
Abstract
Heart disease is the leading cause of morbidity and mortality, and cardiac gene transfer has potential as a novel therapeutic approach. We previously demonstrated safe and efficient gene transfer to the canine heart using a percutaneous transendocardial injection procedure to deliver self-complementary (sc) adeno-associated virus 6 (AAV6) vector. In the present study, we proceed with our vertical translation study to evaluate cardiac gene transfer in nonhuman primates (NHPs). We screened approximately 30 adult male rhesus macaques for the presence of neutralizing antibodies against AAV6, AAV8, and AAV9, and then selected seven monkeys whose antibody titers against these three serotypes were lower than 1/5. The animals were then randomized to receive either scAAV6 (n=3), scAAV8 (n=1), or scAAV9 (n=3) vector expressing the enhanced green fluorescent protein (EGFP) reporter gene at a dose of 5.4×1012 genome copies/kg, which was administered according to a modified version of our previously developed transendocardial injection procedure. One animal treated with scAAV6 died secondary to esophageal intubation. The remaining animals were euthanized 7 days after gene transfer, at which time tissue was collected for analysis of EGFP expression, histopathology, and biodistribution of the vector genome. We found that (i) transendocardial delivery of AAV is safe in the NHP, (ii) AAV6 and AAV8 provide efficient cardiac gene transfer at similar levels and are superior to AAV9, and (iii) AAV6 is more cardiac-specific than AAV8 and AAV9. The results of this NHP study may help guide the development AAV vectors for the treatment of cardiovascular disease in humans.
Introduction
Although much work has recently been performed evaluating cardiac gene transfer in large animals such as sheep, pigs, and canines as described above, very little investigation has been undertaken in the nonhuman primate (NHP) model. Both lentivirus (Tarantal et al., 2005) and AAV (Tarantal and Lee, 2010) have been evaluated following fetal delivery of reporter constructs, and it was found that AAV induced higher levels of transgene expression than lentivirus and that AAV9 and AAV10 were superior to AAV5. In addition, a separate group demonstrated approximately fourfold greater transgene activity following injection with AAV9 compared with AAV1 in neonatal NHPs (Pacak et al., 2006). However, evaluation of cardiac gene transfer in adult NHPs has not been described in the literature.
We recently developed an efficient method for cardiac gene transfer in the canine using a percutaneously inserted injection catheter (Bish et al., 2008, 2011), and our goal in this study was to use a modified version of this approach to evaluate the cardiac gene transfer efficiency of self-complementary (sc) AAV6, AAV8, and AAV9 in the adult rhesus macaque. We report that (i) transendocardial delivery of AAV is safe in the NHP, (ii) AAV6 and AAV8 provide efficient cardiac gene transfer at similar levels and are superior to AAV9, and (iii) AAV6 is more cardiac-specific than AAV8 and AAV9.
Materials and Methods
Vector design and production
Each vector was designed to express the enhanced green fluorescent protein (EGFP) reporter gene under control of the constitutive chicken β-actin promoter with cytomegalovirus (CMV) enhancer (CB promoter). Vectors were produced according to the previously described pseudotyping protocol by the Vector Core of the Children's Hospital of Philadelphia (Herzog et al., 1999). In brief, recombinant AAV genomes containing AAV2 inverted terminal repeats (ITRs) were packaged by triple transfection of 293 cells with a cis-plasmid containing the EGFP transgene, an adenovirus helper plasmid, and a chimeric trans-plasmid containing the AAV2 rep gene fused to the capsid gene of the AAV serotype of interest. Self-complementary vectors contained a mutation in the termination sequence of the 5′ ITR to allow synthesis and encapsidation of a dimeric inverted repeat of the transgene cassette (McCarty et al., 2001).
Animal use and vector delivery protocol
All animal experiments were performed at the Chengdu National Center for Safety Evaluation of Drugs in Chengdu, China. All animals were handled in compliance with the Association for the Assessment and Accreditation of Laboratory Animal Care International and institutional guidelines that were approved by the Institutional Animal Care and Use Committee of the Chengdu National Center for Safety Evaluation of Drugs. Thirty adult male rhesus macaques were screened for the presence of neutralizing antibodies (NAbs) against AAV6, AAV8, and AAV9. Titer was determined by incubating Huh7 cells with serial dilutions of canine serum and AAV-CMV-EGFP of the serotype in question and observing the dilution at which the number of green fluorescent protein (GFP)-positive cells was reduced by 50% compared with control wells.
