
Abstract
Sarcoma at advanced stages remains a clinically challenging disease. Interferons (IFNs) can target cancer cells by multiple antitumor activities, including the induction of cancer cell death and enhancement of immune response. However, the development of an effective cancer immunotherapy is often difficult, because cancer generates an immunotolerant microenvironment against the host immune system. An autologous hematopoietic stem cell transplantation (HSCT) is expected to reconstitute a fresh immune system, and expand tumor-specific T cells through the process of homeostatic proliferation. Here we examined whether a combination of autologous HSCT and IFNs could induce an effective tumor-specific immune response against sarcoma. First, we found that a type I IFN gene transfer significantly suppressed the cell growth of various sarcoma cell lines, and that IFN-β gene transfer was more effective in inducing cell death than was IFN-α in sarcoma cells. Then, to examine the antitumor effect in vivo, human sarcoma cells were inoculated in immune-deficient mice, and a lipofection of an IFN-β-expressing plasmid was found to suppress the growth of subcutaneous tumors significantly. Finally, the IFN gene transfer was combined with syngeneic HSCT in murine osteosarcoma models. Intratumoral IFN-β gene transfer markedly suppressed the growth of vector-injected tumors and inhibited formation of spontaneous lung and liver metastases in syngeneic HSCT mice, and an infiltration of many immune cells was recognized in metastatic tumors of the treated mice. The treated mice showed no significant adverse events. A combination of intratumoral IFN gene transfer with autologous HSCT could be a promising therapeutic strategy for patients with sarcoma.
Introduction
Soft-tissue sarcoma also represents a rare entity of malignant tumors of various histologies. The cure of early disease depends on the appropriate multidisciplinary therapy of the primary tumor, including surgery and radiotherapy (Sarcoma Meta-analysis Collaboration, 1997). Adjuvant chemotherapy has not been shown to improve overall survival significantly, and chemotherapeutic interventions in metastatic soft-tissue sarcoma were generally disappointing (Borden et al., 1990; Antman, 1997). New therapeutic approaches are needed in the therapeutic strategy of malignant bone and soft-tissue sarcoma.
The type I interferon (IFN) is a pleiotropic cytokine that can induce antiproliferation, cancer cell death, antiangiogenesis, and immunomodulation, and has been used for the treatment of a variety of cancers, including chronic myelogenous leukemia, melanoma, and renal cancer (Pfeffer et al., 1998; Belardelli et al., 2002). In terms of sarcoma, Müller and co-workers suggested the efficacy of IFN as adjuvant treatment for a high-grade osteosarcoma (Muller et al., 2005). However, clinical experiences with IFN protein therapy for solid tumors have not been encouraging in most of the cases (Einhorn and Grander, 1996). In the conventional regimen of IFN clinical trials, the recombinant IFN protein is systemically administered through subcutaneous or intramuscular routes. As the protein is rapidly degraded in the blood circulation and only a small portion of subcutaneously injected IFN can reach the target sites (Salmon et al., 1996), the limited therapeutic efficacy of treatments with IFN protein reflects the inability to target the cytokine to the right place at a sufficiently high dose. Alternative delivery strategies are needed to achieve a safe and effective IFN delivery. Several animal studies demonstrated that gene- and cell-based delivery of type I IFNs into tumors suppressed the growth of various cancers, such as breast and prostate cancers (Zhang et al., 1996; Hottiger et al., 1999; Ahmed et al., 2001; Benedict et al., 2004; Studeny et al., 2004; De Palma et al., 2008).
Although type I IFN was long thought to act mainly through the direct suppression of tumor cell proliferation in vivo, it has recently been established that IFNs have important roles in regulating the innate and adaptive arms of the immune system: up-regulation of major histocompatibility complex (MHC) class I gene, promotion of the priming and survival of T cells, increase of the cytotoxic activity of natural killer (NK) cells and CD8+ T cells, and activation of dendritic cells (DCs) (Ferrantini et al., 2008; Santini et al., 2009). We also reported that, in addition to the direct cytotoxicity at the injected site, an intratumoral IFN gene transfer elicits a systemic tumor-specific immunity in several animal models (Hatanaka et al., 2004; Ohashi et al., 2005; Hara et al., 2007, 2009).
However, the development of effective cancer immunotherapy is often difficult in the clinical setting, because cancer generates an immunotolerant microenvironment against the host immune system (Rabinovich et al., 2007); an additional therapeutic strategy is required to bring a breakthrough. Hematopoietic stem cell transplantation (HSCT) following a preconditioning is expected to introduce a fresh immune system, in which tolerance to tumor cells is not yet induced, and may present an opportunity to augment the efficacy of IFN immune therapy. Furthermore, it has been reported recently that autologous HSCT can induce antitumor immunity by homeostatic proliferation (HP) of T cells following preconditioning-induced lymphopenia. It is known that T-cell HP is driven by the recognition of self antigens, and that the availability of tumor antigens during HP leads to effective antitumor autoimmunity with specificity and memory. The effect is mediated by a reduction in the activation threshold of low-affinity tumor-specific T cells, leading to their preferential engagement and expansion (Baccala et al., 2005; Wrzesinski and Restifo, 2005). Induction of lymphopenia can also enhance cytokine production or increase homeostatic cytokine levels [interleukin (IL)-7, IL-15] by removing cells that compete for these endogenous cytokines (Gattinoni et al., 2005; Wang et al., 2005). In fact, several animal studies showed that lymphopenic conditions alone are able to create an environment sufficient to mount an efficient antitumor immunity (Borrello et al., 2000; Dummer et al., 2002). In this study, we investigated whether a combination of syngeneic HSCT and intratumoral IFN gene transfer induces an effective antitumor immune response against sarcoma.
Materials and Methods
Tumor cell lines and plasmids
143B, U2OS, HOS, MG-63, and HuO 9N2 are human osteosarcoma cell lines. NHOS and LM8 are murine osteosarcoma cell lines, which were derived from BALB/c or C3H mice, respectively. SK-LMS-1, SK-UT-1B, and SKN are human leiomyosarcoma cell lines. SYO1 is a human synovial sarcoma cell line, HT-1080 is a human fibrosarcoma cell line, and RD is a human rhabdomyosarcoma cell line. 143B, U2OS, HOS, and SYO1 cells were maintained in Dulbecco's modified Eagle's medium containing 10% fetal bovine serum (FBS; ICN Biomedicals, Inc., Irvine, CA). HuO 9N2 and NHOS were maintained in RPMI-1640 containing 10% FBS. MG-63, SK-LMS-1, SK-UT-1B, HT-1080, and RD were maintained in Eagle's minimal essential medium containing 10% FBS. SKN was maintained in Ham's F12 medium with 10% FBS. An LM8 cell line that stably expresses the firefly luciferase gene was generated by retrovirus vector-mediated transduction and designated as LM8-Luc.
