
Abstract
Vascular endothelial growth factor (VEGF) plays an important role in the pathogenesis of neovascular age-related macular degeneration and diabetic retinopathy. Bevacizumab, an anti-VEGF monoclonal antibody, is efficacious for these disorders, but requires monthly intravitreal administration, with associated discomfort, cost, and adverse event risk. We hypothesized that a single intravitreal administration of adeno-associated virus (AAV) vector expressing bevacizumab would result in persistent eye expression of bevacizumab and suppress VEGF-induced retinal neovascularization. We constructed an AAV rhesus serotype rh.10 vector to deliver bevacizumab (AAVrh.10BevMab) and assessed its ability to suppress neovascularization in transgenic mice overexpressing human VEGF165 in photoreceptors. Intravitreal AAVrh.10BevMab directed long-term bevacizumab expression in the retinal pigmented epithelium. Treated homozygous mice had reduced levels of neovascularization, with 90±4% reduction 168 days following treatment. Thus, a single administration of AAVrh.10BevMab provides long-term suppression of neovascularization without the costs and risks associated with the multiple administrations required for the current conventional bevacizumab monoclonal drug delivery.
Introduction
In addition to the burden on the patient and the economic burden on the health-care system, repeated intravitreal administrations pose a risk of visually devastating ocular complications. The most serious adverse event is infectious endophthalmitis. Although the per-injection rate of endophthalmitis has a reported incidence ranging between 0.03% and 0.16%, after 1–2 years of repeated injections, the per-eye infection rate approaches 1.0% (Jager et al., 2004; Brown et al., 2006; Rosenfeld et al., 2006; Elman et al., 2010). More frequent (up to 5%), although less devastating, ocular adverse events associated with repeated intravitreal administrations include vitreous hemorrhage, retinal detachment, traumatic cataract, corneal abrasion, subconjunctival hemorrhage, and eyelid swelling (Jager et al., 2004; Brown et al., 2006; Rosenfeld et al., 2006; Elman et al., 2010; Folk and Stone, 2010). In the context of these issues, an intraocular therapy with a prolonged duration of anti-VEGF action following a single intravitreal administration would decrease the treatment burden of repeated intraocular injections and reduce the cost of chronic therapy, resulting in a profound impact on the treatment of both AMD and DR.
Adeno-associated viral (AAV) vectors are attractive for ocular gene therapy, as they can direct long-term transgene expression with low toxicity and immunogenicity, with established expression in a variety of retinal cell types (Bainbridge et al., 2008; Buch et al., 2008; Roy et al., 2010; Simonelli et al., 2010; Lukason et al., 2011). There are currently six human clinical trials using intraocular administration of AAV vectors: five directed toward the treatment of Leber's congenital amaurosis (with RPE165 as the gene product), and one anti-VEGF therapy for the treatment of AMD (with soluble receptor to VEGF, sFLT01 as the gene product) (ClinicalTrials.gov, 2011). Knowing that intravitreal administration of bevacizumab is highly effective in treating AMD and DR (Avery et al., 2006; Gulkilik et al., 2010) and that of AAV gene-transfer vectors can be safely administered intraocularly to humans with persistent expression of a therapeutic transgene (Bainbridge et al., 2008; Buch et al., 2008; Roy et al., 2010; Simonelli et al., 2010; MacLachlan et al., 2011), we hypothesized that a single intravitreal administration of an AAV vector expressing bevacizumab would result in sustained intraocular expression of bevacizumab at levels sufficient for long-term suppression of ocular neovascularization. Based on AAVrh.10, a clade E, nonhuman primate (rhesus macaque)–derived gene-transfer vector that we are using in human clinical trials for gene therapy for CNS hereditary disease (Sondhi et al., 2007), we designed AAVrh.10BevMab, coding for the heavy and light chains of bevacizumab. The efficacy of AAVrh.10BevMab was tested for its ability to inhibit ocular neovascularization in the transgenic rho/VEGF mouse model that constitutively expresses the human VEGF165 isoform in photoreceptors under the rhodopsin promoter (Okamoto et al., 1997). Unlike bevacizumab itself, which is effective in this model for 14 days (Miki et al., 2009), the data demonstrate that a single administration of AAVrh.10BevMab is effective in suppressing ocular neovascularization in this murine model for at least 168 days, the longest time point evaluated.
