
Abstract
Filamin A interacting protein 1-like (FILIP1L), which was reported to be consistently absent in ovarian cancer cell lines, has been identified to hold therapeutic potential for inhibiting tumor growth, and its COOH-terminal truncation mutant (FILIP1LΔC103) was found to be more potent than the wild-type. The use of polymeric nanoparticles to deliver functional gene intraperitoneally holds much promise as an effective therapy for ovarian cancer. In this study, a recombinant plasmid expressing FILIP1LΔC103 (FILIP1LΔC103-p) was constructed, and biodegradable cationic heparin-polyethyleneimine (HPEI) nanogels were prepared to deliver FILIP1LΔC103-p into human ovarian cancer SKOV3 cells. The expression of FILIP1LΔC103 in vitro and in vivo was determined using RT-PCR and western blot analysis. Moreover, a SKOV3 intraperitoneal ovarian carcinomatosis model was established to investigate the antitumor activity of HPEI+FILIP1LΔC103-p complexes in nude mice. Tumor weights were evaluated during the treatment course. Cell proliferation and apoptosis were evaluated by Ki-67 immunochemical staining and TUNEL assay, respectively, and the antiangiogenic effect of FILIP1LΔC103-p was assessed by CD31 immunochemical staining and alginate-encapsulated tumor cell assay. FILIP1LΔC103-p could be efficiently transfected into SKOV3 cells by HPEI nanogels. Intraperitoneal administration of HPEI+FILIP1LΔC103-p complexes led to effective growth inhibition of ovarian cancer, in which tumor weight decreased by almost 72% in the treatment group compared with that in the empty-vector control group. Meanwhile, decreased cell proliferation, increased tumor cell apoptosis, and reduction in angiogenesis were observed in the HPEI+FILIP1LΔC103-p group compared with those in the control groups. These results indicated that HPEI nanogels delivering FILIP1LΔC103-p might be of value in the treatment against human ovarian cancer.
Introduction
Ovarian cancer is the most common and lethal malignancy of the female reproductive tract and presently ranks fifth in causing female cancer-related mortality (Jermal et al., 2010). Although much progress has been made in ovarian cancer therapy, the 5-year survival rate of women with this disease has not improved substantially (Raki et al., 2006). Thus, finding novel therapeutic approaches is essential.
Gene therapy, whose potential in cancer treatment has been widely recognized (Anderson et al., 2004; Edelstein et al., 2004; Veiseh et al., 2009), holds great promise for the treatment of a wide variety of diseases. However, except for the target gene, the gene delivery system is the crucial factor affecting the activity of gene therapy. Nonviral vectors hold many advantages over viral carriers, including relative safety, low immunogenicity and toxicity, and ease of large plasmid DNA production (Young and Mautner, 2001; Zhdanov et al., 2002; Gao et al., 2008; Montier et al., 2008; Morille et al., 2008; Suzuki et al., 2010). Polyethyleneimine (PEI), which was used as a gene vector as far back as 1951, becomes progressively one of the most effective nonviral gene carrier agents (Boussif et al., 1995; Lungwitz et al., 2005; Neu et al., 2005). Currently, the commercial PEI25K (25 kg/mol) has been widely considered as a “gold standard” to assess the transfection efficiency of new polymer-based gene carriers. However, PEI, whose transfection efficiency increases along with an increase of cytotoxicity, is not biodegradable. Moreover, PEI has the shortcoming of inducing obvious aggregation of erythrocytes and hemolysis (Kunath et al., 2003; Neu et al., 2005). In our laboratory, PEI2K was chemically conjugated by heparin to form a novel biodegradable cationic nanogel (Fig. 1a), named HPEI. Furthermore, in our previous study, the novel biodegradable cationic HPEI nanogels were used to effectively deliver a plasmid expressing vesicular stomatitis virus matrix protein (pVSVMP) into C-26 cells in vitro and in vivo, and the growth of abdominal and pulmonary metastases in C-26 colon carcinoma in BALB/c mice was efficiently suppressed by pVSVMP/HPEI complexes (Gou et al., 2010). In this work, biodegradable cationic HPEI nanogels were prepared and then used as a gene carrier to deliver a recombinant plasmid encoding FILIP1LΔC103 to treat an SKOV3 intraperitoneal ovarian carcinomatosis in nude mice, with the aim to evaluate the potential effect of FILIP1LΔC103 against ovarian cancer.