Plasmids expressing the IFN-α or IFN-β gene under the control of the Rous sarcoma virus promoter were also used for intratumoral gene transfer. The human IFN-α- or IFN-β-expressing plasmids were designated as phIFN-α or phIFN-β, respectively. The mouse IFN-α- or IFN-β-expressing plasmids were designated as pmIFN-α or pmIFN-β, respectively. The plasmids that express an enhanced green fluorescent protein (pEGFP) and luciferase (pLuc) were used as a negative control.
In vitro IFN gene transfer and cell-proliferation assay
Culture cells were seeded at 3–8×103 cells/well in 96-well plates 1 day before transfection, and 0.12 μg of plasmid DNA-liposome (Lipofectamine 2000; Invitrogen, Carlsbad, CA) complex was added into each well in accordance with the manufacturer's protocol. The cell numbers were assessed by a colorimetric cell viability assay using a water-soluble tetrazolium salt (Tetracolor One; Seikagaku Corp., Tokyo, Japan). The absorbance was determined by spectrophotometer using a wavelength of 450 nm with 600 nm as a reference. The growth suppression was expressed as the relative cell growth, which was calculated as OD450 of pIFN-α- or pIFN-β-transfected cells/OD450 of pEGFP-transfected cells. The assays (carried out in six wells) were repeated three times.
Annexin V assay
Cells were seeded at 5–10×104 cells/well in six-well plates 1 day before transfection. A 2-μg plasmid DNA-liposome complex was added to each well. Three days later, the cells were collected and stained with Annexin V-FITC (Medical & Biological Laboratories Co., Ltd., Nagoya, Japan), which detects phosphatidylserine of inverted plasma membranes. The cells were examined by fluorescence-activated cell sorting (FACSCalibur; BD Biosciences, San Jose, CA). The apoptotic cell death was expressed as specific cell death, which was calculated as the cell death fraction induced by phIFN or pmIFN (%) minus that by pLuc (%). The measurement of the wells was carried out in triplicate, and the means±SD was plotted. The assay was repeated two times.
Cell-cycle analysis
Cells were seeded at 5–10×104 cells/well in six-well plates 1 day before transfection. A 2-μg plasmid DNA-liposome complex was added to each well, and 3 days later the cells were stained with propidium iodide (PI; Sigma, St. Louis, MO), followed by cell-cycle analysis with flow cytometry. The percentage of the subG1 fraction, which indicates the cell-death population, was calculated compared with the total of the subG1, G0/G1, S, and G2/M phases. A fraction of subG1 shows a major population of cell cycles in some cell lines, which obscures the effect of type I IFN on cell-cycle arrest. Therefore, the percentages of the G0/G1, S, and G2/M fractions were calculated compared with the total of the G0/G1, S, and G2/M fractions except for the subG1 fraction. Then, to demonstrate how the cell cycle is affected by type I IFN expression but not plasmid DNA transfection per se, we subtracted the percentages of each cell-cycle phase in pLuc-transfected cells from those in phIFN-α- or phIFN-β-transfected cells, and expressed them as the relative fraction of cells in the cell-cycle phases. The assays were carried out in triplicate.
Animals and IFN-β gene transfer
The sarcoma cells (5–10×106) were injected subcutaneously on the legs of BALB/c nude mice (Charles River Japan, Kanagawa, Japan). When the subcutaneous tumor was established (∼0.7 cm in diameter), 150 μl of plasmid DNA-liposome complexes was injected directly into the tumors three times every other day. The plasmid DNA-liposome complex was prepared by the addition of 30 μg of plasmid DNA into a total of 75 μl of PBS per mouse, followed by the addition of 75 μl of 0.15 mmol/L DMRIE-DOPE [(±)-N-(2-hydroxyethyl)-N, N-dimethyl-2,3-bis(tetradecyloxy)-1-propanaminium bromide/dioleoylphosphatidylethanolamine] (Vical, Inc., San Diego, CA). The shortest (r) and longest (l) tumor diameters were measured at the indicated days, and the tumor volume was determined as r 2×l/2. Tumor sizes (means±SD) were measured on the days indicated.
For NK cell-depletion experiments, we started the intraperitoneal injection (50 μl) of anti-asialo GM1 antibody (30 mg/ml; Wako Chemicals USA Inc., Richmond, VA) 2 days before the plasmid/liposome administration; the treatment was repeated every 6 days. Animal studies were carried out according to the Guideline for Animal Experiments of the National Cancer Center Research Institute and approved by the Institutional Committee for Ethics in Animal Experimentation.
Hematopoietic stem cell transplantation
Seven- to 9-week-old female BALB/c or C3H mice were purchased from Charles River Japan. The mice received a lethal (9 Gy) irradiation on the day of transplantation, and the irradiated mice were injected intravenously with 5×106 bone marrow (BM) cells and 2×106 splenic T cells from syngeneic donor mice. BM cells were isolated from donors by flushing each femur and tibia with an RPMI-1640 medium supplemented with 5% FBS (complete RPMI), and splenic cells were prepared by macerating the spleens. After lysis of the erythrocytes, the splenic cells were incubated with anti-Thy-1.2 immunomagnetic beads (Miltenyi Biotec GmbH, Bergisch Gladbach, Germany) at 4°C for 15 min, followed by selection of T cells by AutoMACS (Miltenyi Biotec). Simultaneously, NHOS (3×106) or LM8-Luc (5×105) cells were injected subcutaneously into the legs of BALB/c or C3H mice, respectively.
ELISpot assay
An IFN-γ ELISpot kit (BD Bioscience) was used according to the manufacturer's instructions. In brief, splenocytes (1×105 cells/well) and mitomycin C (MMC)-treated tumor cells (1×104 cells/well) were cocultured in 96-well plates precoated with mouse IFN-γ (BD Bioscience) for 20 hr at 37°C in complete RPMI medium in triplicate. MMC was used to prevent proliferation of tumor cells in the culture with lymphocytes. After the wells were washed, a biotinylated anti-mouse IFN-γ antibody (2 μg/ml) was added and incubation continued for 2 hr at room temperature. Then a streptavidin–horseradish peroxidase solution was added and incubation continued for 1 hr at room temperature. After the addition of an aminoethylcarbazole substrate solution, spots were counted under a stereomicroscope.
