
Abstract
Adult human mesenchymal stromal cells (hMSCs) are an important source for tissue repair in regenerative medicine. Notably, targeted gene therapy in hMSCs to promote osteogenic differentiation may help in the development of novel therapeutic approaches for bone repair. We recently showed that α5 integrin (ITGA5) promotes osteoblast differentiation in bone marrow–derived hMSCs. Here, we determined whether lentiviral (LV)-mediated expression of ITGA5 in hMSCs derived from the bone-marrow stroma of healthy individuals may promote bone repair in vivo in two relevant critical-size bone defects in the mouse. In a first series of experiments, control or LV-ITGA5-transduced hMSCs were seeded on collagen-based gelatin sponge and transplanted in a cranial critical-size defect (5 mm) in Nude-Foxn1nu mice. Microcomputed tomography and quantitative histological analyses after 8 weeks showed no or little de novo bone formation in defects implanted with collagen sponge alone or with hMSCs, respectively. In contrast, implantation of collagen sponge with LV-ITGA5-transduced hMSCs showed greater bone formation compared with control hMSCs. We also tested the bone-repair potential of LV-mediated ITGA5 expression in hMSCs in a critical-size long-bone defect (2 mm) in femur in Nude-Foxn1nu mice. Bone remnants were stabilized with external fixation, and control or LV-ITGA5-transduced hMSCs mixed with coral/hydroxyapatite particles were transplanted into the critical-size long-bone defect. Histological analysis after 8 weeks showed that LV-ITGA5-transduced hMSCs implanted with particles induced 85% bone regeneration and repair. These results demonstrate that repair of critical-size mouse cranial and long-bone defects can be induced using LV-mediated ITGA5 gene expression in hMSCs, which provides a novel gene therapy for bone regeneration.
Introduction
Materials and Methods
Lentivirus production and transduction
The ITGA5 coding sequence (Giancotti and Ruoslahti, 1990) was amplified by PCR from pcDNA3 by using the following primers: ITGA5-ATG, 5’ CAGGGAAGAGCGGGCGCTATGG; and ITGA5-STOP, 5’ GGGAGTCTGAAATTGGGAGGACTCAGG. The amplified ITGA5 coding sequence was cloned into the pCR8/GW/TOPO TA plasmid (Invitrogen, Carlsbad, CA) and transferred into pLentiGW vector bearing the cytomegalovirus (CMV) promoter (Invitrogen) by in vitro recombination. Viral production was performed using human embryonic kidney cells HEK293T grown in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum, 1% L-glutamine, penicillin/streptomycin (10,000 U/ml and 10,000 μg/ml, respectively), and 2 mM HEPES buffer (Hamidouche et al., 2008). The day before transfection, 2×106 cells were seeded on a 175-cm2 flask. LV transfer vector (50 μg), vesicular stomatitis virus glycoprotein (VSV-G) viral envelope plasmid (pCMV-G; 10 μg), and packaging construct (pCMV DR8.2; 50 μg) were mixed with water up to 800 μl and 200 μl of 2 M CaCl2 and then added to 1 ml of HEPES-buffered saline solution 2×(5 mM NaCl, 1 mM KCl, 150 mM Na2HPO4, 0.5 mM HEPES, pH 7) and incubated for 20 min at room temperature. This DNA solution was then added dropwise onto HEK293T cells with medium, swirled gently, and then incubated overnight at 37°C. The following day, the transfection solution was removed, and the cells were rinsed with serum-free medium before addition of 15 ml of complete medium. After 24 and 48 hr of incubation, the supernatants were collected, centrifuged at 1,200 rpm to remove cell debris, filtered through a 0.45-μm low-protein binding filter (Corning, Bath, UK), aliquoted, and stored at −80°C. hMSCs were derived from the bone-marrow stroma of healthy individuals after informed consent was obtained (Delorme and Charbord, 2007). For transduction, subconfluent hMSCs were incubated with LV particles and 4 μg/ml Polybrene (Sigma, St. Louis, MO) for 48 hr in complete medium. After 48 hr, transduction medium was discarded and cells were ready for experiments. Previous data showed that all cells were transduced using this method (Hamidouche et al., 2009), and that the expression of the transgene was found to be optimal for increasing in vitro osteogenesis with a two- to threefold increase in ITGA5 expression level (Hamidouche et al., 2009).