Seven macaques (6–10 kg) whose titer against all three serotypes was <1:5, which is the detection limit of the assay, were selected for this study. The animals were randomized to receive AAV6 (n=3), AAV8 (n=1), or AAV9 (n=3). The vector delivery procedure was performed under general anesthesia, and macaques were placed in left lateral recumbency. Heart rate, respiratory rate, systolic blood pressure, electrocardiogram, and oxygen saturation were monitored throughout the anesthetic period. A right carotid arterotomy was performed, and a 7 French introducer was placed in the vessel, followed by insertion of the injection catheter. This catheter was a steerable injection catheter with an adjustable length core needle (MyoCath, Bioheart Inc., Sunrise, FL), which was previously used to deliver AAV vector to canines in our laboratory (Bish et al., 2008, 2011). The catheter was flushed with heparinized blood prior to vector infusion to prevent inactivation of the virus (Marshall et al., 2000). Next, under fluoroscopic guidance, the catheter was advanced into the left ventricular cavity, and by steering the needle tip and adjusting the needle length, approximately 60 transendocardial injections of 250 μl each were performed to target the left ventricular free wall and interventricular septum from base to apex and from endocardium to epicardium with AAV vector. Contrast medium was added to the vector solution so that injection sites could be visualized. This allowed us to differentiate between injected and uninjected regions of the heart and helped to ensure that the vector solution was distributed globally throughout the myocardium.
For each procedure, vector was mixed with 2 ml of sterile contrast solution (Omnipaque) and diluted with sterile saline to produce 15 ml for injection. Lidocaine (2 mg/kg as a bolus followed by a constant rate infusion at 50 μg/kg/min) was initiated if ventricular tachycardia developed during the procedure. After recovery, animals were treated with carprofen for 2 days and amoxicillin-clauvolonic acid for 5 days.
Histologic analysis of transgene expression and pathology
Macaques were euthanized 7–8 days following treatment, and tissues were harvested for determination of reporter transgene expression and vector biodistribution. For analysis of GFP fluorescence, the heart was divided into four sections along the short axis from the apex to base and fixed in 4% paraformaldehyde overnight at 4°C. The tissue was then washed 3×10 min in PBS and dehydrated in 20% sucrose overnight at 4°C. Next, each section was frozen in OCT embedding compound (Tissue Tek, Torrance, CA), and 10-μm cryosections were prepared. Slides were mounted with Vectashield DAPI media (Vector Laboratories, Burlingame, CA) and examined for GFP fluorescence using a Leitz DMRBE fluorescent microscope (Leica, Bannockburn, IL) equipped with a Micro Max digital camera (Princeton Instruments, Trenton, NJ) interfaced with Image Pro Plus software (Media Cybernetics, Bethesda, MD). For each animal, four representative photographs were recorded under constant exposure conditions using the 10× objective from the interventricular septum and left ventricular free wall from the endocardium to the epicardium in each of the four heart sections. A liver sample was also processed as above to monitor hepatic GFP expression. For pathology, slides were stained independently with hematoxylin and eosin (H&E) and trichrome (Sigma, St. Louis, MO) to identify mononuclear infiltrate and fibrosis, respectively.
Biodistribution analysis
For biodistribution analysis, samples were snap-frozen in liquid nitrogen. Following DNA extraction, genome copy titers were quantified by TaqMan PCR (Applied Biosystems, Foster City, CA) using primers and probes designed against the EGFP transgene. One hundred nanograms of template DNA was used per reaction for triplicate reactions, and several controls were performed to confirm the specificity and accuracy of the PCR. A spike control was performed in which the test sample was spiked with exogenous test assay target to rule out PCR inhibition. The assay was also performed on samples from a negative control, saline-injected animal to determine background signal of the assay, which was negligible [<1 genome copy (gc) per 5,000 cells, or >1 log lower than the minimum genome copies detected in experimental samples]. Finally, a control endogenous gene [glyceraldehyde 3-phosphate dehydrogenase (GAPDH)] was run as a loading control and displayed minimal sample-to-sample variation (<0.25 cycle) with 100 ng of template DNA.