Materials and Methods
Gene-therapy vectors
The AAVrh.10BevMab vector is based on the nonhuman primate–derived rh.10 capsid pseudotyped with AAV2 inverted terminal repeats surrounding the expression cassette consisting of cytomegalovirus (CMV)-enhancer chicken β-actin promoter (Niwa et al., 1991; Daly et al., 1999; Sondhi et al., 2007), the bevacizumab anti-human VEGF heavy-chain and light-chain sequence separated by a furin 2A self-cleavage site (Fang et al., 2005), and the rabbit α-globin polyadenylation signal (see Fig. 1A). Nucleotide sequences for the antibody heavy- and light-chain variable domains were derived from the protein sequence for human kappa Fab-12, the original humanized version of the murine mAb corresponding to bevacizumab (Chen et al., 1999). The coding sequences for the human IgG1 constant domain were added to the variable domain by overlap PCR.

Function of the AAVrh.10BevMab gene-transfer vector.
AAVrh.10BevMab was produced by cotransfection of 293orf6 cells with three plasmids: (1) an expression cassette plasmid (pAAVrh.10BevMab) carrying the humanized anti-human VEGF antibody cDNA; (2) a packaging plasmid (pAAV44.2) that contains the AAV2 rep gene and AAVrh.10 cap necessary for vector replication and capsid production; and (3) pAdDF6, an adenovirus helper plasmid (Xiao et al., 1998; Sondhi et al., 2007). For AAVrh.10 vector production, pAAVrh.10BevMab (600 μg), pAAV44.2 (600 μg), and pAdDF6 (1.2 mg) were cotransfected into 293orf6 cells, a human embryonic kidney cell line expressing adenovirus E1 and E4 genes (Gao et al., 2002; Sondhi et al., 2007), using Polyfect (Qiagen, Valencia, CA). At 72 hr post transfection, the cells were harvested, and a crude viral lysate was prepared using four cycles of freeze/thaw and clarified by centrifugation. AAVrh.10BevMab was purified by iodixanol gradient and QHP anion-exchange chromatography. The purified AAVrh.10BevMab was concentrated using an Amicon Ultra-15 100K centrifugal filter device (Millipore, Billerica, MA) and stored in PBS, pH 7.4, at −80°C. The negative control vectors AAVrh.10LacZ encodes β-galactosidase (Wang et al., 2010), AAVrh.10GFP encodes green fluorescent protein (GFP) (Sondhi et al., 2007), and AAVrh.10αV encodes an unrelated antibody against Y. pestis V antigen, which replaces the bevacizumab coding region of the AAVrh.10BevMab vector. Vector genome titers were determined by quantitative TaqMan real-time PCR analysis using a chicken β-actin promoter-specific primer–probe set (Applied Biosystems, Foster City, CA).
Assessment of AAVrh.10BevMab in vitro
Expression and specificity of the AAVrh.10BevMab-expressed bevacizumab from infected cells were assessed using western analysis. For expression, 293orf6 cells were infected with AAVrh.10BevMab [2×105 genome copies (gc)/cell], and infected cell supernatants were harvested 72 hr after infection. Supernatants were concentrated by passage through Ultracel YM-10 centrifugal filters (Millipore) and evaluated by western analysis, using a peroxidase-conjugated goat anti-human kappa light chain antibody (Sigma, St. Louis, MO) under nonreducing conditions or reducing conditions with the addition of peroxidase-conjugated goat anti-human IgG antibody (Santa Cruz Biotechnology, Santa Cruz, CA). Detection was by enhanced chemiluminescence reagent (GE Healthcare Life Sciences, Piscataway, NJ). The specificity of the AAVrh.10-expressed bevacizumab was determined by western analysis against human VEGF-165 and mouse VEGF-164 (Watanabe et al., 2008). AAVrh.10BevMab-infected cell supernatants were used as the primary antibody, followed by a peroxidase-conjugated goat anti-human kappa light-chain antibody and enhanced chemiluminescence reagent.