Cranial critical-size defect
Athymic Nude-Foxn1nu mice, 8 weeks old and weighing 20–25 g (Harlan Laboratories, Jerusalem, Israel), were used for the cranial critical-size defect model. All procedures involving the use of animals were conducted in accordance with the guidelines of the Institutional Animal Care and Use Committee of the Technion, Israel. Two bilateral full-thickness circular defects (5-mm diameter) were created with a hand drill and trephine bit in the parietal bones of the skull on both sides of the sagittal suture line. Care was taken not to damage the sagittal suture or the dura mater beneath the bone. Control or LV-ITGA5-transduced hMSCs were seeded on collagen-based gelatin sponge (Levy Dental Co., Tel Aviv, Israel) cut in the shape of a disc (5×1 mm) and transplanted in the defects. Defects with collagen sponge only served as a negative control. Five replicates were made for each group (five mice per group). After 8 weeks, mice were euthanized and the harvested skulls underwent microcomputed tomography (μCT) scanning and histological analyses.
Long-bone critical-size defect
To confirm the bone-repair potential of ITGA5-expressing hMSCs, the cells were transplanted in a mouse long-bone critical-size defect model as previously described (Srouji et al., 2011). In brief, athymic Nude-Foxn1nu mice, 8 weeks old and weighing 25 g (Harlan Laboratories), were used for the experiments. Mice were anesthetized with a 0.5-ml intraperitoneal injection of xylazine and ketamine (1:1). Under aseptic conditions, a longitudinal incision was made over the lateral aspect of the thigh, and muscles were split at the fascia lata to expose the femur proximally and distally to the medial aspect of the femoral condyle. Four holes, two in the distal region and two in the proximal region, were drilled in the midshaft of the femurs. Rods were inserted manually into the drilled holes by penetration through one lateral cortex to the opposite cortex. The protruding ends of the rods on the lateral and medial sides were then connected with acrylic dental paste (Unifast Trad; GC America Inc., Alsip, IL). Under saline irrigation, a critical-size bone defect of 2 mm between the rods was created in the cylindrical midshaft part of the femur using a motorized minidrill. The muscles were opposed over the bone defect, and the wound closed with vicryl sutures (Srouji et al., 2011). Control or LV-ITGA5-transduced hMSCs mixed with coral/hydroxyapatite particles were transplanted into the critical-size defect for 8 weeks. Defects with coral/hydroxyapatite particles only served as a negative control. Five replicates were performed for each group. At the end of the experiment, mice (five per group) were euthanized, and the harvested legs were taken for histological analysis for accurate investigation of bone healing.
μCT analysis
Upon termination of the experiment, animals were euthanized, and transplanted skulls were fixed in 10% neutral buffered formalin for μCT analysis on a 40 Imaging System 6 (Scanco Medical AG, Bassersdorf, Switzerland) operated at an energy of 55 kVp and an intensity of 145 μA with an acquisition time of 200 ms and no frame averaging. Scans were performed in high-resolution mode resulting in a nominal isotropic resolution of 30 μm. A constrained Gaussian filter (sigma 1.2, support 1) was used to partly suppress the noise in the images. Mineralized bone tissue was segmented from nonmineralized tissue using a global thresholding procedure (21% of the maximum gray value) (Ruegsegger et al., 1996). The defect region was then identified by a cylindrical contour, and the bone volume was calculated within this fixed volume of interest (Müller and van Lenthe, 2006). Furthermore, we calculated the bone coverage from a projection of the superior part of the skull in the superior–inferior direction to create a high-resolution pseudoradiograph (Lutolf et al., 2003).