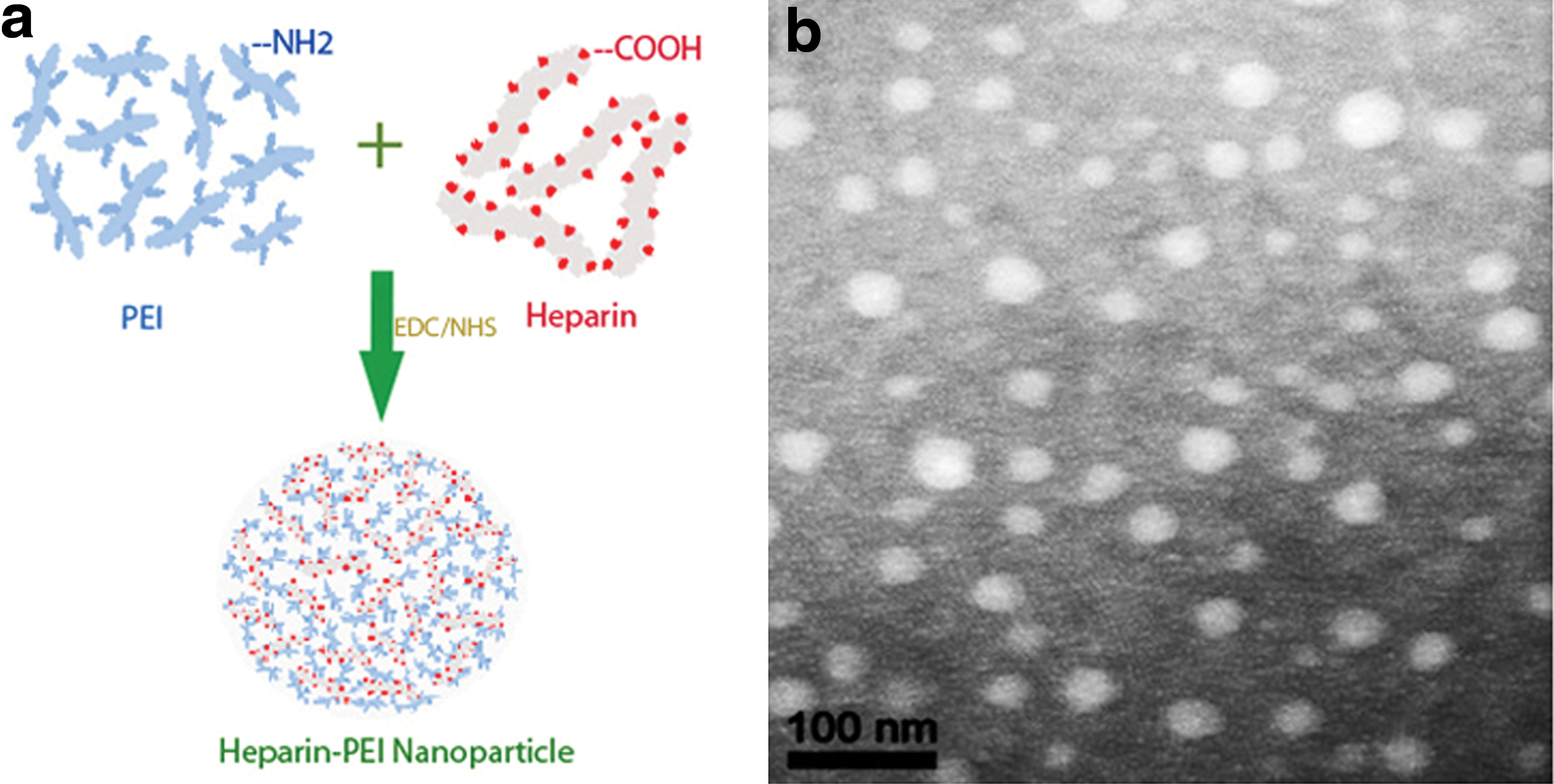
Preparation scheme and characterization of HPEI nanogels. (
Materials and Methods
Cell culture
HUVECs were isolated from human umbilical vein vascular wall as described previously (Baudin et al., 2007). In brief, human umbilical vein vascular wall was digested with collagenase IV at 37°C for 10 min, and the homogenate was centrifuged at 750 g for 10 min. Cells were suspended and seeded on fibronectin-coated plates and cultured in Earle's salts medium supplemented with 10% fetal calf serum (FCS). The human epithelial serous cystadenocarcinoma cell line SKOV3 was obtained from the American Type Culture Collection (ATCC, Rockville, MD) and grown in RPMI 1640 medium (GIBCO, Carlsbad, CA) supplemented with 10% FCS, 2 mM L-glutamine, 100 U/ml penicillin, and 100 μg/ml streptomycin. Cells were maintained in a humidified incubator with 5% CO2 atmosphere at 37°C.
RT-PCR and plasmid construction
The primers of FILIP1LΔC103 were designed based on its cDNA sequence with upstream primer 5′-CGC
pVAX1 plasmid (Invitrogen, San Diego, CA) expressing FILIP1LΔC103, named FILIP1LΔC103-p, was constructed in our laboratory and is illustrated schematically in Fig. 2A. In brief, cultured HUVECs were harvested and total RNA was extracted using TRIzol reagent (Invitrogen) according to the manufacturer's protocol. The RNA sample was then subjected to RT-PCR for amplification of the encoding region of mFILIP1LΔC103, using a One Step RNA PCR Kit (AMV; Takara Bio Inc., Kyoto, Japan) with the above primers and conditions of reaction. The amplified mFILIP1LΔC103 DNA (2,370 bp) and eukaryotic expression vector pVAX1 were digested with BamHI and XhoI, respectively, and then ligated overnight at 16°C using T4 DNA ligase, followed by transformation into competent Escherichia coli DH5α. The resultant recombinant plasmid was double-digested to ensure its correctness, and the identity of FILIP1LΔC103 cloned into the expression plasmid pVAX1 was finally confirmed by automated DNA sequencing. Plasmid without FILIP1LΔC103 was used as an empty-vector control (hereafter called E-p). Colonies of Escherichia coli containing FILIP1LΔC103 or E-p were cultured in Luria-Bertani broth containing 100 μg/ml kanamycin. Large-scale plasmid DNA was purified using an EndoFree Plasmid Giga kit (Qiagen, Valencia, CA). The DNA was dissolved in sterile endotoxin-free water, adjusted eventually to 1.0 mg/ml, and then stored at −20°C for future use.
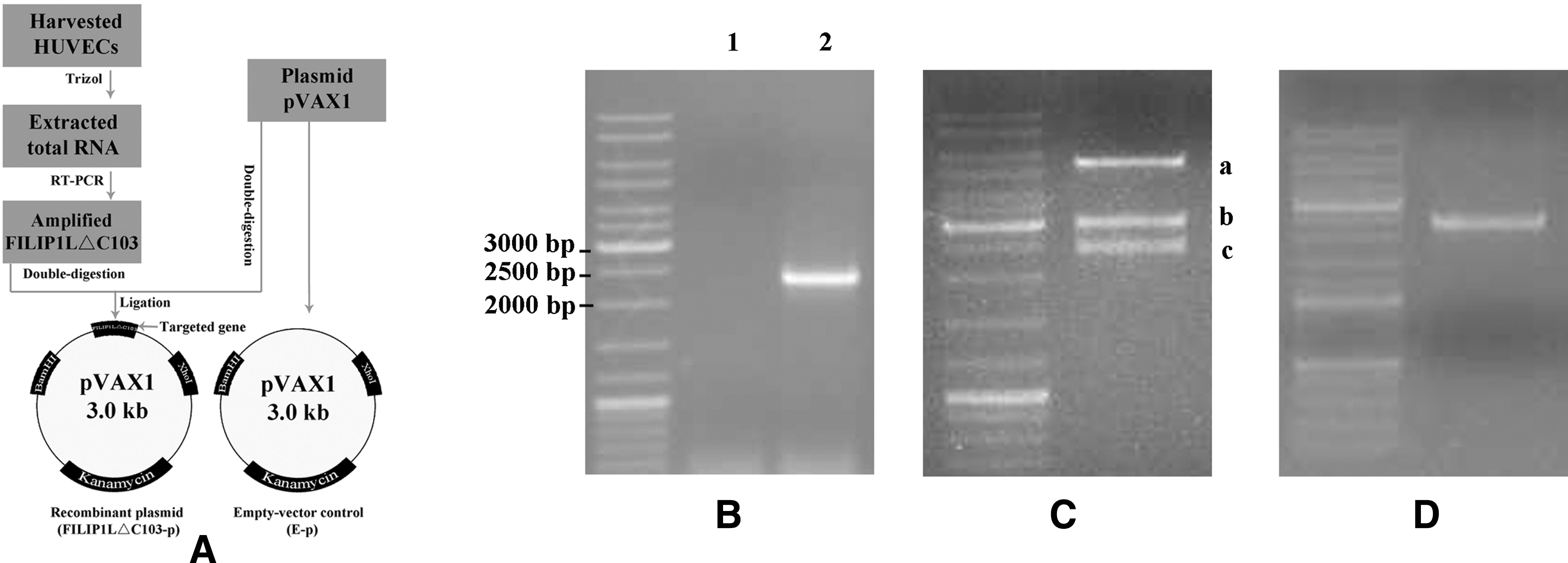
Construction and identification of recombinant plasmid FILIP1LΔC103-p. (
Preparation of HPEI and transfection of plasmid
Biodegradable HPEI nanogels were synthesized as previously described (Gou et al., 2010). In brief, 0.05 g of heparin was first dissolved in 2-(N-morpholino)ethanesulfonic acid (MES) buffering agent (100 ml, 50 mM); 0.02 g of 1-ethyl-3-(3-dimethylaminopropyl)carbodiimide (EDC) and 0.03 g of N-hydroxysuccinimide (NHS) were subsequently added into the above solution agents to activate the carboxylic acid groups of heparin. After 20 ml of PEI2K solution (7.5 mg/ml) was dropped into the above solution, the reaction was carried out at room temperature overnight. Thereafter, the resulting HPEI nanogels were dialyzed in double-distilled water for 3 days. Later, HPEI nanogels were filtered using a syringe filter, then adjusted to a final concentration of 1.0 mg/ml, and stored at 4°C.