Cytotoxic assays
The splenocytes were cultured for 4 days with MMC-treated NHOS stimulators, and then the responder cells were collected and used as effector cells. NHOS target cells were labeled with 51Cr (PerkinElmer Japan Co., Kanagawa, Japan). For a 4-hr chromium release assay, 2×105, 1×105, 5×104, and 2.5×104 effector cells were mixed with 0.5×104 target cells per well in a 96-well round-bottom plate (Corning Inc., Corning, NY). Supernatants were harvested and counted in a gamma counter (Packard Bioscience Company, Meriden, CT). The percentage of cytotoxicity was calculated as [(experimental cpm – spontaneous cpm)/(maximum cpm – spontaneous cpm)]×100. Each assay was done in triplicate.
In vivo imaging of tumor metastasis
The C3H mice with LM8-Luc tumors were administered D-luciferin (150 mg/kg) (Wako Pure Chemical Industries) by intraperitoneal injection. Ten minutes later, photons from animal whole bodies were counted using an in vivo imaging system (IVIS; Xenogen, Alameda, CA).
Immunohistochemistry and TUNEL assay
Immunostaining was performed using the streptavidin–biotin–peroxidase complex techniques (Nichirei, Tokyo, Japan). Consecutive cryostat tissue sections (6 μm) were mounted on glass slides and fixed in 99.5% ethanol for 20 min. After blocking with normal rat serum, the sections were stained with rat anti-mouse CD4 and CD8 antibodies (BD Biosciences). The cryostat sections were also processed for the terminal deoxynucleotidyltransferase-mediated dUTP-digoxigenin nick-end-labeling (TUNEL) assay (ApopTag in situ apoptosis detection kit; Intergen Company, Purchase, NY). Negative controls without primary antibodies were examined in all cases. The sections were counterstained with methyl green.
Statistical analysis
Two-sided t tests were used to validate the significance of the observed differences, which were considered statistically significant when p<0.05.
Results
Transfection of IFN plasmid suppressed cell growth
First, to examine the gene-transduction efficiency, a variety of sarcoma cell lines, one lung cancer cell line, and one prostate cancer cell line were transfected with pEGFP in six-well plates. Flow cytometry showed that the percentage of EGFP+ cells varied [1.6% (LM8) to 75.9% (HT1080)]; however, it was less than 20% in almost all cell lines except for two cell lines (Fig. 1A). Next, to compare the expression levels of IFN-α and IFN-β with the gene-transduction efficiency the IFN concentration was measured in the supernatants of IFN plasmid-transfected cells. The enzyme-linked immunosorbent assay (ELISA) showed that the IFN concentrations were also different; however, they appeared to be related to the gene-transduction efficiency (Fig. 1B). The IFN-α concentration was generally higher than the IFN-β concentration in each cell line (Fig. 1B). The IFN-α gene might be more effectively translated than the IFN-β, or the IFN-α protein might be more stable than IFN-β in the supernatant of the culture cells.
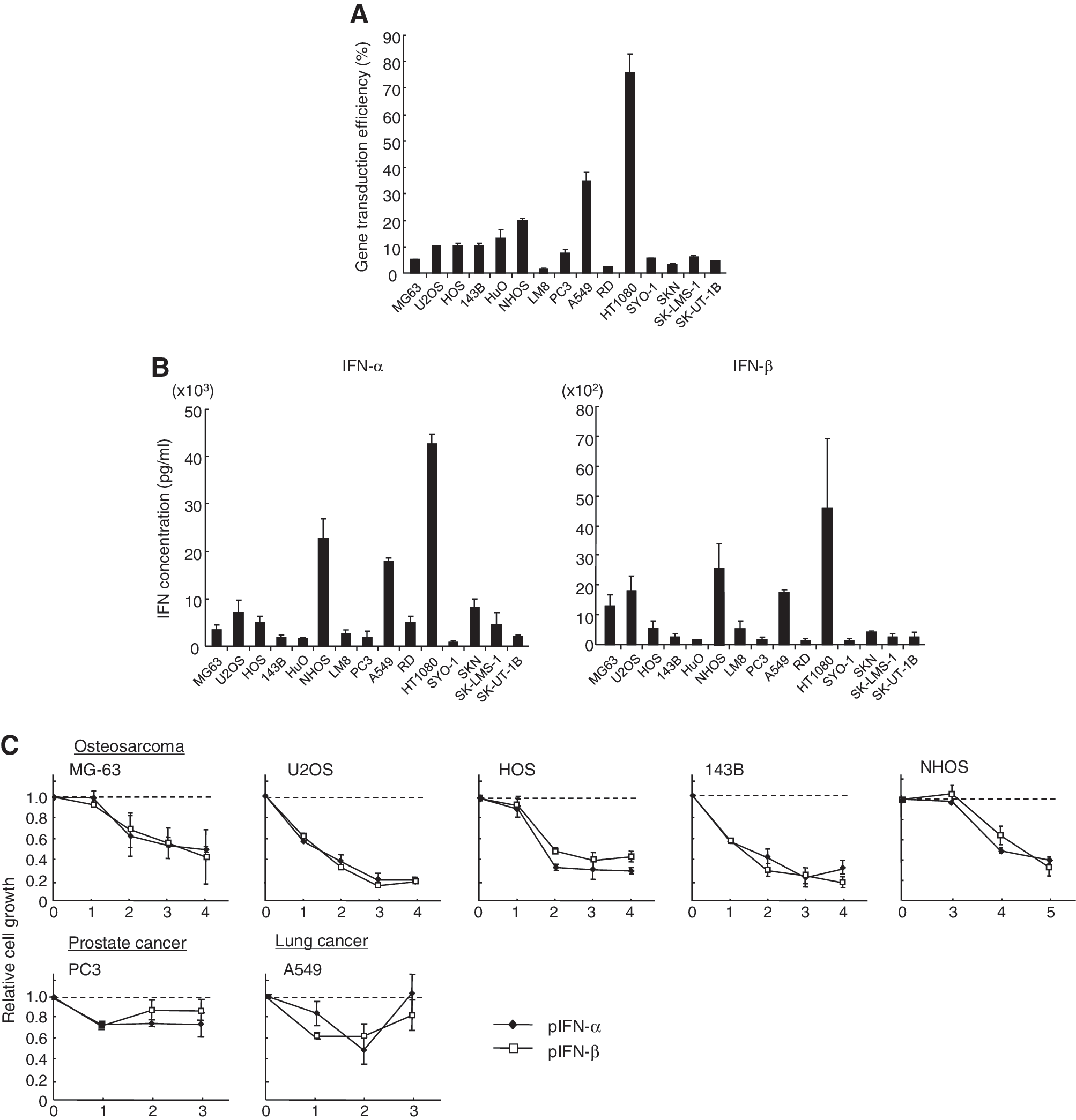
Cytotoxic effect of type I IFN gene transfer in various sarcoma cell lines.
