
Abstract
Age-related macular degeneration (AMD) is a complex disease that has two phases: a degenerative phase often referred to as nonneovascular AMD (non-NVAMD) or dry AMD and a phase dominated by growth of new blood vessels in the subretinal space, referred to as NVAMD or wet AMD. Advances in the understanding of the molecular pathogenesis of NVAMD have led to new drug therapies that have provided major benefits to patients. However, those treatments require frequent intraocular injections that in many patients must be continued indefinitely to maintain visual benefits. Gene transfer to augment expression of endogenous antiangiogenic proteins is an alternative approach that has the potential to provide long-term stability in patients with NVAMD. Studies in animal models that mimic aspects of NVAMD have identified several possible transgenes, and a clinical trial in patients with advanced NVAMD has suggested that the approach may be feasible. Many important questions remain, but the rationale and preliminary data are compelling. The results of two ongoing clinical trials may answer several of the questions and help direct future research.
Introduction
Relevant anatomy of the eye
The photoreceptors occupy the outer one-third of the retina, with cell bodies in the outer nuclear layer (ONL) connecting to inner segments, packed with mitochondria, that connect to outer segments containing chromophores, which capture light. The outer segments interdigitate with the apical surface of retinal pigmented epithelial (RPE) cells, and under normal circumstances the subretinal space is only a potential space between the outer segments and the RPE (Fig. 1, cross-section). The RPE sits on Bruch's membrane, which separates the RPE from the highly vascular choroid. The outer retina is avascular and receives all of its oxygen by diffusion from the choroid. The choroidal vasculature is an extremely dense, high-flow system that ends in a fenestrated capillary bed, the choriocapillaris, that is separated by Bruch's membrane from the RPE and photoreceptors. The fenestrations of the choriocapillaris allow plasma to pool around the RPE, which has tight junctions that prevent fluid from entering the retina. The RPE allows passage of oxygen and facilitates transport of nutrients to the photoreceptors. The apical tight junctions and other barrier characteristics of the RPE constitute the outer blood–retinal barrier.
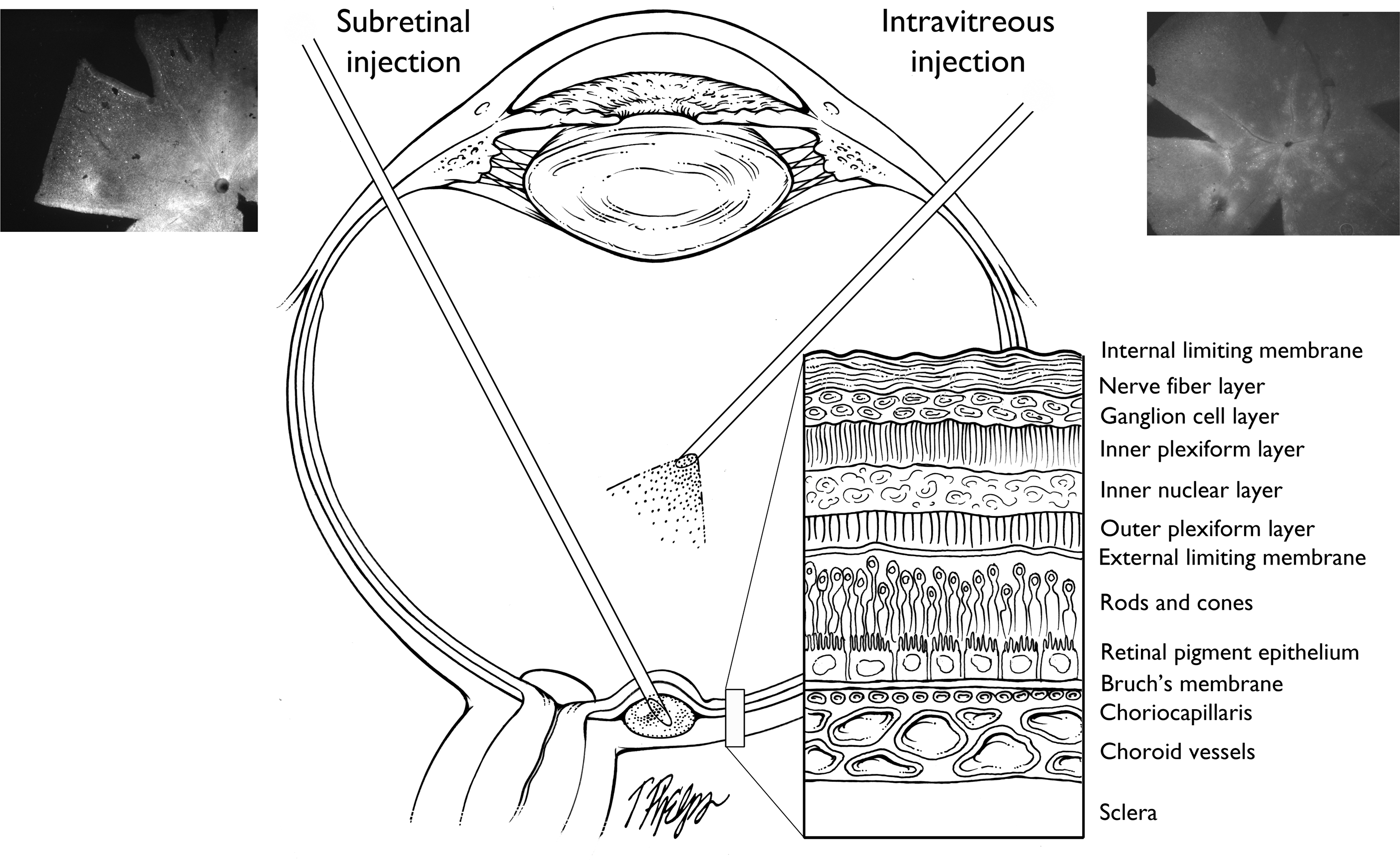