SKOV3 ovarian cancer cells (2.0×105) were grown on six-well plates in RPMI 1640 medium and cultured for 24 hr. Twenty micrograms of HPEI and 2 μg of FILIP1LΔC103-p or E-p were diluted in 1 ml of RPMI 1640 medium without antibiotic and serum, respectively, and then combined at a ratio of 10:1. The combinations were transfected to cells with 50–60% confluence. Meanwhile, the medium alone was used as a control agent. After 4 to 6 hr, the medium was replaced by 2 ml of RPMI 1640 medium. After 48 hr of incubation, the cells and supernatants were collected for further assay.
Western blot analysis
Cells or tumor tissues were lysed in RIPA lysis buffer containing 50 mM Tris-HCl (pH 7.4), 0.25% sodium deoxycholate, 150 mM NaCl, 1% NP-40, 1 mM NaF, 1 mM Na3V4, and 1 mM cocktail (Sigma, St. Louis, MO). Protein concentrations were determined using the Bradford assay (Bio-Rad, Hercules, CA). Equal amounts of protein were resolved on a 10% SDS-PAGE, and then electroblotted to polyvinylidene difluoride (PVDF) membranes (Millipore, Bedford, MA), with GAPDH as the loading control. Then the PVDF membranes were blocked with 5% nonfat milk for 2 hr and incubated with rabbit polyclonal antibodies against FILIP1LΔC103 (1: 800 dilution; Santa Cruz Biotechnology, Santa Cruz, CA). Then the membranes were incubated with goat anti-rabbit secondary antibody at 1:5,000 dilution (Abcam, Cambridge, MA) in PBS with Tween 20 for 1 hr. The blot bands were developed using the enhanced chemiluminescence detection system (Pierce Biotech Inc., Rockford, IL) according to the manufacturer's instruction.
Tumor xenograft model and animal treatment
All animal research procedures were approved by the Institutional Animal Care and Use Committee of Sichuan University (Chengdu, People's Republic of China). Female athymic nude mice were used to establish an intraperitoneal carcinomatosis model according to the previous study (Lin et al., 2007). In brief, SKOV3 cells (5×106) in 100 μl of RPMI 1640 medium were injected subcutaneously into the dorsal sides of four mice. When the diameter was about 1 cm, tumors were collected and then minced into tiny particles. Tumor particles were mixed with the RPMI 1640 solution to reach a final volume of 10 ml, and 20 mice were inoculated intraperitoneally with 0.5 ml of the above mixture. Mice were assigned randomly to one of the following groups (five per group): (a) untreated, 100 μl of 5% glucose solution; (b) HPEI, 50 μl of HPEI in 50 μl of 5% glucose solution; (c) HPEI+E-p, 5 μg of pVAX1/50 μl of HPEI in 50 μl of 5% glucose solution; and (d) HPEI+FILIP1LΔC103-p, 5 μg of FILIP1LΔC103-p/50 μl of HPEI in 50 μl of 5% glucose solution. Intraperitoneal administration was initiated 5 days after inoculation. The HPEI/DNA complexes were prepared and incubated at room temperature for 30 min before administration. Mice received therapy every 3 days and were sacrificed after 12 treatments. Intraperitoneal tumors were resected and weighed immediately to assess the antitumoral efficacy.
Alginate-encapsulated tumor cell assay
To explore inhibition of angiogenesis, the alginate-encapsulation assay was done. In brief, SKOV3 cells were resuspended in a 1.5% solution of alginate (Sigma) and added dropwise into a solution of 250 mM CaCl2; an alginate bead was formed containing 1×105 cells. Four beads were then implanted subcutaneously in the back of nude mice. Eight mice were then grouped and treated as described earlier. Treatment was initiated on the same day of implanting beads. After 2 weeks, mice were injected intravenously with 0.2 ml of a 50 mg/kg fluorescein isothiocyanate (FITC)–dextran (Sigma) solution. Alginate beads were photographed after being exposed surgically, and then rapidly removed 20 min after FITC-dextran injection. The uptake of FITC-dextran was measured as described (Hoffmann et al., 1997).
Immunohistochemistry
Deparaffinized tumor sections were immersed in 10 mM citrate buffer (pH 6.0) and then heated in an autoclave for 5 min in saturated steam for antigen retrieval. Endogenous peroxidase activity was quenched in 3% H2O2 for 10 min, and nonspecific binding sites of reagents were subsequently blocked with homeotypic nonimmunoglobulin of secondary antibody at 37°C for 20 min. Then tumor sections were incubated with the primary antibody (R&D Systems, Minneapolis, MN) at 4°C overnight. Tumor sections were incubated with the biotinylated secondary antibody at 37°C for 40 min, followed by sequential incubation with streptavidin–biotin–horseradish peroxidase complex for 40 min at 37°C. Colorimetric detection was performed with diaminobenzidine.
TUNEL assay for apoptotic cells
TUNEL staining was performed to analyze apoptotic cells in tumor tissues using apoptotic cell kits according to the manufacturer's protocol (Promega, Madison, WI). The apoptosis index was calculated by analyzing the average percentage of green fluorescence-positive cells in 10 random fields from different sections at ×400 magnification.
Evaluation of toxicity
The potential treatment-related toxicity and side effects such as weight changes of mice, diarrhea, toxic death, or behavior were observed and evaluated during the whole treatment course. Sections of organs such as heart, liver, spleen, lung, and kidney were stained with hematoxylin and eosin (HE) and observed by different pathologists in a blinded manner.
Statistical analysis
All results are expressed as the means±SD, and one-way ANVOA for multiple-group comparisons was used to analyze differences among four groups. A p value below 0.05 was considered statistically significant.
Results
Cloning FILIP1LΔC103 cDNA and plasmid construction
To obtain FILIP1LΔC103 cDNA fragments, total RNA was extracted from harvested HUVECs, and then RT-PCR was performed. One percent agarose gel electrophoresis revealed one band about 2,370 bp in size as expected. We obtained a fragment of approximately 2,370 bp in length, as shown in Fig. 2B, consistent with the expected size.