Results
Study design
In this NHP study, we compared the cardiac gene transfer efficiency of three AAV vectors 1 week following transendocardial injection via an injection catheter inserted through a peripheral artery. Each AAV vector was designed to express the EGFP reporter gene under control of the chicken β-actin promoter with CMV enhancer (AAV-CB-EGFP). Rhesus macaques were randomized to receive 5.4×1012 gc/kg of either AAV6 (n=3), AAV8 (n=1), or AAV9 (n=3). These seven animals were selected from a pool of approximately 30 macaques after their NAb titer against AAV6, AAV8, and AAV9 was determined to be <1:5, the lower limit of detection of the assay used.
Cardiac GFP expression
GFP expression was evaluated in the NHPs 7 days after delivery of AAV6, AAV8, or AAV9. Expression of GFP was minimal following injection with AAV9 (Fig. 1D). In contrast, high levels of GFP expression were observed throughout the heart following injection with AAV6 (Fig. 1B) and AAV8 (Fig. 1C). It should be noted that the AAV8 expression is based on n=1 and should be considered preliminary.
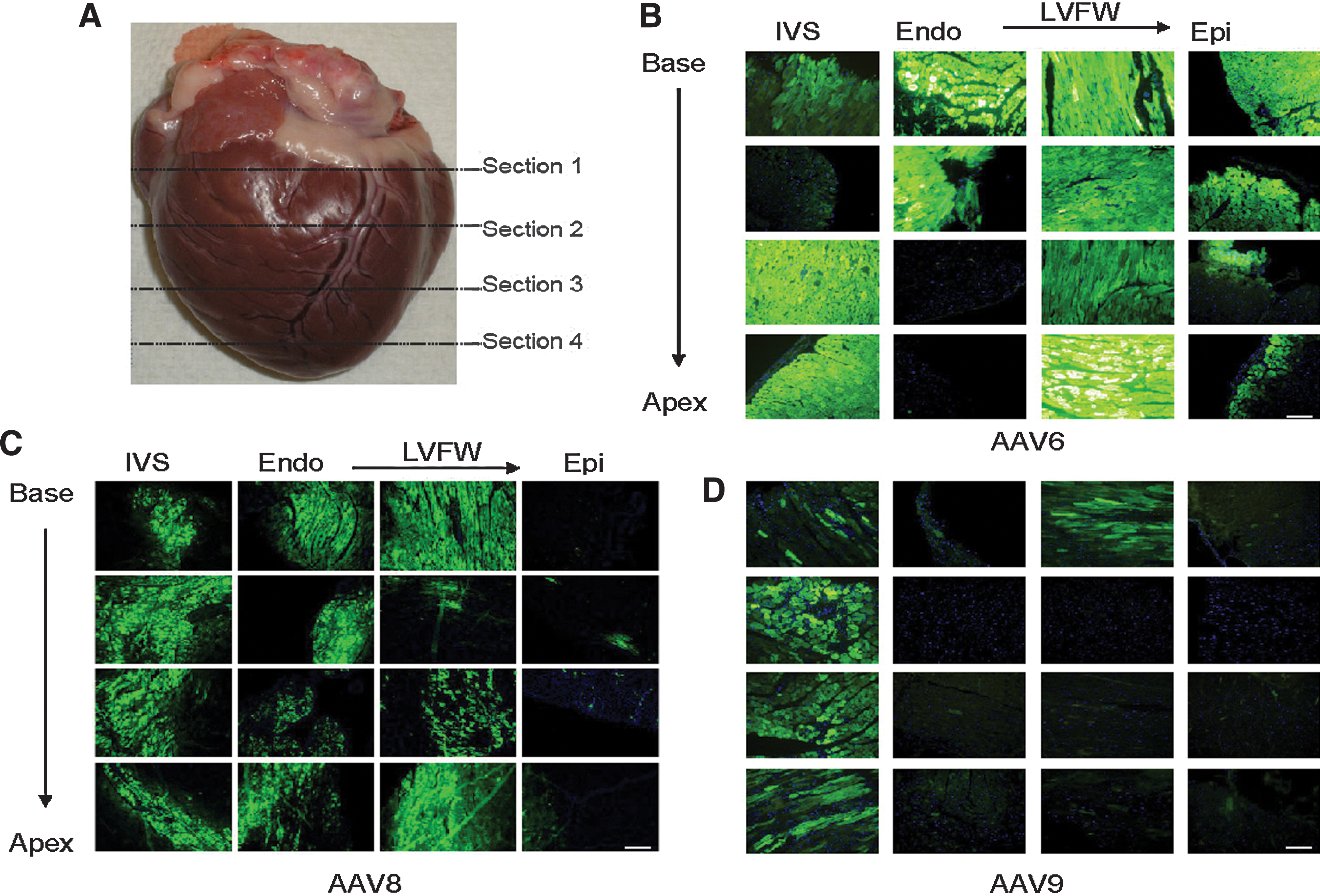
Representative images (10× objective) of GFP expression in the NHP heart.