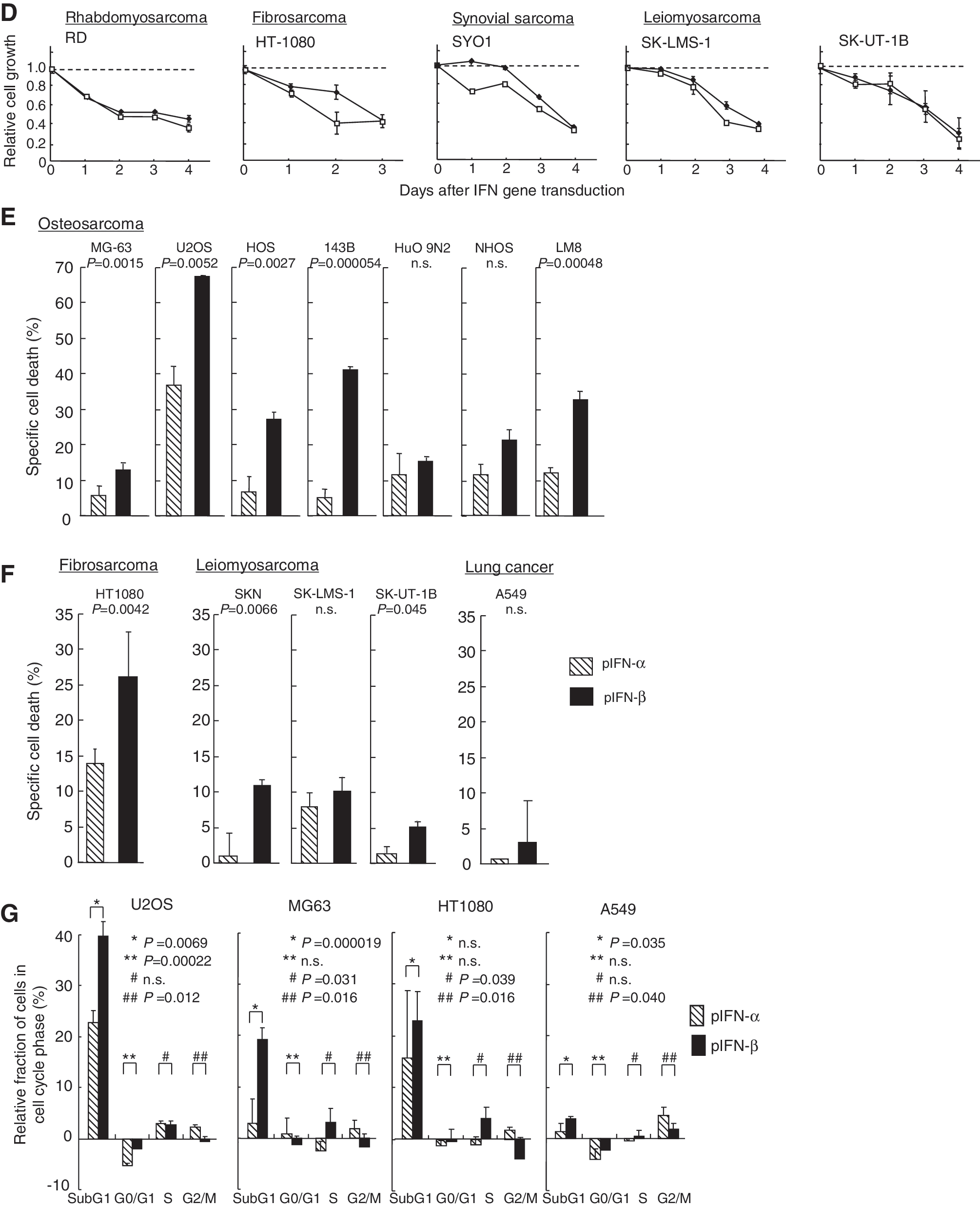
To examine whether the expression of IFN gene effectively inhibits the growth of osteosarcoma cells, four human osteosarcoma cell lines and one mouse osteosarcoma cell line (NHOS) were transfected with pIFN-α or pIFN-β. The transfection of the IFN plasmid inhibited the growth of all osteosarcoma cells to less than 40% of that of the control plasmid (pEGFP)-transfected cells, whereas the growth suppression was less conspicuous in the prostate (PC3) and lung (A549) cancer cells (Fig. 1C). Then, to investigate the growth-suppressive effect of IFN gene transduction in various human soft-tissue sarcoma cell lines, phIFN-α or phIFN-β was introduced to one rhabdomyosarcoma, one fibrosarcoma, one synovial sarcoma, and two leiomyosarcoma cell lines. IFN expression significantly suppressed cell growth in all sarcoma cell lines (Fig. 1D).
To examine cell-death induction by IFN gene transduction, Annexin V assay was performed in various sarcoma cell lines transfected with the IFN-α or IFN-β plasmid. The assay showed that IFN expression effectively induced apoptosis in many osteosarcoma cell lines and soft-tissue sarcoma cell lines. IFN-β gene transfer was more effective in inducing cell death than IFN-α gene transfer in most sarcoma cell lines (Fig. 1E and F). Although the suppression of cell growth and induction of apoptosis were more effective in the sarcoma cells as compared with the prostate and lung cancer cells, it was not closely related to the IFN concentration; for example, the transfected A549 cells produced a relatively high IFN level, but the apoptosis induction was less conspicuous, suggesting that sarcoma cell lines are susceptible to type I IFN expression. The IFN concentration may not be increased in the IFN-sensitive cells due to the decrease of cell number by apoptosis induction.