The purified PCR product was first double-digested with BamHI/XhoI, and then FILIP1LΔC103 was cloned into the BamHI/XhoI site of the pVAX1 vector treated with the above enzymes. The identity of recombinant plasmid FILIP1LΔC103-p was confirmed by PCR (Fig. 2D), double-digestion (Fig. 2C), and direct automated DNA sequencing.
Preparation and characterization of HPEI
Catalyzed by EDC and NHS, biodegradable cationic HPEI nanogels were formed when PEI2K was chemically conjugated by heparin, as shown in Fig. 1a. According to the image from a transmission electron microscope (TEM; Fig. 1b), cationic HPEI nanogels loading FILIP1LΔC103-p were monodispersed and spherical with a diameter of about 28 nm.
Expression of FILIP1LΔC103 in vitro
First, we used RT-PCR to test the expression of the FILIP1LΔC103 gene in human ovarian cancer SKOV3 cells. Semiquantitative RT-PCR indicated the complete absence of FILIP1LΔC103 transcript expression in SKOV3 ovarian cancer cells, whereas it was expressed in HUVECs, as shown in Fig. 2B.
Next, to investigate whether FILIP1LΔC103 was expressed in human ovarian cancer SKOV3 cells transfected by HPEI+FILIP1LΔC103-p complexes, SKOV3 cells were seeded in six-well plates and then transfected with HPEI+FILIP1LΔC103-p complexes, HPEI+E-p complexes, HPEI nanogels, and 5% glucose (untreated group). After 48 hr of incubation, the in vitro expression of the FILIP1LΔC103 gene in transfected SKOV3 cells was confirmed using RT-PCR and western blot analysis. As shown in Fig. 3A, the expression of FILIP1LΔC103 in SKOV3 cells transfected with HPEI+FILIP1LΔC103-p could be detected, whereas there was no expression of FILIP1LΔΔC103 in SKOV3 cells from the untreated, HPEI, or HPEI+E-p groups.
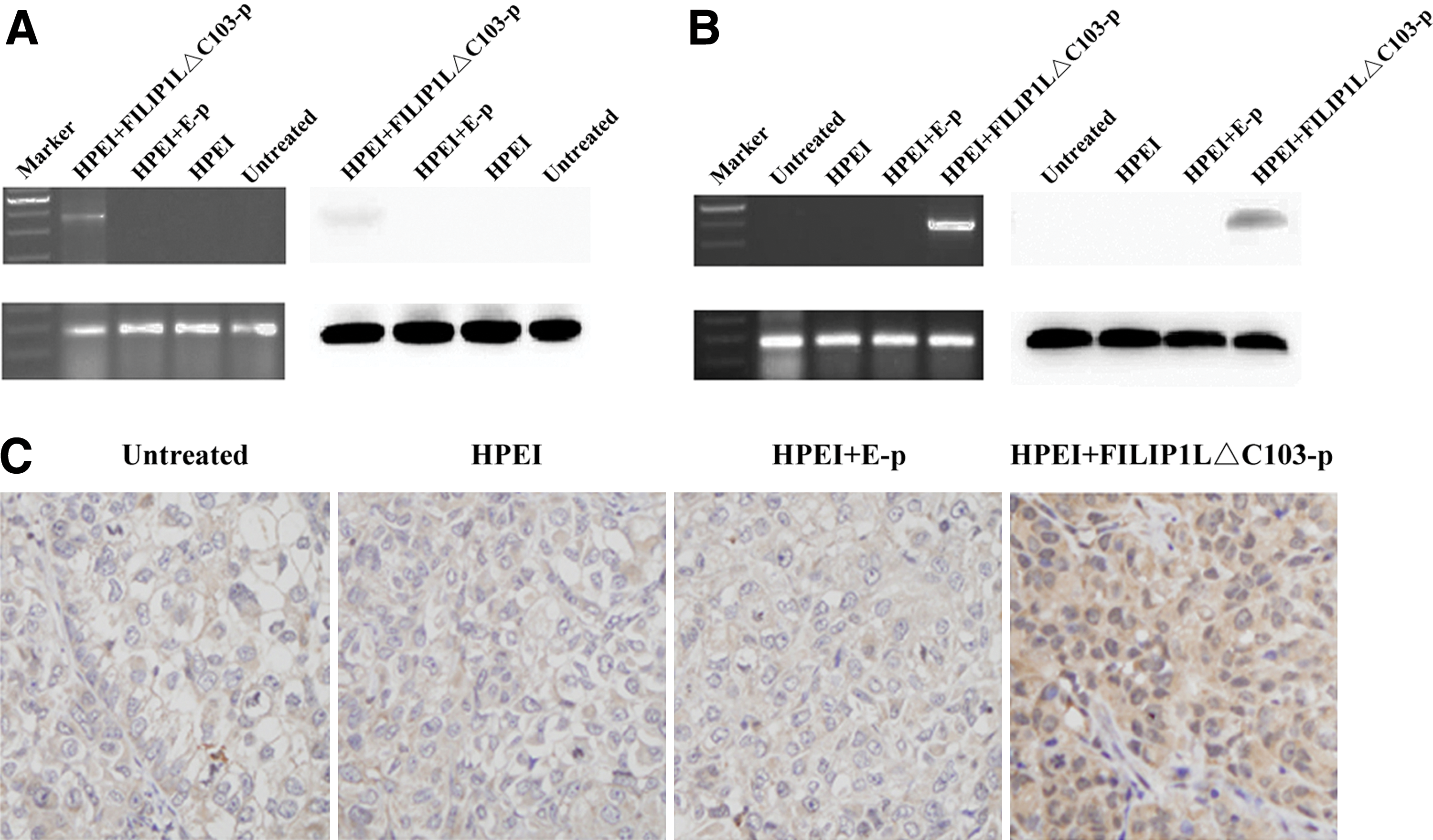
Expression of FILIP1LΔC103-p in vitro and in vivo. (
Expression of FILIP1LΔC103 in vivo
To examine the expression of FILIP1LΔC103 in vivo, an intraperitoneal ovarian carcinoma model was established in nude mice, which were then treated with HPEI+FILIP1LΔC103-p, HPEI+E-p, HPEI, or 5% glucose. Tumors were collected for RT-PCR and western blot on the third day after the final treatment. The level of FILIP1LΔC103 mRNA and expression of FILIP1LΔC103 protein were detected in tumor tissues from the HPEI+FILIP1LΔC103-p group, whereas there was no expression in the HPEI, E-p, and 5% glucose groups (Fig. 3B). In addition, immunochemical staining of FILIP1LΔC103 in tumor tissues also suggested expression of FILIP1LΔC103 in the HPEI+FILIP1LΔC103-p complex–treated group as shown in Fig. 3C.