Histological and immunohistochemical analyses
Following μCT analysis, bone specimens were decalcified using 10% ethylenediaminetetraacetic acid, embedded in paraffin wax, and 6-μm-thick serial sections were stained with hematoxylin and eosin (H&E) for general histology. The orientation and alignment of the bones were carefully taken into consideration during paraffin embedding in order to clearly view the defect. Longitudinal serial sections of the femur and anteroposterior sections of the calvaria were stained with Masson's Trichrome for general histology and histomorphometry. The area of newly formed bone in the defect was measured on H&E-stained sections by using the ImagePro 6 image analysis software (Media Cybernetics, Silver Spring, MD); the results are expressed as a percentage of the area of the newly formed bone in the area of the original critical-size defect. The results were determined from five H&E-stained sections of each mouse (five mice per group). Unstained paraffin sections were used for immunohistochemical localization of human osteocalcin using a specific antibody (R&D Systems Inc., Minneapolis, MN) and a mouse osteocalcin antibody (Abcam, Cambridge, UK).
Statistical analysis
The results are expressed as means±SEM. Comparisons between data were performed using one-way ANOVA with Bonferroni's multiple comparison test, with p<0.05 considered as significant.
Results and Discussion
The development of efficient gene therapy that can induce osteogenic differentiation of hMSCs is of major importance for optimally promoting bone regeneration and repair. In this study, we demonstrate that bone formation and repair can be promoted in vivo by LV-mediated overexpression of ITGA5 in hMSCs. In this context, cells were transduced with a proviral copy number of the LV-ITGA5 transgene, giving rise to a two- to threefold increase in the level of expression of ITGA5 mRNA and protein (Hamidouche et al., 2009). The beneficial effect of overexpressing ITGA5 using lentivirus in hMSCs was first demonstrated by the improved bone formation in a critical-size mouse cranial defect. In this model, μCT analysis showed minimal de novo bone formation and thereby no healing in control defects in the absence of hMSC implantation after 8 weeks. We determined the effect resulting from the use of LV-transduced hMSCs to overexpress ITGA5 in bone regeneration at this time point, because earlier time points may not give sufficient information on the optimal capacity of LV-ITGA5-transduced hMSCs to fully regenerate bone. Although some de novo bone formation was observed after hMSC implantation, bone coverage in the zone of interest was only ∼38% above controls in the empty defect (Figs. 1 and 2). Remarkably, quantification of the de novo bone formed showed that the implantation of eight LV-ITGA5-transduced hMSCs showed a significantly greater bone formation compared with control hMSCs. In this condition, bone coverage was increased by ∼55% compared with control defects (Fig. 1). Histological analysis confirmed that the group with hMSCs had less and thinner newly formed bone and less coverage of the defect compared with the large amount of bone formed in the MSCs+vector group. In the latter group, the bone defect was almost completely filled with bone similar to normal calvarial bone (Fig. 2). Quantitative analysis confirmed that bone volume/tissue volume was increased significantly by 30% with hMSCs alone and significantly more with hMSCs transduced with LV-ITGA5 (47%, p<0.05), confirming the beneficial effect of LV-ITGA5-transduced hMSCs on de novo bone formation.