Furthermore, to examine the effect of IFN gene transduction in the cell cycle, three sarcoma cell lines and one lung cancer cell line were transfected with phIFN-α or phIFN-β. Flow cytometry showed that the subG1 fraction was significantly increased in the IFN-β-transduced cells compared with the IFN-α-transduced cells (Fig. 1G), which confirmed the results of the Annexin V assay. In terms of the effect of IFN on the cell cycle, it has been reported that type I IFN induces G1 cell-cycle arrest in renal cancer cells (Shang et al., 2011) and G2/M arrest in hepatocellular carcinoma (Fujioka et al., 2006) and colon cancer cells (Barnes et al., 2003). In this study, IFN-α transduction increased the percentage of cells in a G2/M phase in sarcoma cell lines (Fig. 1G). The cell growth inhibition by IFN-α transduction was similar to IFN-β in the cell-proliferation assay (Fig. 1C and D). Although the cell-death induction was effectively induced by IFN-β, IFN-α also was able to significantly induce apoptosis, and the G2/M arrest by IFN-α transduction might add to the apoptosis induction.
Antitumor effect of intratumoral IFN-β gene transfer in subcutaneous tumor model
To examine the antitumor effect of IFN-β gene transfer in vivo, 143B human osteosarcoma or SK-UT-1B human leiomyosarcoma cells were inoculated on the legs of BALB/c nude mice, and then a phIFN-β-liposome complex was injected directly into the tumors three or six times. The growth of the 143B tumor was more rapid than that of the SK-UT-1B tumor. The IFN-β gene transfer showed a significant suppressive effect against the 143B tumors in a dose-dependent manner, whereas a three-time injection of phIFN-β-liposome was sufficient to eradicate the SK-UT-1B tumors (Fig. 2A). The ELISA showed that the IFN-β expression in the 143B subcutaneous tumors continued for more than 7 days after intratumoral injection of phIFN-β (30 μg) three times (Fig. 2B).
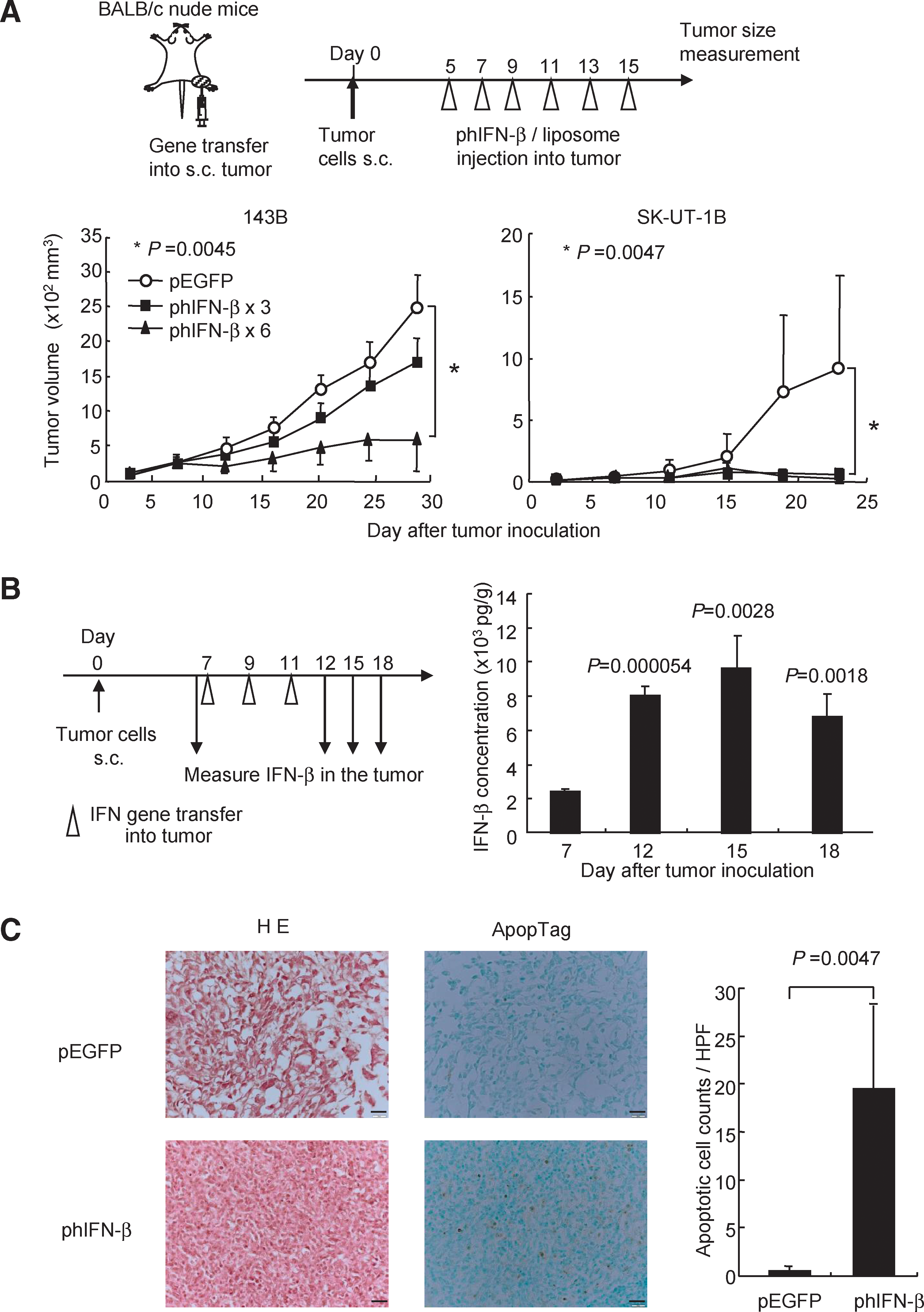
Growth suppression of human sarcoma xenografts by intratumoral IFN-β gene transfer.

Then, to examine whether the human IFN-β expression induces cell death in the tumors, the TUNEL assay was` performed using tumors of mice treated with human IFN-β gene transfer. The assay revealed the cell death of many cancer cells in the phIFN-β-transfected 143B tumors (Fig. 2C).