Bevacizumab levels after in vivo administration of AAVrh.10BevMab
Male C57BL/6 mice, 6–8 weeks of age, obtained from The Jackson Laboratory (Bar Harbor, ME), were housed under pathogen-free conditions. AAVrh.10BevMab (1011 gc) or AAVrh.10LacZ (1011 gc) in 100 μl of PBS was administered by the intravenous route to C57BL/6 mice through the tail vein. At various times 0–24 weeks after vector administration, blood was collected through the tail vein, allowed to clot for 60 min, and centrifuged at 13,000 rpm for 10 min. Bevacizumab levels in serum were assessed by enzyme-linked immunosorbent assay (ELISA) using flat-bottomed 96-well EIA/RIA plates (Corning Life Sciences, Lowell, MA) coated overnight at 4°C with 0.2 μg of human VEGF-165 per well in a total volume of 100 μl of 0.05 M carbonate buffer and 0.01% thimerosal. The plates were washed three times with PBS and blocked with 5% dry milk in PBS for 60 min. The plates were washed three times with PBS containing 0.05% Tween 20. Serial serum dilutions in PBS containing 1% dry milk were added to each well and incubated for 60 min. The positive control standard was 25 μg/μl bevacizumab (Genentech). The plates were washed three times with PBS containing 0.05% Tween 20 followed by 100 μl/well 1:5,000 diluted peroxidase-conjugated goat anti-human kappa light-chain antibody in PBS containing 1% dry milk for 60 min. The plates were washed four times with PBS containing 0.05% Tween 20 and once with PBS. Peroxidase substrate (100 μl/well; Bio-Rad, Hercules, CA) was added, and the reaction was stopped at 15 min by addition of 2% oxalic acid (100 μl/well). Absorbance at 415 nm was measured. Antibody titers were calculated with a log (OD)–log (dilution) interpolation model with cutoff value equal to twofold the absorbance of background (Watanabe et al., 2008). The titers were converted to a bevacizumab concentration using results from the bevacizumab standard data curve.
AAVrh.10BevMab (1010 gc) and AAVrh.10αV (1010 gc) in 1 μl of PBS were administered by intravitreal injection to the left and right eyes, respectively, of C57BL/6 male mice. Intravitreal injection was done under a dissecting microscope with a 32-gauge needle (Hamilton Company, Reno, NV). After 0–24 weeks, mice were killed with CO2; eyes were collected, homogenized by sonication in 100 μl of T-PER tissue protein extraction reagent (Thermo Scientific, Rockford, IL), and centrifuged at 13,000 rpm for 5 min; and supernatant was collected. Bevacizumab levels in supernatant were assessed by a human VEGF-specific ELISA as described above. Bevacizumab levels were standardized to total protein levels, which were assayed by the bicinchoninic protein assay (Thermo Scientific, Waltham, MA). The expression of bevacizumab in the eye at 12 weeks post intravitreal injection was evaluated by Western analysis as described above.
Localization of bevacizumab expression by immunofluorescence
To assess the intraocular site of bevacizumab expression, male C57BL/6 mice were injected with AAVrh.10BevMab and AAVrh.10αV, as described above, or left uninjected. Treated and control virus-injected eyes were enucleated 5 weeks after intravitreal injection, fixed in formalin, embedded in paraffin wax, sectioned, deparaffinized, and treated sequentially with biotin-conjugated donkey anti-human IgG(H+L) (dilution 1:100; Jackson ImmunoResearch, West Grove, PA) and Cy3-conjugated streptavidin (dilution 1:1,000; Jackson ImmunoResearch). Nuclei were stained with 4′6-diamidino-2-phenylindole (DAPI; dilution 1:2,000; Invitrogen, Carlsbad, CA). The sections were embedded (Histoserv, Germantown, MD) and examined with a fluorescence microscope.