Biodistribution studies
AAV vector genomes were detected in all tissues examined (Fig. 2A); however, the ratio of genomes detected in the heart to genomes detected in other tissues varied significantly from vector to vector (Fig. 2B). Of the three vectors examined, AAV6 had the highest cardiac specificity with heart to tissue ratios ranging from 0.5 in the liver to approximately 25 in the spleen and up to 1,000 in other tissues. In contrast, the heart to organ ratios for the other vectors evaluated were approximately 0.1 or less for the liver and 0.2 or less in the spleen (Fig. 2B). GFP fluorescence was also minimal in liver sections from animals treated with AAV6 compared with AAV8 (Fig. 2C).
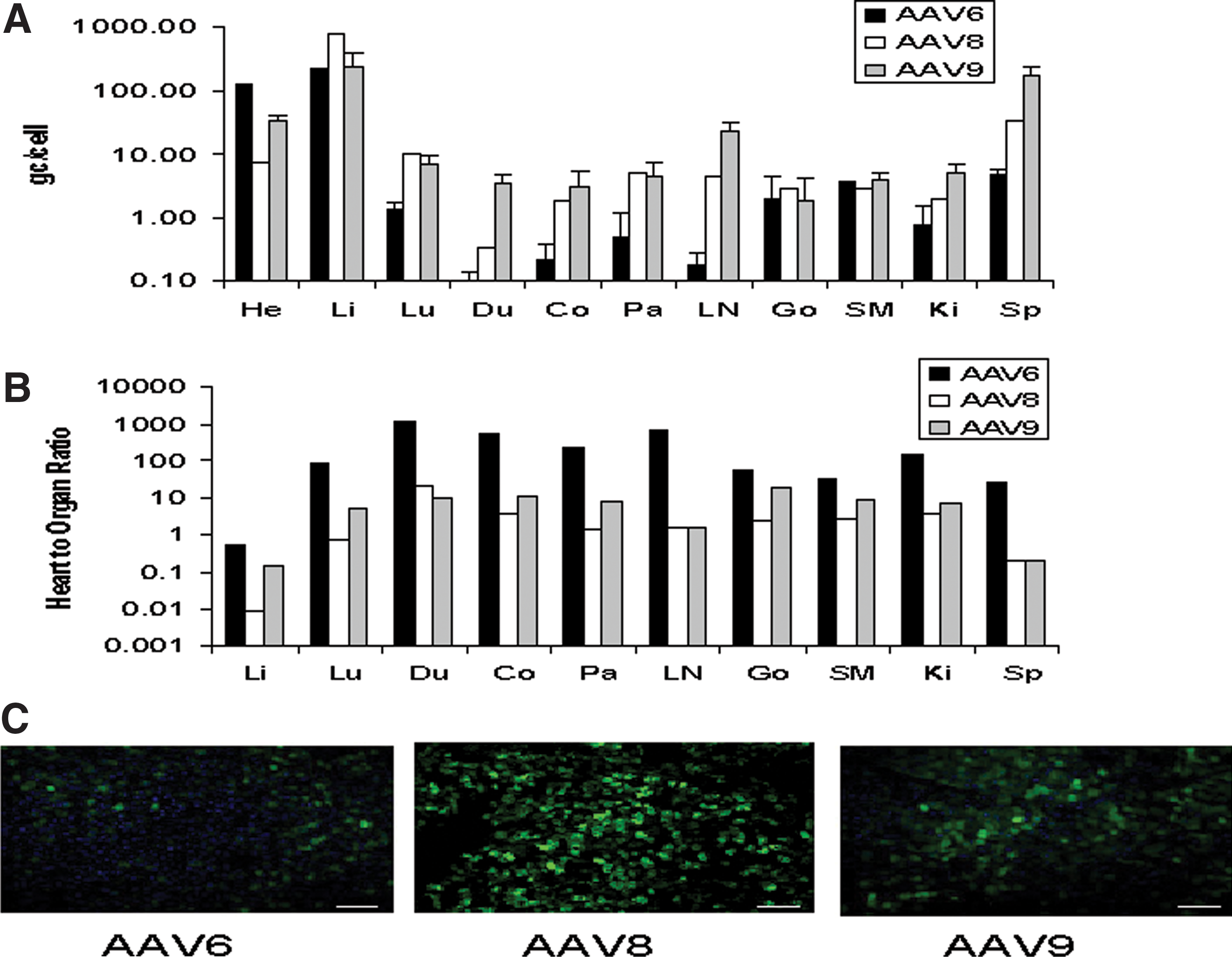
Biodistribution of AAV vectors 7 days following delivery to the NHP heart.