Schematic showing subretinal and intravitreous injection routes and structure of the retina. Left: Subretinal injection. The needle enters the eye through the pars plana of the ciliary body, passes through the vitreous and retina, and vector is injected into the subretinal space. A flat mount of a mouse retina 6 weeks after subretinal injection of AAV2.GFP is shown; there is diffuse fluorescence, indicating strong expression of GFP around the optic nerve and throughout the posterior retina that was detached by injection of the vector. Right: Intravitreous injection. The needle enters through the pars plana and vector is injected into the vitreous cavity in front of the retina. The flat mount shows a mouse retina 6 weeks after subretinal injection of AAV2.GFP. There are small focal areas of fluorescence that retinal sections show to be due primarily to transduction of ganglion cells. The cross-section shows the cell layers from the sclera to the vitreous cavity.
Pathogenesis of AMD
AMD is a complex disease in which genetic susceptibilities combined with environmental exposures result in a disease phenotype that consists of extracellular deposits (drusen) that occur along a diffusely thickened Bruch's membrane and gradual death of photoreceptors and RPE cells. This phase of the disease is referred to as atrophic or nonneovascular AMD (non-NVAMD) or by the lay community as dry AMD. Over time the majority of patients with AMD experience gradual deterioration of central vision from progressive death of photoreceptors and RPE cells, resulting in patches of atrophy referred to as geographic atrophy. Genetic studies have suggested that complement-mediated damage plays a role in susceptibility to AMD (Edwards et al., 2005; Haines et al., 2005; Klein et al., 2005; Gold et al., 2006), but exactly how it plays a role is unknown. In general, the pathogenesis of non-NVAMD is not sufficiently understood to determine a clear path to development of a therapy at this time.
About 20% of patients with non-NVAMD convert to NVAMD (also called wet AMD) by developing new blood vessels that originate from the choroid and/or the deep capillary bed of the retina and invade the subretinal space (subretinal NV). The exact mechanism by which this conversion occurs is unknown, but vascular endothelia growth factor (VEGF) plays a critical role because blockade of VEGF is sufficient to strongly suppress the development of subretinal NV (Kwak et al., 2000). Rupture of Bruch's membrane by laser photocoagulation in mice is accompanied by transient upregulation of VEGF in the surrounding retina and RPE and results in subretinal NV (Tobe et al., 1998); it has become a widely accepted model, because results in the model (Kwak et al., 2000; Saishin et al., 2003) have been predictive of results in patients with NVAMD (Brown et al., 2006; Rosenfeld et al., 2006). Intraocular injections of VEGF antagonists provide tremendous benefit in patients with NVAMD and have become standard of care.