AAVrh.10BevMab-mediated suppression of neovascularization
Rho/VEGF mice (a gift of Peter Campochiaro, Department of Ophthalmology and Neuroscience, The Johns Hopkins School of Medicine, Baltimore, MD) (Okamoto et al., 1997) were housed and bred under pathogen-free conditions. At postnatal day 14, homozygous rho/VEGF mice were injected intravitreally with 1 μl of PBS to one eye and 1010 gc of AAVrh.10BevMab in 1 μl to the other eye. At 2, 14, 28, 84, and 168 days post therapy, mice were anesthetized and perfused with 2 ml of 25 mg/ml fluorescein-labeled dextran (2×106 average molecular weight; Sigma, St. Louis, MO) in PBS. The eyes were removed and fixed for 1 hr in 4% paraformaldehyde/PBS. The cornea and lens were removed, and the entire retina was carefully dissected from the eyecup, radially cut from the edge of the retina to the equator in all four quadrants, and flat-mounted in Prolong Gold antifade reagent (Invitrogen). The retinas were examined by fluorescence microscopy at 200×, providing a narrow depth of field to enable subretinal focus for neovascular buds on the outer surface of the retina. AxioVision LE (Carl Zeiss, Oberkochen, Germany) digital image analysis software was used by three investigators blinded to treatment group for quantifying subretinal neovascular growth area per retina.
Statistical analysis
All data are presented as the means or geometric means±standard error. Assessment of significant effects of treatment and variability due to observer or mouse was performed using a two-factor (treatment, observer) and three-factor (treatment, observer, mouse) ANOVA model fit separately at each time point (2, 14, 28, 84, and 168 weeks). As the data were nonnormal, a nonparametric approach was used to assess significance of each factor by implementing a permutation analysis. This was conducted by fitting the ANOVA model to the data and calculating p values associated with each factor using standard parametric statistics. The data were then permuted 10,000 times and, for each permutation, the ANOVA models were fit to the permuted data and p values calculated for each of the factors. For each factor, the rank of the data p values were then determined within the ordered list of permutation p values, where the rank of the data was used to determine the nonparametric p value. Overall, for the three-factor ANOVA, we conducted three tests at each of the five time points to produce 15 independent tests, whereas the two-factor ANOVA produced 10 tests that were highly nonindependent of the three-factor ANOVA tests. We therefore considered cases where p<0.05/15=0.0033 to be significant after a multiple test correction, e.g., we consider p>0.0033 as providing no evidence of significant effect of treatment or variability in a factor.
Results
Characterization of AAVrh.10BevMab
The AAVrh.10BevMab vector was tested for in vitro expression of human heavy and light chain by infection of 293orf6 cells (Fig. 1). Cell culture supernatant at 72 hr post infection, assessed by Western analysis under nonreducing and reducing conditions, established expression of the intact heavy and light chains and their ability to form the intact antibody (Fig. 1B and C). Infection with the control AAVrh.10GFP vector under identical conditions had no detectable bands, reduced or nonreduced, for human antibody. The supernatant from AAVrh.10BevMab-infected cells was tested for the capacity to specifically recognize human VEGF by probing a western against human VEGF165 and mouse VEGF164 (Fig. 1D). Only the human form of VEGF was recognized as expected from the known specificity of bevacizumab. In contrast, supernatants from AAVrh.10GFP-infected cells did not recognize either protein. To assess the ability of the AAVrh.10BevMab vector to direct persistent expression of bevacizumab in vivo, serum antibody levels of human VEGF were assessed by ELISA at 0–24 weeks after intravenous administration of 1011 gc of the AAVrh.10BevMab. Antibody levels peaked at about 12 weeks and were sustained through the 24 weeks (Fig. 1E), the last time point examined. No antibody was detected in serum of mice that received similar intravenous injection of AAVrh.10LacZ (control vector expressing β-galactosidase).