Safety of transendocardial delivery of AAV vectors
One animal treated with AAV6 died in postprocedure recovery, and it was determined that death occurred secondary to esophageal intubation. No mortality was associated with the injection procedure itself. Ventricular ectopy occurred in conjunction with myocardial injections in all NHPs undergoing treatment; however, all of the arrhythmias responded to lidocaine administration and had converted to sinus by the end of the anesthetic protocol. Histopathological analysis of H&E and trichrome stained cardiac sections revealed patchy cardiomyocyte degeneration and necrosis with mononuclear cell infiltration and fibrosis at the injection sites (Fig. 3). Finally, no significant alterations in baseline values of liver-function studies, complete blood count, or creatinine were noted (data not shown).
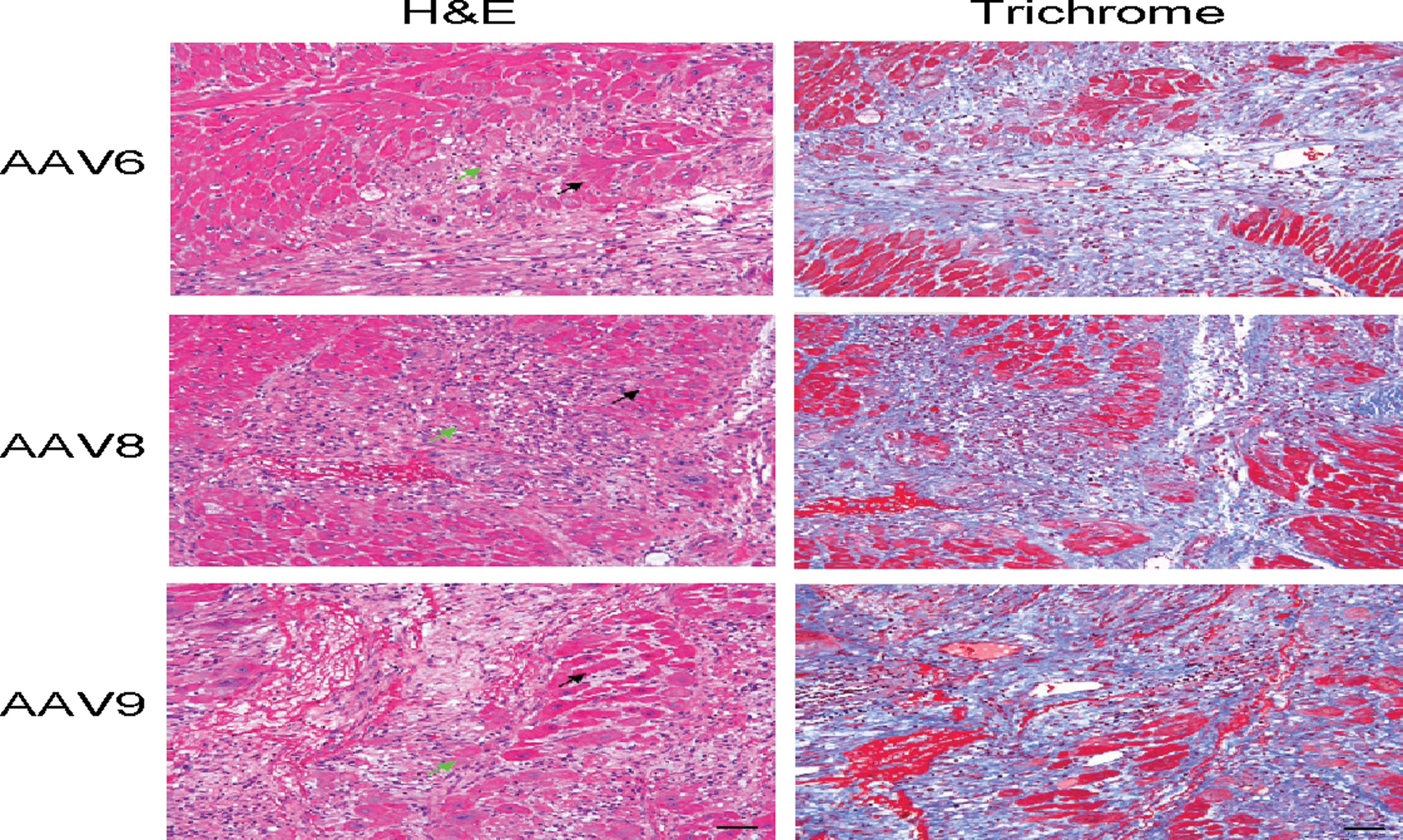
Representative high-power magnification images (20× objective) from NHP cardiac cryosections 7 days following injection with AAV. H&E staining demonstrates patchy cardiomyocyte degeneration (black arrow) and necrosis (green arrow) with mononuclear cell infiltration at the injection site. Trichrome staining reveals fibrosis at the injection site. Scale bar=80 μm. Color images available online at
Discussion
To our knowledge, this is the first investigation to compare the cardiac tropism of several AAV serotypes in the adult rhesus macaque. We previously developed a method to deliver vector to canine hearts via a percutaneously inserted injected catheter (Bish et al., 2008, 2011), and we applied a modified version of the technique in this report, where we compared the cardiac gene transfer efficiencies of AAV6, AAV8, and AAV9. We found that (i) transendocardial delivery of AAV is safe in the NHP, (ii) AAV6 and AAV8 provide efficient cardiac gene transfer at similar levels and are superior to AAV9, and (iii) AAV6 is more cardiac-specific than AAV8 and AAV9. It should be noted that the AAV8 data are based on n=1 and should be considered preliminary. Future investigation will be directed at increasing the number of NHPs treated with AAV8.