As mouse IFN shows no cross-species activity in human cells, it does not directly induce cell death in 143B or SK-UT-1B tumors, whereas mouse, but not human, type I IFN can enhance the cytotoxicity of mouse immune cells, such as NK cells (Ohashi et al., 2005). To examine whether mouse IFN-β gene therapy leads to suppression of human osteosarcoma xenografts in nude mice, 143B or SK-UT-1B cells were inoculated on bilateral legs of BALB/c nude mice, and a pmIFN-β-liposome complex was injected directly into the tumors three times. The mouse IFN-β gene transfer significantly suppressed growth of not only the transfected right tumors, but also of the untransfected left tumors (Fig. 2D). The results indicated that an intratumoral IFN-β gene transfer effectively induces systemic antitumor immunity. To confirm that mouse IFN-β gene transfer activated the NK cells, the mice were treated with an anti-asialo GM1 antibody to purge the NK cells. Flow cytometry showed that the treatment with antibody depleted approximately 80% of NK (CD49b+) cells in the spleen (Fig. 2D). Depletion of NK cells canceled the antitumor effect of IFN-β gene transfer and rather increased tumor volume as compared with the control pEGFP-injected group (Fig. 2D); this suggests that NK cells in nude mice have a significant activity to suppress the tumor growth of sarcoma cells and that an intratumoral IFN gene transfer enhances the suppressive effect of NK cells.
Antitumor effect of syngeneic HSCT and intratumoral type I IFN gene transfer against osteosarcoma
To examine whether the antitumor immunity induced by intratumoral type I IFN gene transfer could be enhanced by HP of T cells in lymphopenic hosts, BALB/c mice were injected subcutaneously with NHOS osteosarcoma cells shortly after lethal irradiation, and syngeneic BM and T cells were infused into the mice. Tumor growth was significantly suppressed in the syngeneic HSCT recipients as previously reported (Kobayashi et al., 2007). Furthermore, a combination of intratumoral IFN-α or IFN-β gene transfer and syngeneic HSCT enhanced the antitumor effect against osteosarcoma (Fig. 3A). The activity to induce the antitumor immunity by IFN-α gene transfer may be compatible with that by IFN-β for NHOS tumors. All treated mice appeared healthy during the course of the experiments, and the blood chemistry showed no abnormal values in the treated mice 5 weeks after the HSCT (data not shown).
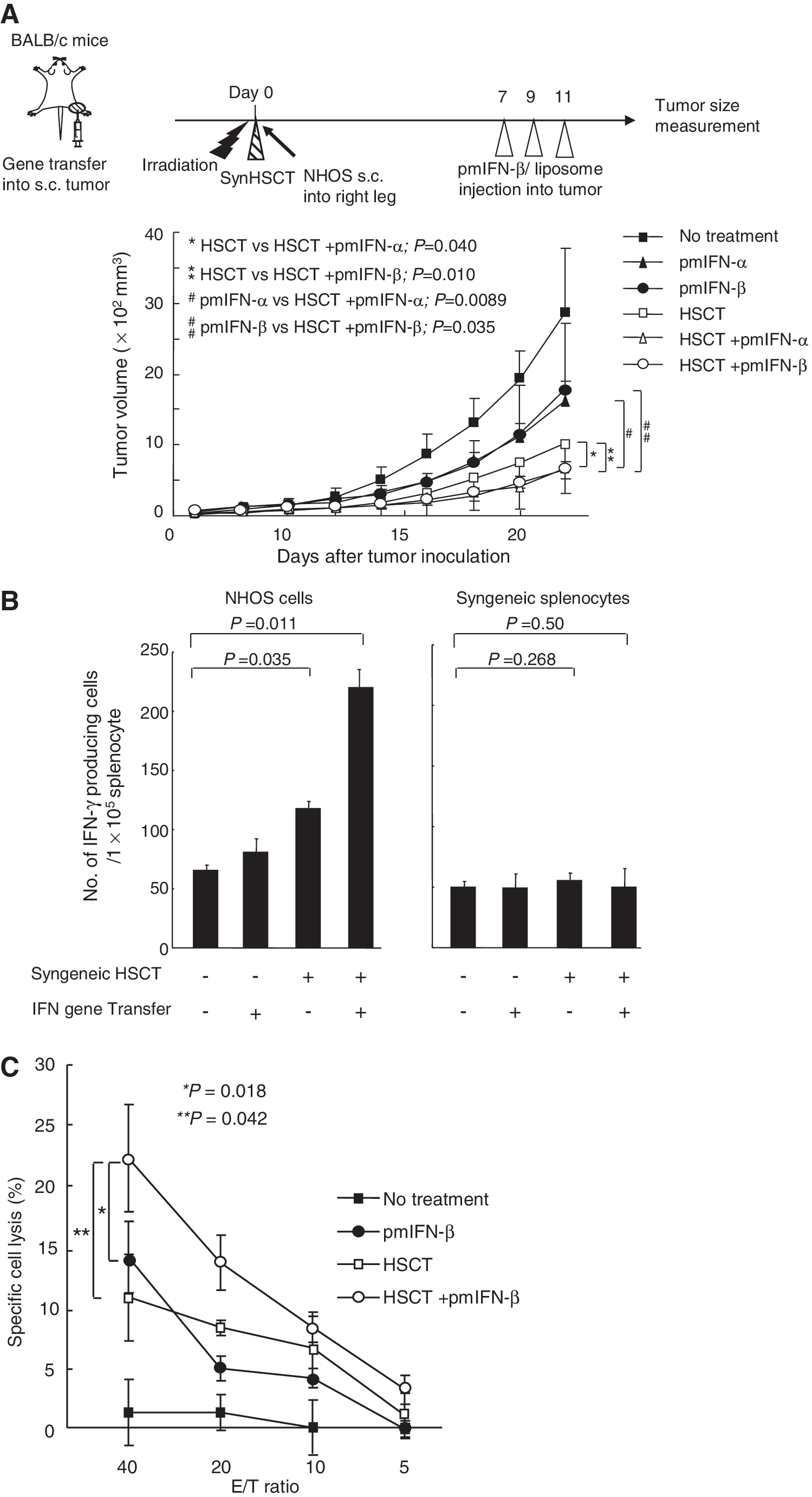
Type I IFN gene transfer induces a significant antitumor effect in syngeneic HSCT recipients.
To examine the immune reaction to intratumoral IFN-β gene transfer in syngeneic HSCT recipients, splenocytes were extracted from the treated mice and cultured with NHOS cells. An ELISpot assay showed that the average number of IFN-γ-producing splenocytes in response to NHOS cells was slightly increased in the syngeneic HSCT alone group, whereas a combination of intratumoral IFN-β gene transfer and syngeneic HSCT further increased the IFN-γ+ spots. The spot number in response to control syngeneic splenocytes was not changed in all groups of treated mice (Fig. 3B). An in vitro cytotoxic assay also showed that the splenocytes derived from the syngeneic HSCT mice recognized and lysed NHOS cells, and that the IFN-β gene transfer enhanced the cytolysis to NHOS cells (Fig. 3C).