Intravitreal administration of AAVrh.10BevMab
To evaluate the expression profile of AAVrh.10BevMab by local administration, antibody levels in the supernatant of eye homogenate were assessed for 0–24 weeks after intravitreal administration of 1010 gc of the vector. The data showed that the bevacizumab levels were above 100 pg/μg total protein at 2 weeks and remained at similar levels to the last time point evaluated at 24 weeks (Fig. 2A). No bevacizumab was detected in the eyes from mice that received intravitreal AAVrh.10αV, a control vector. The expression of bevacizumab in the eye post intravitreal administration of AAVrh.10BevMab was confirmed by Western analysis. Soluble protein collected from the AAVrh.10BevMab-injected eyes was positive for the presence of human antibody heavy and light chains, whereas no human antibody was detected in eyes injected with AAVrh.10αV, which expresses a mouse mAb, or uninjected naive eyes (Fig. 2B).
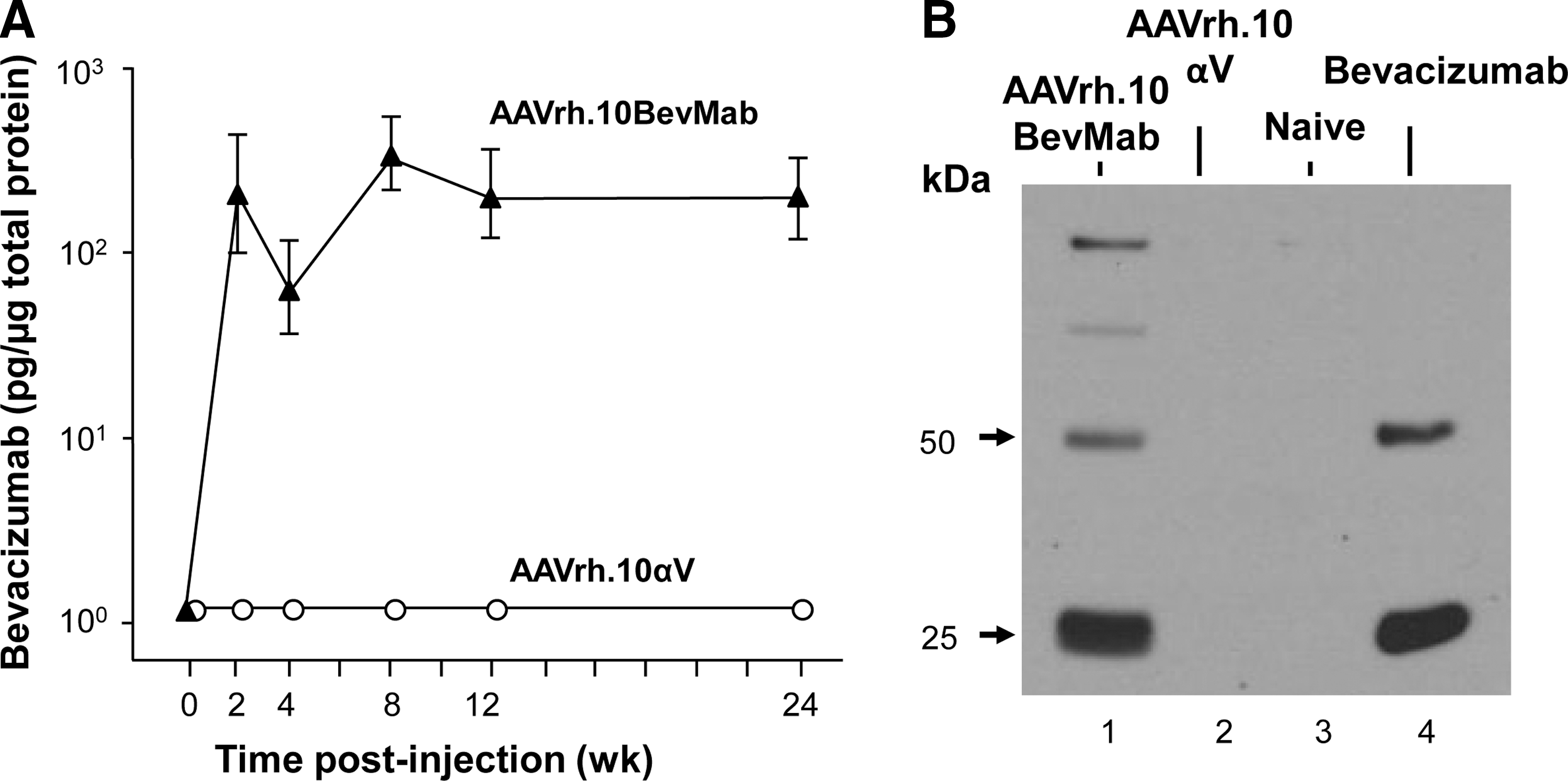
Ocular levels of bevacizumab after a single local administration of the AAVrh.10BevMab vector. AAVrh.10BevMab (1011 gc) was administered to C57BL/6 mice by the intravitreal route, with AAVrh.10αV, an AAVrh.10 vector coding for an irrelevant IgG antibody, as a control.