Currently, there are several ongoing clinical trials evaluating the therapeutic efficacy of cardiac gene transfer as a novel treatment for heart failure; the two vector delivery methods being used in these trials include coronary perfusion and direct transendocardial injection (Chaanine et al., 2010). Based on this, we believe that the transendocardial delivery method used in our current NHP study has the potential for clinical translation. Furthermore, we believe that direct transendocardial injection is a more versatile delivery method than coronary perfusion, because it could be used in both nonischemic and ischemic cardiomyopathy. Coronary perfusion would have limited utility in ischemic heart disease, because blood flow would be limited in the targeted areas of the heart. In addition, although coronary perfusion should theoretically deliver vector globally throughout the heart, histological analysis in large-animal models demonstrates that delivery is somewhat heterogeneous (Kawase et al., 2011), similar to what has been reported following direct transendocardial injection (Bish et al., 2011). Finally, vector delivered via direct transendocardial injection is less likely to be neutralized by pre-existing NAbs than vector delivered via coronary perfusion. Indeed, heart failure patients with an NAb titer of 1:2 did not respond to coronary perfusion of AAV-SERCA2a (Jaski et al., 2009), whereas we previously observed robust transgene expression following direct transendocardial injection of AAV-GFP in canines with NAb titers ranging from 1:20 to 1:40 (Bish et al., 2008). In summary, we believe that both coronary perfusion and direct transendocardial injection are delivery methods with potential clinical utility and that future investigation should be directed at comparing both methods in an NHP model.