Vessels within subretinal NV lack tight junctions and leak fluid into the subretinal space and retina, causing edema and fluid-filled cavities. This excess fluid causes reversible loss of vision, and the major effect of intraocular injections of VEGF antagonists is to reduce leakage, allowing reduction (often elimination) of excess fluid and improvement in vision. In most patients, excess production of VEGF is sustained and injections as frequently as every month are necessary to prevent reaccumulation of fluid. If excess fluid is persistent for many months, there may be damage to the photoreceptors, resulting in permanent loss of vision. Another source of permanent vision loss is subretinal fibrosis, which occurs because the new vessels recruit other cells; this recruitment and hence subretinal fibrosis is prevented by aggressive treatment with anti-VEGF agents. A third possible cause of permanent vision loss is subretinal hemorrhage, which if sufficiently thick, reduces delivery of oxygen and nutrients from the choroid to the overlying retina, resulting in massive cell death. In general, repeated intraocular injections of a VEGF antagonist are needed and clinical trial outcomes are superior when injections are given monthly as opposed to less frequently. Thus, constant blockade of VEGF provides the best chance of maximizing vision and preventing gradual deterioration from chronic edema or fibrosis, or a catastrophic event such as a large subretinal hemorrhage.
Gene transfer to express antiangiogenic proteins
The actions of proangiogenic factors, such as VEGF, are opposed by a variety of antiangiogenic proteins. Gene transfer to augment expression of these endogenous inhibitors or related engineered proteins is a potential alternative to suppress subretinal NV and avoid frequent intraocular injections. There is considerable preclinical and emerging clinical data suggesting that this approach may be feasible. There are two major routes of administration of viral vectors: intravitreous injection and subretinal injection (Fig. 1). Intravitreous injection of adenoviral (Ad) vectors results in little transduction of retinal cells, but strong transduction of cells in the iris and cornea, which can result in substantial levels of a secreted protein in the retina (Mori et al., 2001b). Subretinal injection of Ad vectors causes strong transduction of RPE cells. With Ad vectors, the duration of expression is transient (about 1 month in mice, but possibly longer in humans), but adeno-associated viral (AAV) vectors and lentiviral vectors provide long-term expression. Intravitreous injection of AAV2.GFP causes small focal areas of fluorescence in the retina, primarily from infection of a small number of ganglion cells (Fig. 1, right), whereas subretinal injection of AAV2.GFP causes diffuse, strong fluorescence throughout the area where the retina was elevated by the injection, due to infection of RPE and photoreceptor cells (Fig. 1, left). With injection of the same AAV2 vector dose, the subretinal route results in much higher expression levels than the intravitreous route. This difference is even greater for lentiviral vectors, for which there is little or no expression after intravitreous injections but high-level expression after subretinal injections, due to strong infection of RPE cells (Takahashi et al., 2002).
Pigment epithelium-derived factor
Pigment epithelium-derived factor (PEDF) is a serine proteinase inhibitor that was isolated from cultured retinal pigmented epithelial (RPE) cells, based on its ability to cause neurite outgrowth from cultured retinoblastoma cells (Tombran-Tink et al., 1991). PEDF promotes survival of cultured neurons and protects photoreceptors from excessive light exposure (Steele et al., 1993; Araki et al., 1998; Bilak et al., 1999; Cao et al., 2001) and also has antiangiogenic activity (Dawson et al., 1999). The combination of neurotrophic and antiangiogenic activities makes increasing expression of PEDF appealing as a potential therapy for AMD. Intravitreous or subretinal injection of an E1-, partial E3-, E4-deleted adenoviral vector expressing human PEDF (AdPEDF.11) suppressed the development of retinal or subretinal NV (Mori et al., 2001b) and caused regression when given after NV was established (Mori et al., 2001c). Intraocular AAV2-mediated gene transfer of Pedf also suppressed retinal and subretinal NV (Auricchio et al., 2002; Mori et al., 2002; Raisler et al., 2002).