The localization of bevacizumab within the eyes of mice administered intravitreal AAVrh.10BevMab was evaluated in paraffin-embedded and stained sections of eyes 35 days post injection. The bevacizumab was localized to the retinal pigment epithelium (RPE), whereas no bevacizumab staining was seen in uninjected eyes or eyes injected with the vector expressing the mouse mAb, AAVrh.10αV (Fig. 3). Intravitreal administration of AAVrh.10 has previously been reported to efficiently transduce a wide range of retinal cells, including the RPE, the ganglion cell layer, the amacrine cells of the inner nuclear layer, the Müller and horizontal cells, as well as bipolar cells (Giove et al., 2010). Therefore, we searched multiple immunohistochemical sections for staining of these cell types, but have observed no staining in any cell type other than RPE.
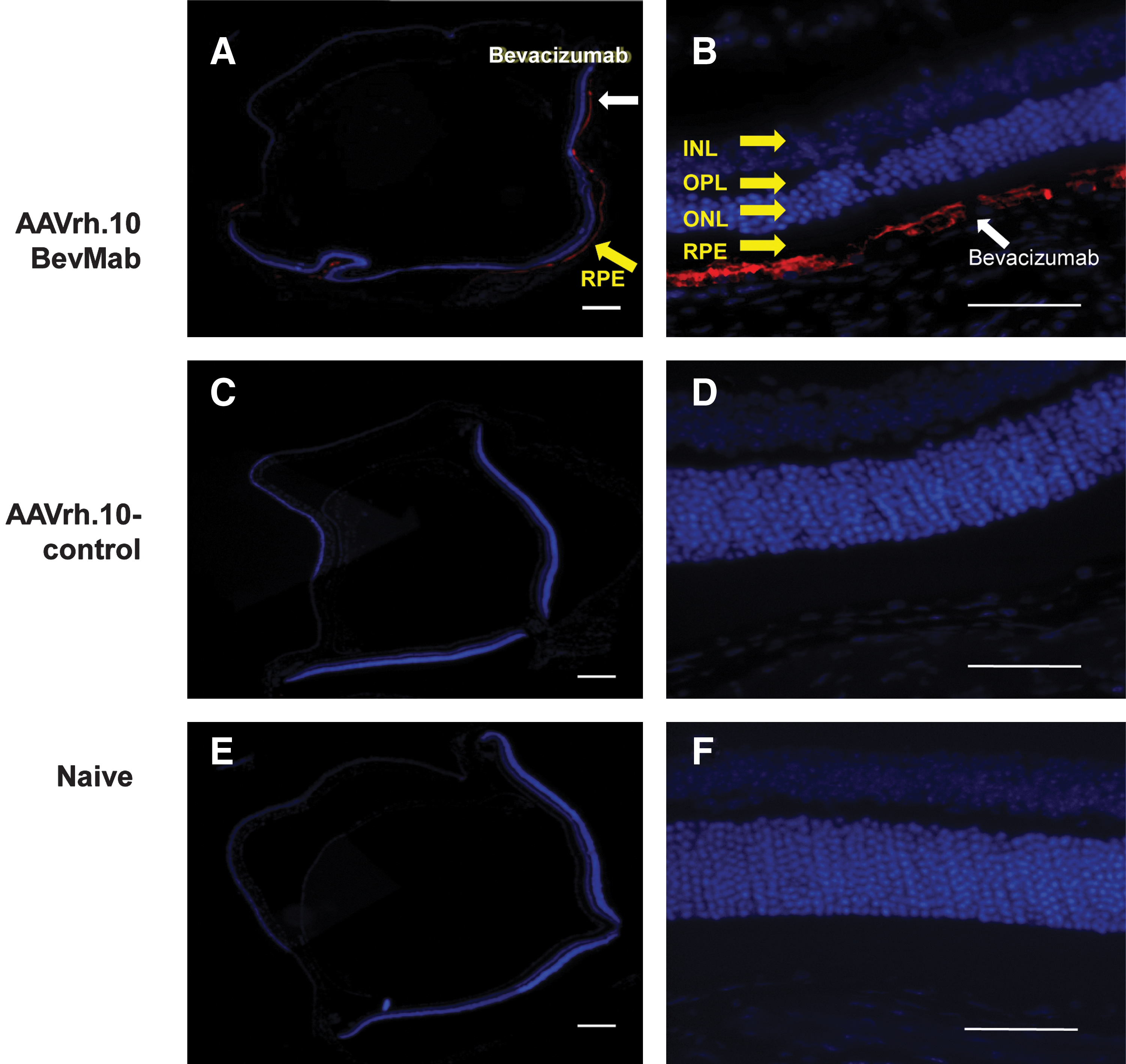
Immunofluorescence localization of bevacizumab after local ocular administration of AAVrh.10BevMab. AAVrh.10BevMab (1010 gc) or the control vector AAVrh.10αV (1010 gc) in 1 μl of PBS was administered by the intravitreal route to C57BL/6 mice. Five weeks later, eyes were enucleated, and paraffin sections were stained using biotin-conjugated donkey anti-human IgG as primary antibody and Cy3-conjugated streptavidin as secondary antibody. Nuclei were stained with DAPI. Untreated control eyes were