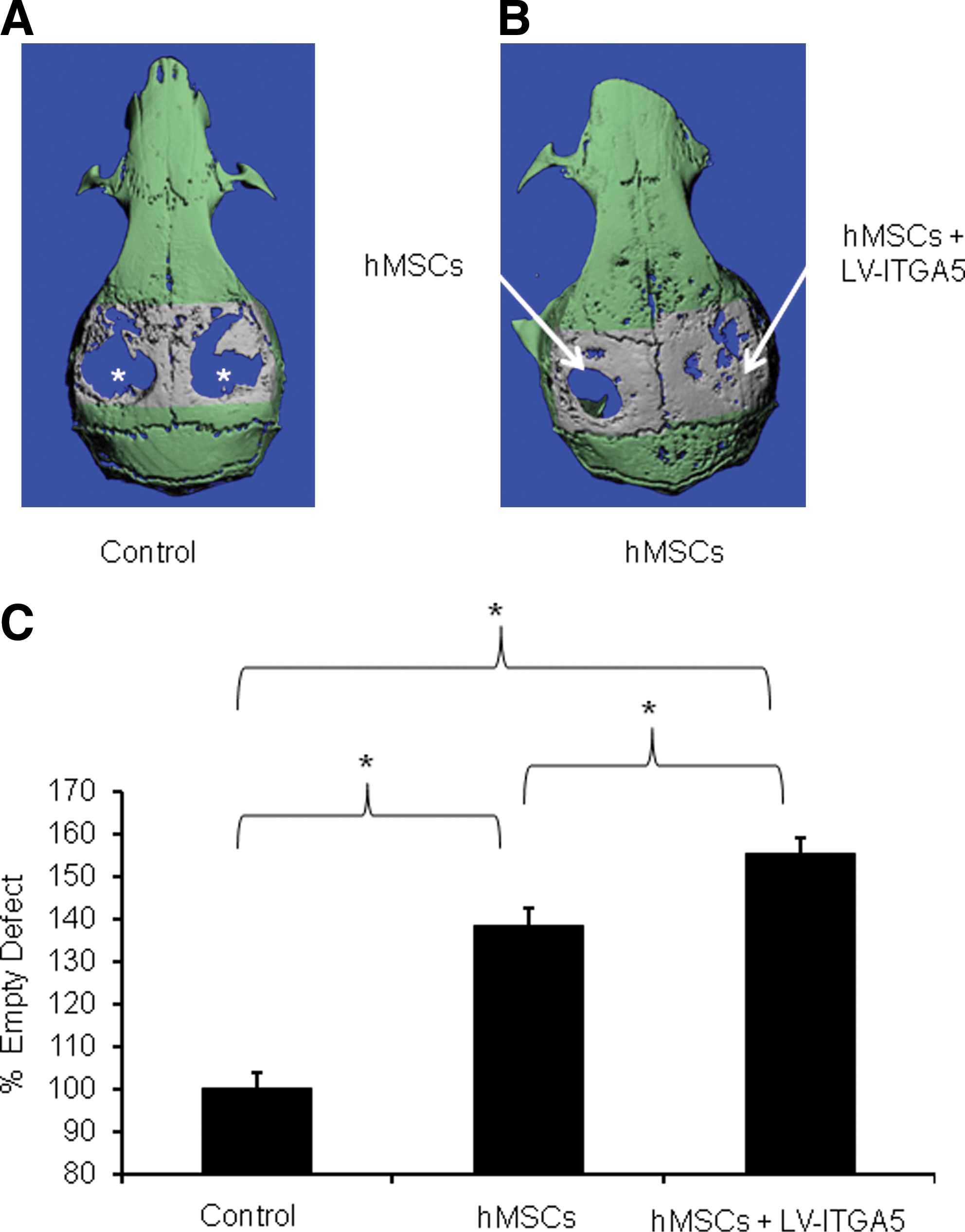
LV-mediated ITGA5 gene expression promotes bone regeneration in a critical-size cranial defect in mice. μCT analysis of critical-size cranial bone defect shows empty defects (stars) in control mice receiving collagen-based gelatin sponge alone
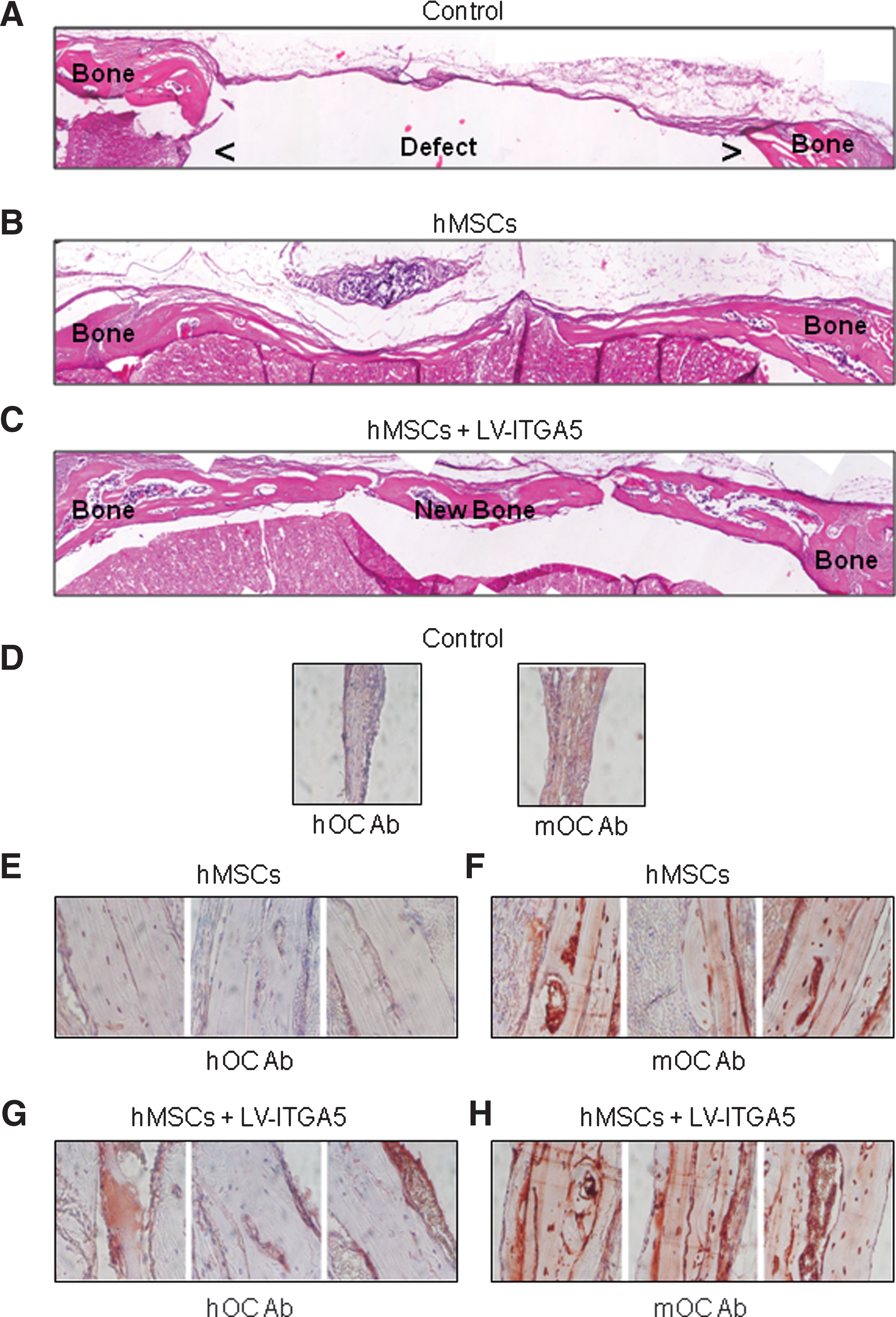
Histological and immunohistochemical analyses of bone regeneration induced by LV-mediated ITGA5 gene expression in a critical-size cranial defect in mice. Histological analysis of critical-size cranial bone defect shows empty defects in control mice receiving collagen-based gelatin sponge alone
We then performed an immunohistochemical analysis using a specific anti-mouse or anti-human antibody against osteocalcin, a late osteoblast marker, to differentiate between human and mouse cells that participated in bone repair. Immunohistochemical analysis using anti-mouse osteocalcin antibody revealed no staining in the empty defect (Fig. 2D), as expected, and a positive staining in the defect implanted with hMSCs (Fig. 2F), as expected from the presence of differentiated mouse osteoblasts in this area. Immunohistochemical analysis using anti-human osteocalcin showed no staining in the empty defect (Fig. 2D), again as expected, and a slight positive staining in the defect implanted with hMSCs (Fig. 2E). After lentiviral transduction of ITGA5 in hMSCs, osteocalcin staining was increased in implanted human cells (Fig. 2G vs. E), whereas osteocalcin staining remained unchanged in murine cells (Fig. 2H vs. F), indicating that ITGA5 overexpression in hMSCs increased osteoblast differentiation and thereby bone formation through activation of implanted human cells.