The encouraging preclinical results with AdPEDF led to a phase I clinical trial in patients with advanced NVAMD (Campochiaro et al., 2006). This was an open label study in which 28 patients were given a single intravitreous injection of AdPEDF.11 with doses ranging from 106 to 109.5 particle units (PU). Signs of mild, transient intraocular inflammation occurred in 25% of patients, but there was no severe inflammation, no serious adverse events, and no dose-limiting toxicities. Six patients experienced increased intraocular pressure that was easily controlled by topical medication. All adenoviral cultures were negative. There was no control group, but by comparison of effects in patients who received low doses, 106–107.5 PU, versus those who received higher doses, 108–109.5 PU, it is possible to derive some assessment of biological activity. Masked grading of lesion size was done in an independent reading center. The percentage of patients who had no change or improvement in lesion size at 3 months was 94% in the high-dose group compared with 55% in the low-dose group, and at 6 months it was 71% in the high-dose group versus 50% in the low-dose group. The median increase in lesion size at 6 and 12 months was 0.5 and 1.0 disc areas in the low-dose group compared with 0 disc areas at both time points in the high-dose group. These data suggest a single intravitreous injection of a dose greater than 108 PU of AdPEDF.11 resulted in antiangiogenic activity that lasted for several months. This study provided proof-of-concept for the strategy of gene transfer of antiangiogenic proteins to treat NVAMD and has helped to stimulate additional investigations using vectors that express for longer durations.
VEGF-binding proteins
The secreted extracellular domain of VEGF receptor-1, sFlt-1, is a naturally occurring protein antagonist of VEGF formed by alternative splicing of the pre-mRNA for the full-length receptor (Kendall et al., 1996; He et al., 1999). Intraocular injection of Ad.sFlt-1 strongly suppressed retinal or subretinal NV in mice (Honda et al., 2000; Rota et al., 2004), and injection of AAV2.sFlt-1 suppressed subretinal NV in mice and monkeys (Lai et al., 2002, 2005). Genzyme (Framingham, MA) has developed a VEGF-binding protein that consists of domain 2 of Flt-1 linked to a human immunoglobulin G1 (IgG) heavy chain Fc fragment (sFLT01). Intravitreous injection of AAV2.sFLT01 suppressed retinal NV in mice and subretinal NV in primates (Pechan et al., 2009; Lukason et al., 2011).
These results combined with results based on the intraocular injection of ranibizumab or bevacizumab in patients, leave little doubt that ocular gene transfer of a VEGF-binding protein will provide efficacy if expression is sufficient, but concern has been raised about the possibility of toxicity. Systemic injections of sFlt-1 three times per week for 8 weeks, and intraocular injections of an anti-VEGF antibody for 6 weeks, caused loss of 50% of retinal ganglion cells in mice (Nishijima et al., 2007). Systemic injection of 2.5 × 109 Ad-CMV-sFlt-1 caused massive cell death in the inner nuclear layer (INL) and outer nuclear layer (ONL) of the retina (Saint-Geniez et al., 2008). Mice that produce VEGF188, but not the soluble isoforms of VEGF, showed choriocapillaris atrophy, loss of RPE, increased photoreceptor apoptosis, and reduced electroretinogram (ERG) b-wave amplitudes starting at 7 months of age, suggesting that soluble forms of VEGF derived from the RPE may be necessary for maintenance of the choriocapillaris (Saint-Geniez et al., 2009).