HPEI+FILIP1LΔC103-p complexes inhibited intraperitoneal ovarian cancer xenograft growth in nude mice
To evaluate the effect of HPEI+FILIP1LΔC103-p on suppressing ovarian cancer growth in vivo, we established an intraperitoneal xenograft model of human ovarian cancer. Nude mice were grouped and treated as aforementioned, then sacrificed at the termination of the animal experiment. Intraperitoneal tumor nodules were collected totally and carefully; the average weights were 1.23±0.18 g, 1.25±0.13 g, 1.04±0.14 g, and 0.29±0.11 g in the untreated, HPEI, HPEI+E-p, and HPEI+FILIP1LΔC103-p groups, respectively (p<0.01; Fig. 4B and C). HPEI+FILIP1LΔC103-p reduced tumor weight by almost 72% compared with the HPEI+E-p group. There was no significant difference in tumor weight among the three control groups (p>0.05).
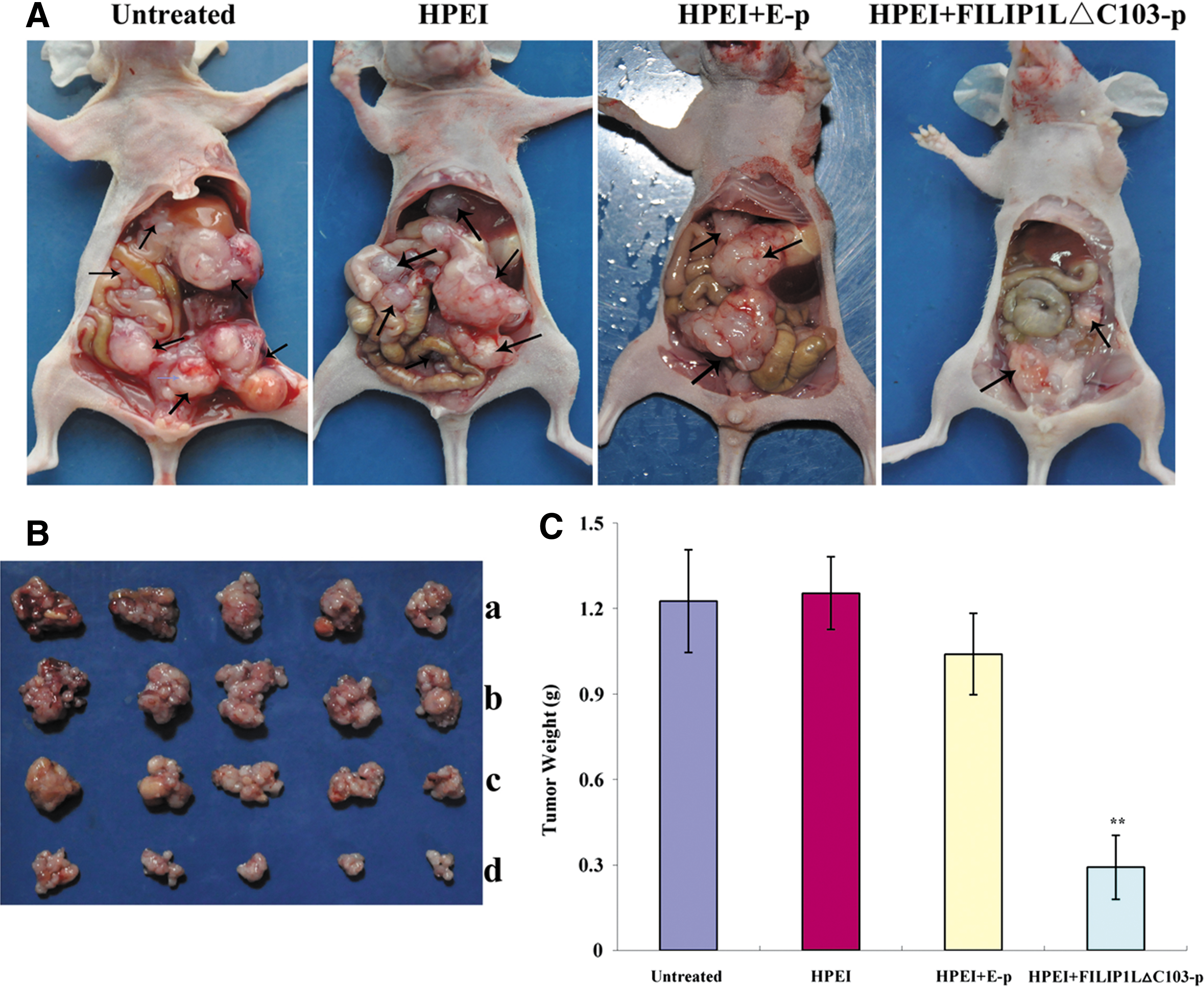
FILIP1LΔC103-p inhibited intraperitoneal ovarian cancer xenograft growth in nude mice. (
Intraperitoneal tumor nodules in a wide range of sizes were scattered predominantly in the pelvis, as well as below the liver, in the three control groups (Fig. 4A). Ascites were found in three mice in the untreated group, in four mice in the HPEI-treated group, and in two mice in the HPEI+E-p–treated group, including hemorrhagic ascites in one, two, and one from the untreated, HPEI, and HPEI+E-p groups, respectively (Table 1). None of the mice in the HPEI+FILIP1LΔC103-p group developed ascites, and intraperitoneal tumor nodules of mice in this group were all scattered in the pelvis.
Volume of ascites or hemorrhagic ascites found in nude mice was all less than 3 ml.
Suppression of angiogenesis by HPEI+FILIP1LΔC103-p in vivo
To explore potential mechanisms underlying the antitumor effect of HPEI+FILIP1LΔC103-p in vivo, and as antiangiogenesis is generally considered to be an important mechanism for cancer therapy, frozen sections were stained by antibody of CD31 to investigate the antiangiogenic effect of HPEI+FILIP1LΔC103-p. Angiogenesis in tumor tissues was evaluated by microvessel density (MVD) in different sections stained with an antibody reactive to CD31, which had high specific affinity for vascular endothelial cells. CD31 immunochemical staining revealed a significant reduction of MVD in tumor tissues of the HPEI+FILIP1LΔC103-p group in comparison with tissues of the untreated, HPEI, and HPEI+E-p groups (5.10±2.73 vs. 26.70±6.13 vs. 27.00±6.75 vs. 24.70±5.50, respectively; p<0.01; Fig. 5A); there was also no significant difference among the numbers of microvessels of the different control groups (p>0.05).