To confirm the interest of LV-ITGA5-transduced hMSCs in bone repair, we investigated whether LV-mediated expression of ITGA5 may be effective in promoting osteogenesis in a nonunion long-bone model in vivo. To this goal, we tested the effect of LV-mediated ITGA5 overexpression in hMSCs implanted in a long-bone critical-size defect in mice (Srouji et al., 2011). In this model, the implantation of coral/hydroxyapatite particles alone induced little de novo bone formation and bone repair (∼24%), as shown by histological analysis after 8 weeks of implantation. Implantation of hMSCs with coral/hydroxyapatite particles showed moderate repair (∼59%). In contrast, the implantation of LV-ITGA5-transduced hMSCs with coral/hydroxyapatite particles greatly (∼85%) increased bone formation and bone repair in this model (Fig. 3). In both calvaria and long-bone models, implanted and not endogenous MSCs participated in the bone regeneration, because in the absence of hMSCs, bone regeneration was absent or limited (Figs. 1A, 2A, and 3A). Overall, the results indicate that bone regeneration in both critical-size mouse cranial and long-bone defects are improved by ITGA5 overexpression using lentivirus in hMSCs.
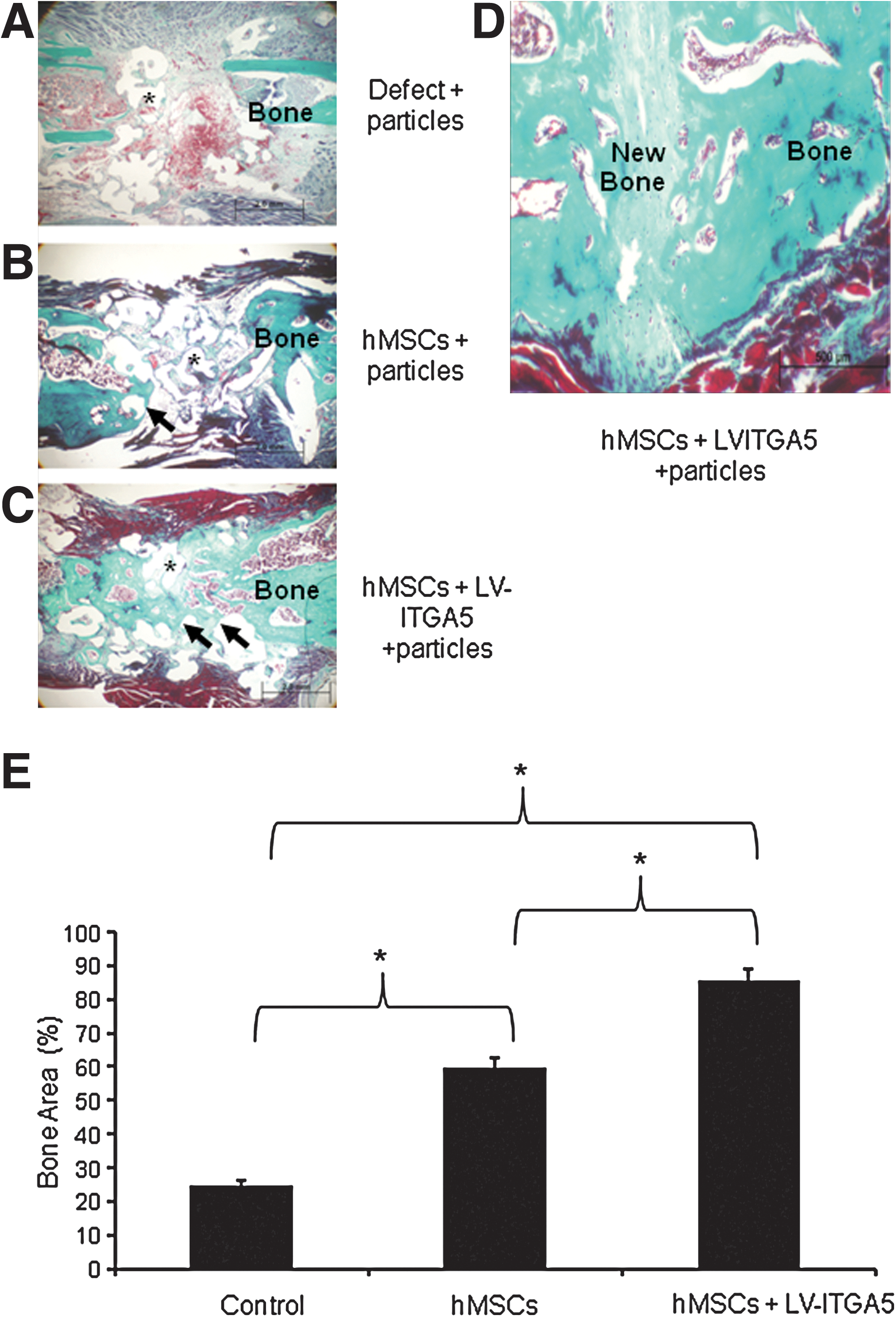
LV-mediated ITGA5 gene expression promotes bone repair in a critical-size long-bone defect in mice.
The important beneficial effects on bone formation observed in vivo with LV-ITGA5-transduction likely resulted from multiple cellular actions of ITGA5 at different steps of bone repair. Both bone regeneration and repair require the recruitment of osteoprogenitor cells and their subsequent differentiation into bone-forming cells (Einhorn, 1998). Our previous findings indicate that ITGA5 activates osteoblast differentiation in adult hMSCs through activation of focal adhesion kinase and ERK1/2-MAPKs, resulting in Runx2 phosphorylation and