CAR-deficient CT26 murine models. The intratumoral injection of Ad-hE/Lipo showed significant antitumor efficiency in the inhibition of CAR-deficient tumor angiogenesis, increasing apoptotic cells, decreasing tumor volume, and extending survival.
In the present study, we constructed cationic liposome-complexed recombinant human endostatin-expressing adenovirus, which expressed biologically active recombinant endostatin as shown in vitro. Expression of recombinant endostatin was confirmed in the supernatant from transfected cells by Western blotting analysis and ELISA. Treatment of endothelial cells with conditioned medium obtained from transfected CT26 cells resulted in apparent inhibition of endothelial cell proliferation. In addition, complexes of Ad5 and cationic liposomes were more capable of protecting Ad5 from neutralization. The transfection efficiency of liposome-delivered genes was highly dependent on the lipid composition, lipid/DNA ratio, particle size of the liposome–DNA complex, and cell lines used (Cohen et al., 2001). We prepared cationic liposome and complexed it with adenovirus, optimizing the lipid composition and charge ratio: at a DOPE:DOTMA:CHOL ratio of 1:1:0.1 the diameter was 73±15 nm, and the ζ potential was 37.5 mV. Our study showed that the expression of endostatin was fairly high in CAR-deficient CT26 cells when 2.5×109 VP of adenovirus per microgram of cationic liposome was administered. Defects in the capillary endothelium of tumor vasculature are typically in the size range of 400 to 600 nm; therefore, liposomes having diameters smaller than 400 nm can efficiently extravasate and accumulate within the tumor interstitial space (Zou et al., 2000). In addition, tumors have impaired lymphatic drainage, which provides extravasated liposomal drug with a long retention time in tumor tissues (Yuan et al., 1995).
In vivo gene transfer by cationic liposomes depends on the manner of administration. On intratumoral injection, there is higher intratumoral retention of liposome complex accompanied by improved intratumoral diffusion and predominant transgene expression in the first tissue the liposomes encounter (Yuan et al., 1995; Bao et al., 2006). Liposomes accumulate at tumor sites because of their ability to extravasate through “pores” or “defects” in the capillary endothelium. These pores seem to be a consequence of the rapid angiogenesis occurring in tumors and are generally not present in normal tissues or organs (Brandwijk et al., 2007). Direct intratumoral delivery methods have significant potential advantages and the feasibility of the approach has been greatly enhanced (Bao et al., 2006). Remarkably, in our study, immunohistochemical analysis showed that in intratumorally injected mice treated with Ad-hE/Lipo angiogenesis was apparently inhibited in tumor tissue. Moreover, blocked angiogenesis accompanied by high proliferation balanced by apoptosis in tumor cells from the mice treated with Ad-hE/Lipo, and the promotion of apoptosis was probably responsible for the antitumor activity. The mechanism by which endostatin mediates tumor cell apoptosis may be due, in part, to the loss of adequate vasculature, which would deprive the tumor of oxygen and nutrients. Endostatin appears to be active in the initial phase of neovascularization in the primary immature vascular network (Kuo et al., 2001; Indraccolo et al., 2002). Endostatin activity may differ between tumor models or depend on timing or the vector used for endostatin gene transfer (Dutour et al., 2005). Controversially, other investigators have shown that endostatin is less effective at interfering with angiogenesis in established tumors (Benjamin et al., 1998). Thus, this may explain the delayed CT26 tumor progression that occurred instead of the expected tumor regression. Our study suggests that recombinant adenovirus encoding endostatin and encapsulated with liposome could work better than adenovirus without liposome.
The binding of adenovirus to its primary receptor may be an important rate-limiting step for gene transfer, and, therefore, various strategies have been evaluated to modify adenovirus tropism in order to circumvent CAR deficiency (Kanerva et al., 2003). Both viral and nonviral vectors possess positive and negative characteristics; investigators have started to combine aspects of each vector type into hybrid vectors that capitalize on the positive characteristics (Wagner, 2008). By combining adenovirus and liposomes, the positive characteristics of both viral and nonviral delivery systems can be exploited (Zhong et al., 2010). For example, cationic liposomes redirect the tissue tropism of adenovirus and overcome the immune barrier (Ma et al., 2002; Han et al., 2008), but meet significant barriers on endosomal escape and nuclear uptake. Adenovirus, however, possesses an inherent capacity to destabilize endosomes and enter nuclei (Seth, 1994). Our current approach took advantage of the conjugation of adenoviral vector with cationic liposomes, which overcame the limitation due to the CAR deficiency of target cells, brought great potential for efficient in vivo gene delivery, and enhanced the antitumor effect. Although these results showed that antiangiogenesis gene therapy resulted in reduced tumor angiogenesis, leading to apoptotic death of CT26 cancer cells and effectively delaying the growth of CAR-deficient colon cancer in murine models, it is also important to note that the complex was unable to inhibit tumor growth fully and further challenges remain. Further studies are necessary to determine the optimal therapeutic schema to obtain better therapeutic efficacy.
Taken together, our data in the present study indicate that cancer gene therapy by intratumoral injection of recombinant adenovirus encoding endostatin encapsulated with cationic liposome, was effective in the inhibition of angiogenesis and tumor growth in CAR-deficient colon cancer murine models. Our findings may help to improve adenovirus-mediated antiangiogenesis gene therapy.
Footnotes
Acknowledgments
The study was supported by the Program for New Century Excellent Talents in University (NCET) (NCET-09-0575), the National 863 Project of China (2007AA021202), the National Nature Science Foundation of China (no. 30470623), and the National Basic Research Program of China (973 Program), no. 2011CB711000.
Author Disclosure Statement
No competing financial interests exist.