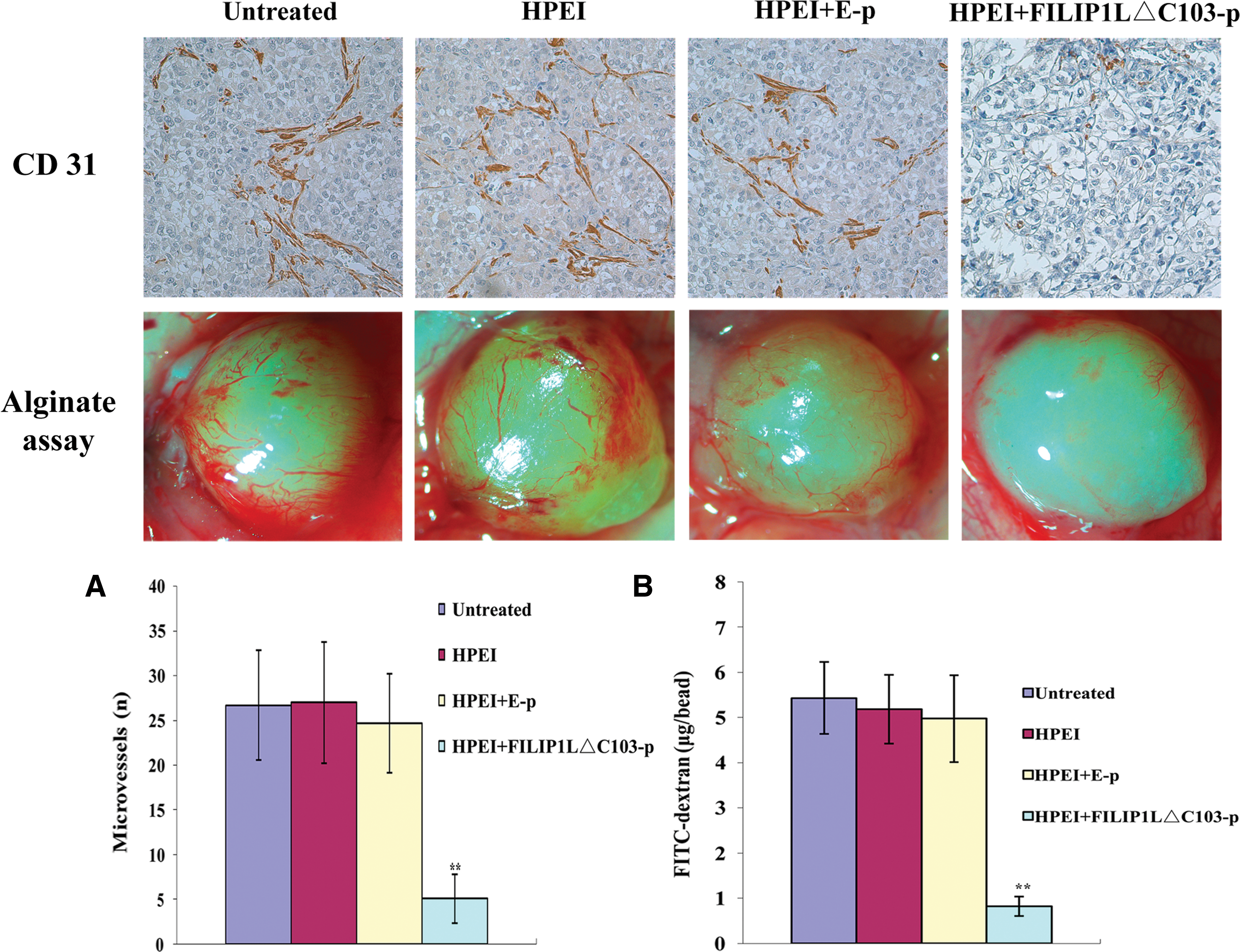
Inhibition of angiogenesis in vivo. (
In addition, the capability of antiangiogenesis was also detected using the alginate-encapsulated tumor cell assay; newborn blood vessels in alginate beads from nude mice treated with HPEI+FILIP1LΔC103-p were apparently fewer than those in the other control groups. Moreover, the levels of FITC-dextran uptake were significantly low in mice treated with HPEI+FILIP1LΔC103-p compared with levels in the three control groups (0.83±0.21 μg/bead vs. 5.43±0.79 μg/bead vs. 5.19±0.76 μg/bead vs. 4.98±0.96 μg/bead, respectively; p<0.01; Fig. 5B).
The above results suggested that angiogenesis of human ovarian cancer xenograft was effectively inhibited in nude mice treated with HPEI+FILIP1LΔC103-p. Therefore, we might conclude that the antitumor activity of HPEI+FILIP1LΔC103-p is due, at least partially, to the inhibition of tumor neovascularization in ovarian cancer.
Inhibition of cell proliferation by HPEI+FILIP1LΔC103-p in vivo
To explore further the role of HPEI+FILIP1LΔC103-p therapy in ovarian cancer in vivo, we performed Ki-67 immunohistochemistry to investigate whether the antitumor effect of HPEI+FILIP1LΔC103-p correlated with the decreased cell proliferation. The proliferation index was determined by a ratio of the Ki-67-positive cells to the total number of cancer cells. The most significant reduction of Ki-67 expression was observed in the tumors of mice receiving HPEI+FILIP1LΔC103-p treatment when compared with the three controls groups (18.50±6.67 vs. 46.40±14.92 vs. 45.10±15.89 vs. 37.40±13.77, respectively; p<0.05; Fig. 6A).