ectopic bone formation (Hamidouche et al., 2009). We also found that hMSC osteogenic differentiation can be induced through interactions between ITGA5 and growth factors (Hamidouche et al., 2010). Additionally, there is in vitro and in vivo evidence from our laboratory that ITGA5 acts as an antiapoptotic molecule in osteoblasts via activation of phosphatidylinositol 3-kinase signaling (Kaabeche et al., 2005; Dufour et al., 2007). The positive effect of LV-mediated overexpression of ITGA5 on cranial and long-bone repair shown here may thus result from activation of early stages of osteoblast differentiation in hMSCs and increased survival of mature osteoblasts, resulting in an increased number of osteocalcin-positive bone-forming cells, as observed in this study.
In conclusion, the clear beneficial effects of targeted LV-mediated gene therapy in the two distinct mouse models reported here support the concept that ITGA5 overexpression in hMSCs may be a valid therapeutic strategy to promote osteogenic differentiation and bone repair in vivo. These results suggest the potential for efficiently improving bone formation in unhealed cranial and nonunion long-bone defects in humans. This knowledge may serve as a basis for designing innovative gene therapy to increase the osteogenic capacity of autologous human bone marrow–derived MSCs in individuals with severe bone loss caused by trauma or tumor resection.
Footnotes
Acknowledgments
The authors thank Dr. S. Kuwada (University of Utah, Salt Lake City, UT) for the gift of the ITGA5 plasmid. This work was supported in part by the Fondation de l'Avenir pour la Recherche Appliquée, Paris, France (no. ET9-521). The use of ITGA5 agonists for applications in tissue regeneration is covered by European patent no. 08290752.8 and U.S. patent no. US-2011-0182916.
Author Disclosure Statement
No competing financial interests exist.