Inhibition of distant metastases by a combination of IFN-β gene transfer and syngeneic HSCT
Lung and liver metastases are among the most frequent causes of mortality in patients with malignant bone and soft-tissue sarcoma. LM8 is a highly metastatic osteosarcoma cell line derived from C3H mouse. Many macroscopic metastatic foci were observed in the liver and lung 20 days after the subcutaneous inoculation of LM8 cells in C3H mice. To evaluate the therapeutic efficacy of IFN-β gene transfer against distant metastases in syngeneic HSCT recipients, LM8-Luc cells, which express the firefly luciferase gene, were inoculated on the right legs of C3H mice after syngeneic HSCT, and 7 days later, a pmIFN-β-liposome complex was injected into the subcutaneous tumor on the leg three times. Intratumoral IFN-β gene transfer alone or syngeneic HSCT alone significantly suppressed tumor growth of subcutaneous tumors on the legs; however, the antitumor effect of an IFN-β gene transfer was clearly enhanced in syngeneic HSCT recipients (Fig. 4A), which was similar to the antitumor effect against NHOS tumors (Fig. 3A).
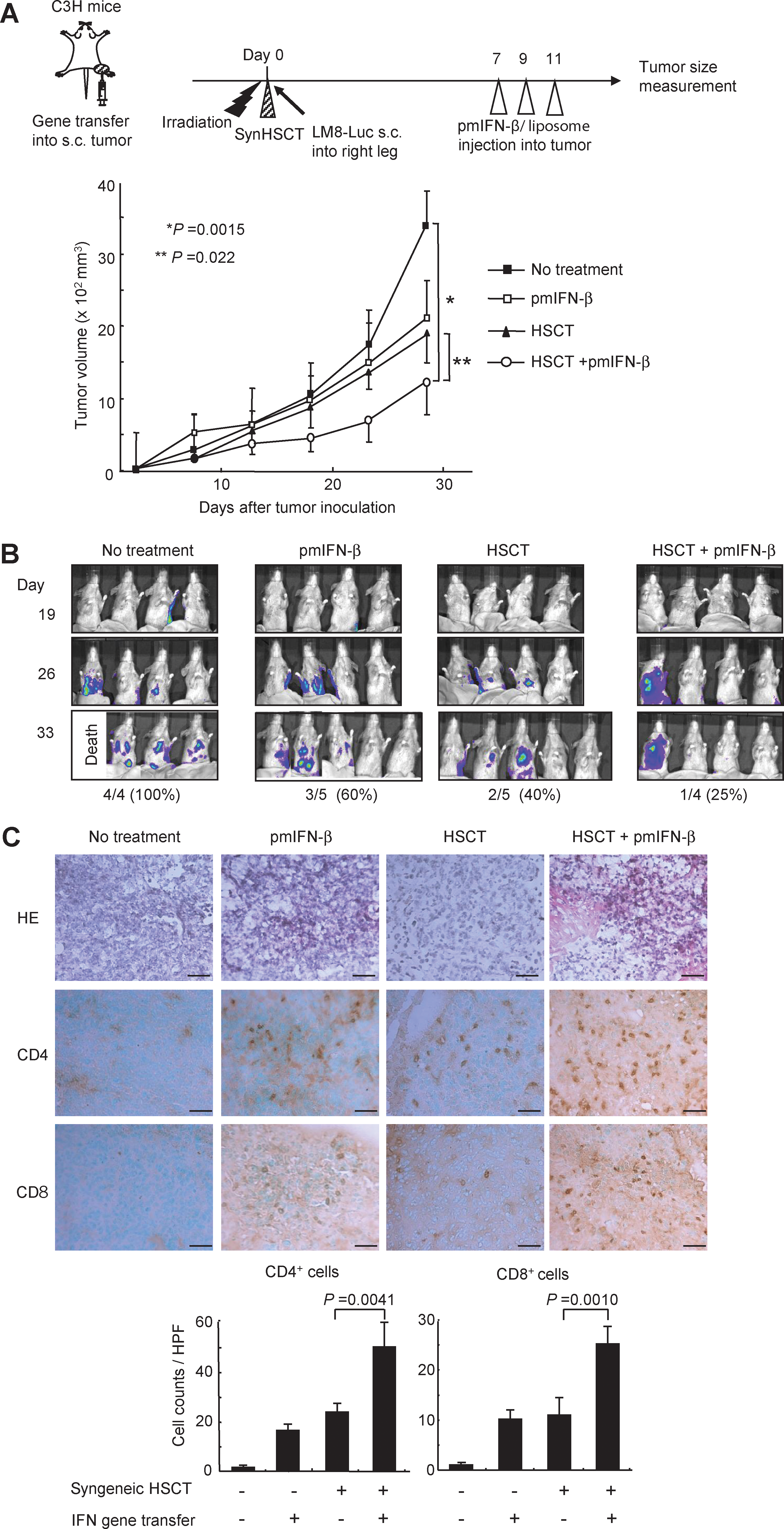
Suppression of tumors at a distant site by IFN-β gene transfer during immune reconstitution.
To examine the suppressive effect for the formation of metastatic tumors, distant metastases in the lung and liver were evaluated by photon counts on the IVIS imaging system in the treated mice (Fig. 4B). Thirty-three days after tumor inoculation, all of the four untreated control mice showed many photon+ spots (one of the four died of cancer progression), whereas three (75%) of the four mice treated with a combination therapy showed no detectable metastatic spots in the lung and liver.
The infiltration of CD4+ and CD8+ T cells was analyzed in the lung tumors of treated mice at day 33 after tumor inoculation. Immunohistochemical staining showed that CD4+ and CD8+ T cells infiltrated into the lung metastases in the group of intratumoral IFN-β gene transfer alone and that of syngeneic HSCT alone, whereas a combination therapy markedly increased the number of these cells in the lung tumors (Fig. 4C). These results indicated that a combination therapy is effective for preventing distant metastases in the liver and lung.
Discussion
In the first part of this study, we showed that type I IFN gene transduction suppressed the cell growth of various sarcoma cells, and that the IFN-β gene was more effective in inducing cell death than was the IFN-α gene. In vivo delivery of the IFN-β gene significantly suppressed the growth of osteosarcoma tumors. Then we demonstrated that the integration of two complementary immune therapies, intratumoral IFN gene transfer (which induces tumor-specific immunity) and HSCT (which reconstructs a fresh immune system), significantly enhances the antitumor immunity against osteosarcoma. An autologous HSCT has obvious advantages over allogeneic HSCT, such as a lack of graft-versus-host disease and an independence of donor availability.