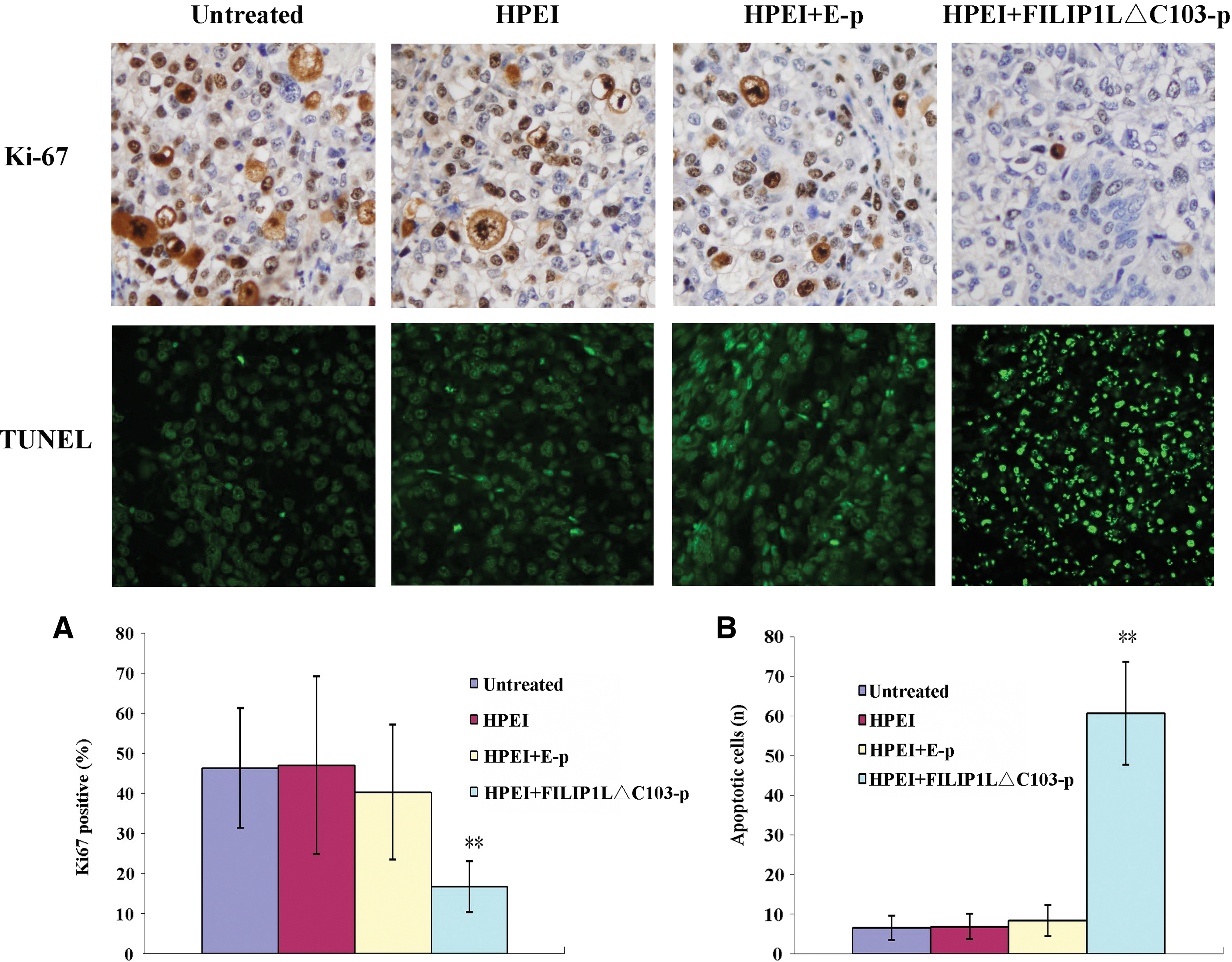
Effects of expression of FILIP1LΔC103-p on cell apoptosis and proliferation in vivo. (
Induction of cell apoptosis by HPEI+FILIP1LΔC103-p in vivo
As tumor growth may be considered an imbalance between cell proliferation and apoptosis, we also applied the TUNEL assay to explore possible mechanisms underlying the antitumor effect of HPEI+FILIP1LΔC103-p in vivo. The TUNEL assay showed many apoptotic cells (TUNEL-positive nuclei) in the tumor sections of the HPEI+FILIP1LΔC103-p complexes group. However, there were few such cells in the three control groups. The apoptotic index revealed that tumors in the HPEI+FILIP1LΔC103-p group had significantly more TUNEL-positive nuclei than tumors in the 5% glucose, HPEI, or HPEI+E-p groups (55.80±9.40 vs. 6.60±3.10 vs. 6.90±3.18 vs. 10.70±3.92, respectively; p<0.01; Fig. 6B).
Observation of toxicity
Because animal weight was considered a parameter for evaluation of physical status, we measured the weight of mice every 3 days (Fig. 7A) and observed no significant differences among the four groups. Moreover, HE histological staining of spleen, liver, lung, kidney, and heart did not demonstrate any significant pathologic differences (Fig. 7B).
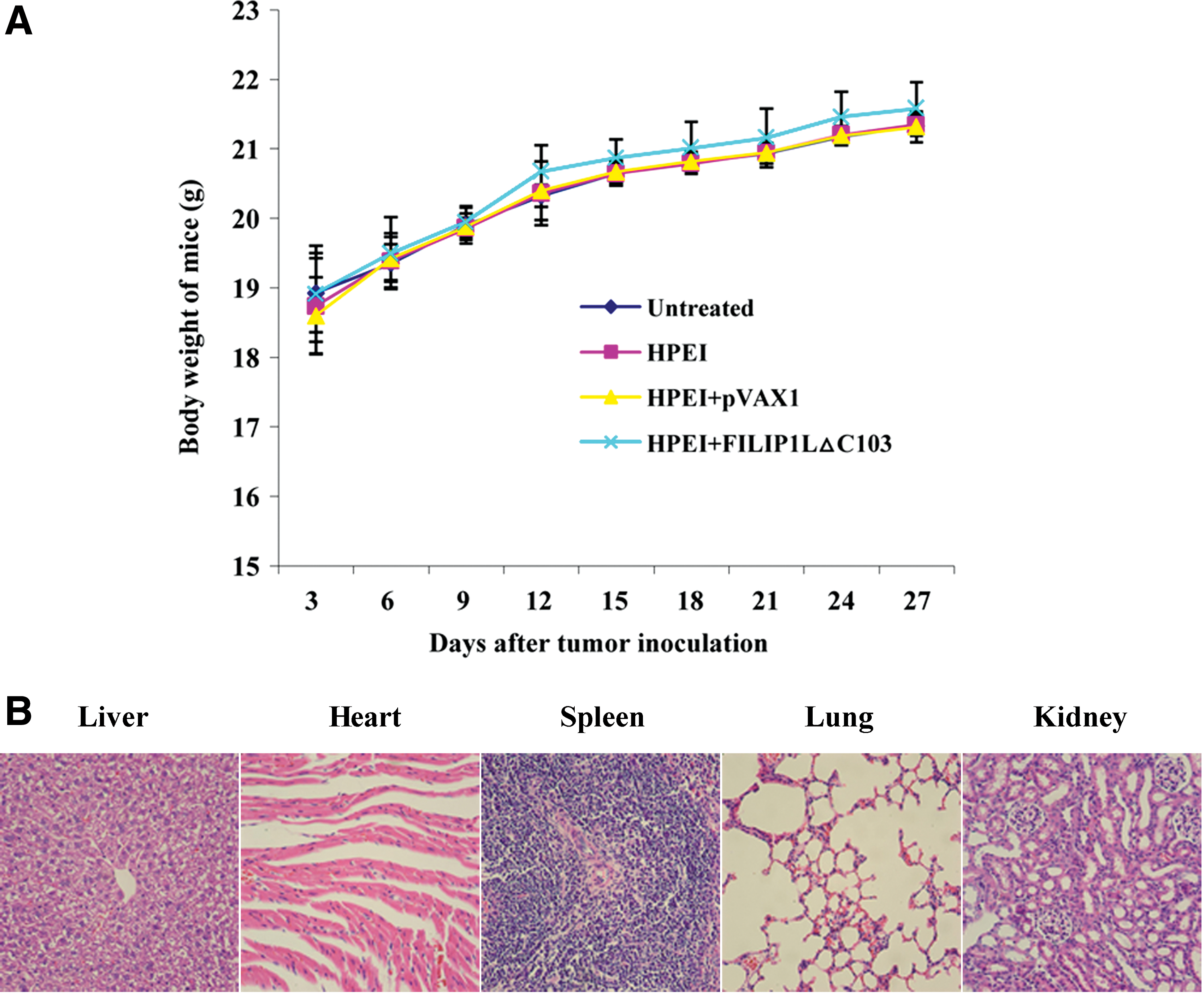
Observation of toxicity in mice treated with HPEI+FILIP1LΔC103. (
Discussion
Previous study has shown that growth of xenografted human malignant melanoma can be suppressed by introducing the different mutant-type genes of FILIP1L, and the COOH-terminal truncation mutant 1-790 (FILIP1LΔC103) demonstrated a most potent activity (Kwon et al., 2008). However, studies on the antitumoral efficacy of the FILIP1LΔC103 gene in human ovarian cancer have been lacking until now. In the present study, we constructed a recombinant plasmid expressing FILIP1LΔC103 (FILIP1LΔC103-p), which was then used to effectively inhibit intraperitoneal xenograft growth of human ovarian cancer in nude mice, with biodegradable cationic HPEI nanogels as a novel delivery system. Meanwhile, treatment with HPEI+FILIP1LΔC103-p complexes in nude mice was shown to be without detectable toxicity.