In contrast, when given doxycycline the retinas of double-transgenic mice with doxycycline-inducible expression of sFlt-1Fc expressed high levels of sFlt-1Fc that completely blocked VEGF-induced retinal vascular permeability and caused strong suppression of subretinal NV at Bruch's membrane rupture sites, but did not show any evidence of retinal toxicity from VEGF blockade (Ueno et al., 2008). Mice with constant expression of sFlt-1Fc in the retina for 7 months had normal ERGs and normal retinal and choroidal ultrastructure including normal fenestrations in the choriocapillaris. They also showed no significant difference from control mice in the number of ganglion cell axons in optic nerve cross-sections or the retinal level of mRNA for three ganglion cell-specific genes. Likewise, complete blockade of VEGF receptor signaling for up to 12 weeks with SU4312, a selective VEGF receptor kinase inhibitor, failed to cause any retinal toxicity (Miki et al., 2010). Cynomolgus monkeys given an intravitreous injection of 2.4 × 109 or 2.4 × 1010 vector genomes (VG) of AAV2.sFLT01 showed no ERG or fluorescein angiographic abnormalities for up to 12 months, the longest time point evaluated (Maclachlan et al., 2010). Mild to moderate vitreous inflammation that was transient and resolved spontaneously was seen in the high-dose, but not the low-dose, group. Histopathological examinations of eyes from the low-dose group were normal and those from the high-dose group showed only occasional inflammatory cells in the trabecular meshwork, vitreous, and/or retina. In the low-dose group (n = 6), aqueous levels of sFLT01 ranged from about 10 to 100 ng/ml at 1 month and decreased slightly but were still in the same general range at 12 months. Vitreous levels of sFLT01 at 12 months were 200–300 ng/ml in two monkeys and undetectable in a third. In the high-dose group (n = 6), aqueous levels of sFLT01 ranged from 100 to 500 ng/ml at 1 month, decreased to 20–100 ng/ml at 3 months, and remained in that range through 12 months, while vitreous levels were 400–600 ng/ml in two monkeys at 12 months and about 100 ng/ml in another monkey measured at 18 months.
These encouraging preclinical data have led to a phase I dose-ranging trial testing four doses, 2 × 108, 2 × 109, 6 × 109, and 2 × 1010 VG, in patients with advanced NVAMD (registration no. NCT01024998;
Endostatin and angiostatin
Endostatin is a cleavage product of collagen XVIII and angiostatin is a cleavage product of fibrinogen, and each inhibits tumor angiogenesis (O'Reilly et al., 1994, 1997). Endostatin participates in physiological regression of the hyaloid vasculature and regulation of retinal vascular development, because mice deficient in collagen XVIII, the source of endostatin, show delayed regression of the hyaloid vasculature and poor development of the retinal vasculature (Fukai et al., 2002). After tail vein injection of Ad.CMV-endostatin or Ad.RSV-endostatin, serum levels of endostatin were 10-fold higher in mice injected with Ad.CMV-endostatin (Mori et al., 2001a). There was a strong positive correlation between endostatin serum level and inhibition of choroidal NV. Subretinal injection of a bovine immunodeficiency lentiviral (BIV) vector encoding endostatin (BIVendostatin) resulted in high expression of endostatin in the RPE that diffused into the overlying retina and outlined retinal vessels, suggesting endostatin binding to a component of vessel walls (Takahashi et al., 2003). In double-transgenic mice with doxycycline-induced expression of VEGF in the retina (Ohno-Matsui et al., 2002), gene transfer of endostatin blocked vascular leakage and markedly suppressed NV and retinal detachment (Takahashi et al., 2003). In mice with oxygen-induced ischemic retinopathy, intraocular injection of AAV2.endostatin inhibited ischemia-induced retinal NV (Auricchio et al., 2002). These data suggest that endostatin may be an endogenous inhibitor of vasopermeability as well as a strong inhibitor of NV. Similarly, intraocular injection of angiostatin packaged in AAV or lentiviral vectors suppressed retinal and subretinal NV (Lai et al., 2001; Raisler et al., 2002; Igarashi et al., 2003).