The direct transendocardial injection procedure was well-tolerated in NHPs, just as in canines (Bish et al., 2008). The one death that occurred in postprocedure recovery was determined to be secondary to esophageal intubation, not the injection procedure itself. The only adverse event associated with the procedure was ventricular ectopy, which was amenable to lidocaine in all cases. Moreover, in all cases, ventricular ectopy had resolved before extubation. All animals that recovered from anesthesia survived until scheduled euthanasia at the 1-week time point. Pathology at this time revealed mononuclear infiltration and fibrosis at the injection sites, similar to what has been observed previously in the canine (Bish et al., 2008) and pig (Su et al., 2006). Unfortunately, no long-term safety data are available, because it was necessary to euthanize the animals at an early time point before a T-cell response was mounted against the cells expressing GFP, a concern raised by recent reports describing this phenomenon in baboon hearts (McTiernan et al., 2007), cynomolgus macaque livers (Gao et al., 2009), and canine livers (Bell et al., 2011) expressing a foreign protein. Although this time constraint did not allow us to analyze peak expression of the AAV vectors, we were still able to gain valuable preclinical information by comparing the relative cardiac tropism of each serotype. Indeed, we used this same strategy in our recent study that compared the cardiac tropism of the same AAV serotypes in the canine (Bish et al., 2008). Future investigation using a nonimmunogenic transgene will be necessary to address long-term expression and safety.
We have also identified AAV6 and AAV8 as vectors with high cardiac tropism in the macaque heart. AAV6 has been previously identified as an efficient cardiac gene transfer vector in several large-animal models, including the canine (Bish et al., 2008), pig (Sasano et al., 2007; Raake et al., 2008), and sheep (White et al., 2011), but this is the first time that AAV6, as well as AAV8, has been evaluated in the NHP. Previous NHP studies have identified AAV9 as superior to AAV5 (Tarantal and Lee, 2010) and AAV1 (Pacak et al., 2006) and equivalent to AAV10 (Tarantal and Lee, 2010) in the heart. We did not evaluate AAV10 in this study, but it is likely that as both AAV6 and AAV8 outperformed AAV9, they would also outperform the equivalent AAV10. We also did not evaluate AAV1 in this study. Although AAV1 is the serotype that has been used in initial clinical trials evaluating the efficacy of gene transfer of SERCA2a for heart failure (Jaski et al., 2009; Chaanine et al., 2010), at least two ongoing clinical trials are using AAV6 (Chaanine et al., 2010). In addition, it has been previously reported that AAV9 is superior to AAV1 in the NHP heart (Pacak et al., 2006), but evaluation of AAV6 in the NHP was lacking. Based on these facts, we decided to compare AAV6 and AAV9 in this study. However, as both AAV1 and AAV6 have been used in clinical trials, future investigation should be directed at comparing AAV1 and AAV6 in the NHP heart.
We also performed analysis of vector genome biodistribution to characterize the cardiac specificity of AAV6, AAV8, and AAV9. Although AAV6 and AAV8 achieved similar levels of cardiac GFP expression, the biodistribution profiles of the two vectors were very different. Vector genomes were detected at high levels in extracardiac tissues, especially the liver, following delivery of AAV8 in comparison with AAV6. For example, the ratio of vector genomes detected in the heart versus liver was 0.540 for AAV6 and 0.009 for AAV8—an approximately 50-fold difference. This cardiac tropism may be explained by enhanced cellular internalization and nuclear uncoating of AAV6 versus other serotypes in cardiomyocytes (Sipo et al., 2007). These results suggest that if cardiac specificity is a high priority, AAV6 should be the vector of choice over AAV8. If desired, cardiac specificity could be further enhanced by transcriptional, posttranscriptional, or transductional targeting (Muller et al., 2006; Xie et al., 2011), and future investigation is necessary to explore these options in the NHP model.
In summary, we have demonstrated that transendocardial delivery of AAV vectors is safe in the NHP. We are the first group to report that AAV6 and AAV8 are superior to AAV9 in the NHP heart and that AAV6 is more cardiac-specific than AAV8 and AAV9. The results of this study may be used to guide development of clinical applications of AAV vectors in the treatment of cardiovascular diseases.
Footnotes
Acknowledgments
This work was supported by the Parent Project Muscular Dystrophy (to H.L.S.), by the International Collaborative Effort (ICE) for DMD (to H.L.S.), and by the Natural Science Foundation of China (NSFC-30870734) (to Y.L.). We thank the Vector Core of the Children's Hospital of Philadelphia and Katherine High, M.D., for vector production.
Author Disclosure Statement
G.G. is an inventor on patents that have been licensed to various biopharmaceutical companies. No competing financial interests exist for L.T.B., M.M.S., L.S., Y.L., J.D., C.H., L.W., H.L.S., and X.M.