The FILIP1L gene, whose translation product has 893 amino acids, is located on the region of chromosome 3 in humans. It was identified as a potentially important regulator of angiogenesis activity. Overexpression of FILIP1L could lead to inhibition of proliferation and induction of apoptosis, whereas FILIP1LΔC103 is more potent than wild-type FILIP1L in mediating these activities (Kwon et al., 2008). Results obtained using the microarray technique revealed that FILIP1L played a possible role in the pattern of suppressing tumor (Ross et al., 2000; Su et al., 2001). A sustained increment of FILIP1L gene expression over the time course was observed in HUVECs in response to 5-fluorouracil treatment, suggesting that the gene might be related to cell death (Tandle et al., 2005). Furthermore, FILIP1L has a novel function in cell senescence, which has an important role in inhibiting the occurrence of an immortal phenotype, a crucial feature in tumorigenesis (Schwarze et al., 2002). FILIP1L was reported to be present in human normal ovarian epithelial cells, but consistently absent in ovarian cancer cell lines (Mok et al., 1994). Consequently, we may wonder whether transferring the FILIP1L gene into ovarian cancer cells by a suitable delivery system could be an approach for treating ovarian cancer.
Over the last decade, many functional genes have been reported to effectively inhibit ovarian cancer in vitro and in vivo (Malek et al., 2008; Nguyen et al., 2010), but the absence of efficient and safe gene delivery technologies is a major obstacle to the clinical use of these gene treatments. Currently, many delivery systems have been applied in gene transfer; these are generally classified as nonviral and viral vectors. Previous studies have demonstrated that nonviral vectors have many advantages over viral vectors (Young and Mautner, 2001; Zhdanov et al., 2002; Gao et al., 2008; Montier et al., 2008; Morille et al., 2008; Suzuki et al., 2010). In this work, we developed a novel nonviral gene delivery system based on PEI. It has the advantages of PEI and resolves the problem of unbiodegradability and the transfection efficiency–dependent cytotoxicity of PEI. In our study, heparin was used to conjugate PEI into biodegradable cationic nanogels for the following reasons. First, it is relatively stable in vitro and easy to degrade through enzymolysis and hydrolysis in vivo. Second, as a natural polysaccharide, it is nontoxic and biocompatible. Third, it could improve the biocompatibility of the nanogels. In our previous study, the transfection efficiency of HPEI was found to be comparable to, but less toxic than, that of PEI25k. Analyses of the erythrocyte aggregation and hemolysis showed that HPEI had a better blood compatibility than PEI25K. Moreover, HPEI nanogels were stable in vitro and could be quickly degraded into the low-molecular-weight chemical compound PEI excreted in the urine (Gou et al., 2010). Based on these advantages, HPEI nanogels were used to deliver a recombinant plasmid to evaluate the antitumor activity of FILIP1LΔC103, as well as the efficacy and safety of this delivery system.
In the current study, we have demonstrated that a recombinant plasmid encoding FILIP1LΔC103 can significantly suppress intraperitoneal xenograft growth of human ovarian cancer in nude mice. This provides proof, in principle, that FILIP1LΔC103 possesses antitumor activity against human ovarian cancer in vivo. To elucidate the antitumor mechanism in vivo of FILIP1LΔC103, inhibition of angiogenesis including the alginate assay and CD31 immunohistochemistry was performed. The results indicated that the MVD of tumor tissue sections was significantly reduced by FILIP1LΔC103-p and that angiogenesis was significantly inhibited in the FILIP1LΔC103-p group compared with that in the other controls. Data from proliferation and apoptosis analysis using Ki-67 staining and the TUNEL technique showed that expression of FILIP1LΔC103 resulted in significant inhibition of cell proliferation and induction of cell apoptosis compared with the control therapies. These results demonstrated that the antitumor activity of FILIP1LΔC103-p in xenografted ovarian cancer might be ascribed to successfully sustained expression of FILIP1LΔC103 in vivo. However, the exact mechanism of its antitumor activity requires further investigation. Previous study had shown that when the gene expression of FILIP1L was silenced, endothelial monocyte-activating polypeptide II (EMAP II) failed to show a modulatory effect on the expression of KLF4, ADM, SOCS3, and TNFAIP3 (Tandle et al., 2005), suggesting that FILIP1L may play a role in mediating the effect of EMAP II. Similarly, the up-regulation of gene expression of TC-1 and KLF4 in response to endostatin was observed with silencing of the gene encoding FILIP1L at the mRNA level using small interfering RNA; it is suggested that FILIP1L might be upstream of TC-1 and KLF4 in a pathway mediating the antiangiogenic response to endostatin treatment (Mazzanti et al., 2004). These data suggest that FILIP1L might play a primary role in mediating some common response pathways observed in endothelial cells after exposure to angiogenic inhibitors.
In this study, using HPEI to deliver FILIP1LΔC103-p to treat nude mice bearing ovarian cancer, we found that intraperitoneal xenografted ovarian cancer could be effectively suppressed; also, evaluation of toxicity indicated that treatment with the HPEI+FILIP1LΔC103-p complex seemed to be safe and without detectable toxicity at the dose we used. Nevertheless, the toxicity of this delivery system in other research requires further investigation, and a study on the antitumor activity of FILIP1LΔC103 in other cancers may be required. Future investigations into these points might be interesting.
In conclusion, our results suggested that FILIP1LΔC103-p could be effectively transfected into human ovarian cancer SKOV3 cells by HPEI nanogels, and biodegradable cationic HPEI nanogels might be a safe and effective delivery system. FILIP1LΔC103, as a potential angiogenesis inhibitor, could efficiently inhibit the growth of xenografted ovarian cancer through the inhibition of angiogenesis and tumor cell proliferation and the induction of tumor-cell apoptosis, and HPEI nanogel-delivered FILIP1LΔC103-p may have a potentially promising application against ovarian cancer.
Footnotes
Acknowledgments
This work was supported by the National Key Basic Research Program (973 Program) of China (2011CB910703).
Author Disclosure Statement
The authors have declared no conflicts of interest.