Oxford BioMedica (Oxford, UK) has taken advantage of the large capacity of lentiviral vectors to package both endostatin and angiostatin in equine infectious anemia viral vectors (EIAV). Expression of endostatin and angiostatin suppresses leakage and growth of subretinal NV in mice (Balaggan et al., 2006; Kachi et al., 2008). A phase I trial in patients with advanced NVAMD, the Gene Transfer of Endostatin/angiostatin for Macular Degeneration Trial (GEM Study, registration no. NCT01301443;
Conclusions
VEGF is a critical stimulus in patients with NVAMD and clinical trials have shown tremendous benefit from intraocular injections of VEGF antagonists, but many patients need frequent, long-term injections to maintain benefits and prevent severe loss of vision. Lapses in treatment can be hazardous. Gene transfer provides a means of providing long-term treatment for this chronic condition with a single injection. One approach is to use gene transfer to provide sustained expression of a VEGF-binding protein. The advantage of this approach is that, based on clinical experience with intravitreous injections of VEGF antagonists, efficacy is assured as long as expression is adequate, but this is balanced by concerns regarding potential retinal toxicity from chronic suppression of VEGF. In the author's opinion, these concerns have been adequately addressed and the likelihood of toxicity is remote, but the results of a phase I clinical trial testing intraocular injection of AAV2.CBA-sFLT01 should provide a more definitive indication as to the viability of this approach.
An alternative gene transfer approach is to express one or more of a number of endogenous proteins that do not bind VEGF, but antagonize its proangiogenic and/or propermeability effects. A theoretical advantage is that if VEGF has neurotrophic activity, this approach may provide efficacy with less chance of toxicity to retinal neurons than is the case with expression of a VEGF-binding protein. Strong preclinical data and a clinical trial testing intravitreous injection of Ad.PEDF.11 have provided proof-of-concept for this approach, but it still involves more unknowns regarding efficacy and mechanisms of action than the strategy of expressing VEGF-binding proteins. The GEM Trial sponsored by Oxford BioMedica should provide valuable information regarding the safety and bioactivity of endostatin and angiostatin in patients with advance NVAMD. This trial complements the AAV2.CBA-sFLT01 trial in other important ways. Preclinical data suggest that either AAV or lentiviral vectors should provide long-term transgene expression in the eye. Because these transgenes are secreted proteins, their measurement in serial aqueous samples will provide important data regarding expression. In addition to potentially providing some assessment of the behavior of the different vector types, the expression data are critical to advance our knowledge regarding potential differences related to site of injection. Intravitreous injections are less invasive and can be done in a clinic setting, whereas subretinal injections must be done in an operating room, but immunological reactions are greater with intravitreous injections and repeat injections may be precluded with intravitreous, but not subretinal, injections (Li et al., 2008). Over the next few years, our knowledge regarding ocular gene transfer will be greatly expanded by the results of these pioneering clinical trials.
Future Considerations
The ongoing clinical trials will provide a great deal of important new information, but there are many questions that will remain unanswered.
Are there other transgenes that should be considered? There are many other antiangiogenic proteins that could be considered. Two for which preclinical gene transfer data are available are tissue inhibitor of metalloproteinases-3 (Apte et al., 1995; Della et al., 1996; Kamei and Hollyfield, 1999; Takahashi et al., 2000) and vasohibin, a negative feedback regulator of VEGF (Watanabe et al., 2004; Shen et al., 2006).
Are there other vectors that should be considered? If AAV2 and EIAV work well, this question is not urgent, but there may be other AAV serotypes, hybrid vectors, or self-complementary vectors that provide better transgene expression after intravitreous injections.
Should regulatable expression be considered? Although it would be helpful to be able to control expression levels and cease expression if necessary, this adds considerable complexity. If ongoing clinical trials show evidence of bioactivity with minimal or no toxicity, it would seem reasonable to continue to develop unregulated vectors for this application while working on regulatable expression systems in parallel for the future.
Are there other applications for gene transfer in the eye? The value of conducting trials in NVAMD is that tools are available for precise, quantitative assessment of outcomes over a relatively short period of time and results with intraocular injections of anti-VEGF agents provide a “positive control” for a successful outcome. If unequivocal benefits are seen in NVAMD, the potential applications for gene transfer in the eye are enormous.
Footnotes
Acknowledgments
This work was supported by grants EY05951, EY12609, and EY10017 from the National Eye Institute. P.A.C. is the George S. and Dolores Doré Eccles Professor of Ophthalmology.
Author Disclosure Statement
The author receives funding from Genzyme and Oxford BioMedica for conduct of clinical trials.