Although the underlying mechanisms for the effective induction of cell death by IFN gene transfer in sarcoma cell lines are not clear, it has been reported that the addition of type I IFN protein showed a substantial inhibitory effect on the in vitro growth of osteosarcoma cells, as well as various soft-tissue sarcoma cells, and that the administration of IFN protein was also effective in arresting the growth of four different human osteosarcoma xenografts in nude mice (Thulasi et al., 1996; Brodowicz et al., 1999; Strander, 2007; Whelan et al., 2010), suggesting that a sensitivity to type I IFN protein is one of the characteristics of sarcoma cells. Furthermore, Yoshida and co-workers reported that IFN-β gene transfer could induce apoptosis in IFN protein-resistant tumor cells, such as glioma, melanoma, and renal cell carcinoma (Yoshida et al., 2004), demonstrating that an IFN gene transfer has an obvious advantage over IFN protein treatment in the induction of cell death at least in certain types of tumor cells. Sarcoma could be a promising target of IFN gene therapy.
In addition to cell-death induction, IFN gene transfer has an ability to mount an antitumor immunity against osteosarcoma, and this antitumor efficacy was significantly enhanced in syngeneic HSCT recipients (Figs. 3A and 4A). A putative mechanism of inducing an antitumor immunity by a combination therapy is as follows: in the “homeostatic proliferation” condition after syngeneic HSCT, T cells effectively recognize the low-affinity self antigen, including tumor-associated antigens (TAAs), leading to an induction of antitumor immunity. IFN-α/β expression in the tumors causes tumor lysis and exposes an increased quantity of TAAs to DCs. IFNs promote maturation and enhance the antigen-presenting capacity of DCs (Hara et al., 2009), leading to the facilitation of TAA recognition by donor lymphocytes primed during homeostatic proliferation. Furthermore, it was reported that type I IFNs augment the cytotoxicity of T cells and NK cells and enhance in vivo proliferation, expansion, and long-term survival of CD8+ T cells in response to specific antigens (Ferrantini et al., 2007; Santini et al., 2009). Last but not least, conditioning of HSCT (by irradiation in our model) destroys the immunotolerance microenvironment developed by sarcoma, contributing to an effective antitumor immunity (our unpublished data). These multiple and supposedly synergistic mechanisms may lead to the development of a strong antitumor effect against sarcoma. The combination of immunostimulatory effects by IFN and the reconstitution of a fresh immune system following HSCT could create an environment mutually supporting the activation of an antitumor response.
The antitumor immunity of gene- and cell-based immunotherapy enhanced by syngeneic HSCT has been shown in a variety of animal models and clinical studies. An adoptive transfer of tumor-specific T cells showed a strong antitumor effect in syngeneic HSCT mice (Wrzesinski and Restifo, 2005). In clinical trials, host lymphodepletion followed by autologous tumor-infiltrating lymphocyte transfer and IL-2 administration results in objective response rates of 50–70% in patients with metastatic melanoma refractory to standard therapies (Dudley et al., 2008). As tumor-reactive T cells are mostly polyclonal, and heterogeneous expressions of various TAAs coexist in a tumor mass, the in vivo stimulation of multiple tumor-reactive lymphocytes might be critical in the clinical application. Therefore, several laboratories reported that a vaccination with tumor cells expressing immunostimulatory molecules and an immunization with DCs pulsed with whole-tumor cell lysates led to an efficient antitumor response early after syngeneic HSCT (Borrello et al., 2000; Asavaroengchai et al., 2002; Jing et al., 2007; Filatenkov et al., 2009).
Compared with the previous approaches, one of the advantages of the in vivo IFN gene transfer is that it does not involve a manipulation and culture of the immune and tumor cells ex vivo, making this strategy more feasible for patients with bone and soft-tissue sarcomas. We previously reported that an allogeneic MHC gene transfer could also enhance an effective antitumor immunity in syngeneic HSCT recipients (Kobayashi et al., 2007). The major difference between the allogeneic MHC and IFN gene therapies is in their local effects on tumor sites transduced with the therapeutic genes: IFN gene transfer directly induces cell death, which may enable local control and increased release of TAAs.
The current and previous studies address important points to consider in the clinical feasibility of the combination therapy. First, a liposome-mediated IFN gene transfer effectively suppressed tumor growth in syngeneic HSCT recipients, suggesting that a high concentration of IFN in the tumors is not necessary to induce an effective antitumor immunity. This facilitates a clinical application, because in vivo lipofection of an IFN-expressing plasmid is much safer than virus vectors (Rodriguez, 2004). Second, the intratumoral route of the vector administration has a better safety feature compared with an intravenous route, which may distribute and express the IFN gene in various organs. In addition to the lower toxicity, we reported that an intratumoral route of IFN gene transfer is superior to an intravenous administration due to the effective induction of antitumor immunity through the exposure of TAAs to DCs and the maturation of DCs in regional lymph nodes by the intratumoral IFN expression (Narumi et al., 2010). Third, a combination therapy was effective in suppressing not only the vector-injected tumors, but also the vector-uninjected distant tumors in a murine spontaneous metastasis model, simulating a clinical setting. On the other hand, remaining questions include further understanding of the kinetics of homeostatic proliferation, which is the basis for the combination therapy of the autologous HSCT and IFN gene transfer; it may be necessary to establish a method to monitor HP and to maximize the therapeutic effect in individual patients.
In conclusion, a combination of intratumoral IFN-β gene transfer with syngeneic HSCT is a promising new approach for sarcomas, due to the activation of tumor-specific immunity and excellent safety features. This therapeutic strategy deserves an evaluation in future clinical trials for patients with sarcomas.
Footnotes
Acknowledgments
This work was supported in part by a grant-in-aid for the 3rd Term Comprehensive 10-Year Strategy for Cancer Control from the Ministry of Health, Labour and Welfare of Japan; by grants-in-aid for Cancer Research from the Ministry of Health, Labour and Welfare of Japan; by the program for promotion of Foundation Studies in Health Science of the National Institute of Biomedical Innovation (NIBIO); and by the Kobayashi Foundation for Cancer Research. T. Udagawa is an awardee of a Research Resident Fellowship from the Foundation for Promotion of Cancer Research. We thank Vical Inc. for providing the DMRIE-DOPE liposome.
Author Disclosure Statement
No competing financial interests exist.