stained in the same way. Bevacizumab is indicated by red, and nucleus by blue. INL, inner nuclear layer; ONL, outer nuclear layer; OPL, outer plexiform layer; RPE, retinal pigment epithelium. Scale bars=200 μm
Efficacy of AAVrh.10BevMab
The efficacy of the AAVrh.10BevMab in the transgenic rho/VEGF mouse was assessed by evaluating the suppression of the development of neovascular buds following intravitreal injection to the eyes of 14-day-old homozygous rho/VEGF mice. As an internal control, the opposite eye received an intravitreal injection of PBS. Fluorescence microscopy of the retina from AAVrh.10BevMab- and PBS-injected eyes at 2, 14, 28, 84, and 168 days post injection was used to assess the phenotype (Fig. 4). In low-magnification views (Fig. 4A), multiple large areas of budding and vascular leak are evident in the PBS-treated eye of the mice at 168 days post injection; these areas were largely absent in the treated eye (Fig. 4B). By examining neovascular buds at higher power (Fig. 4C), the time-dependent increase in budding was seen. At 2 days post injection, AAVrh.10BevMab- and PBS-injected eyes appeared to have similar amounts of neovascular buds, but at longer time points, AAVrh.10BevMab-injected eyes had significantly fewer subretinal neovascular buds than retinas from eyes injected with PBS.
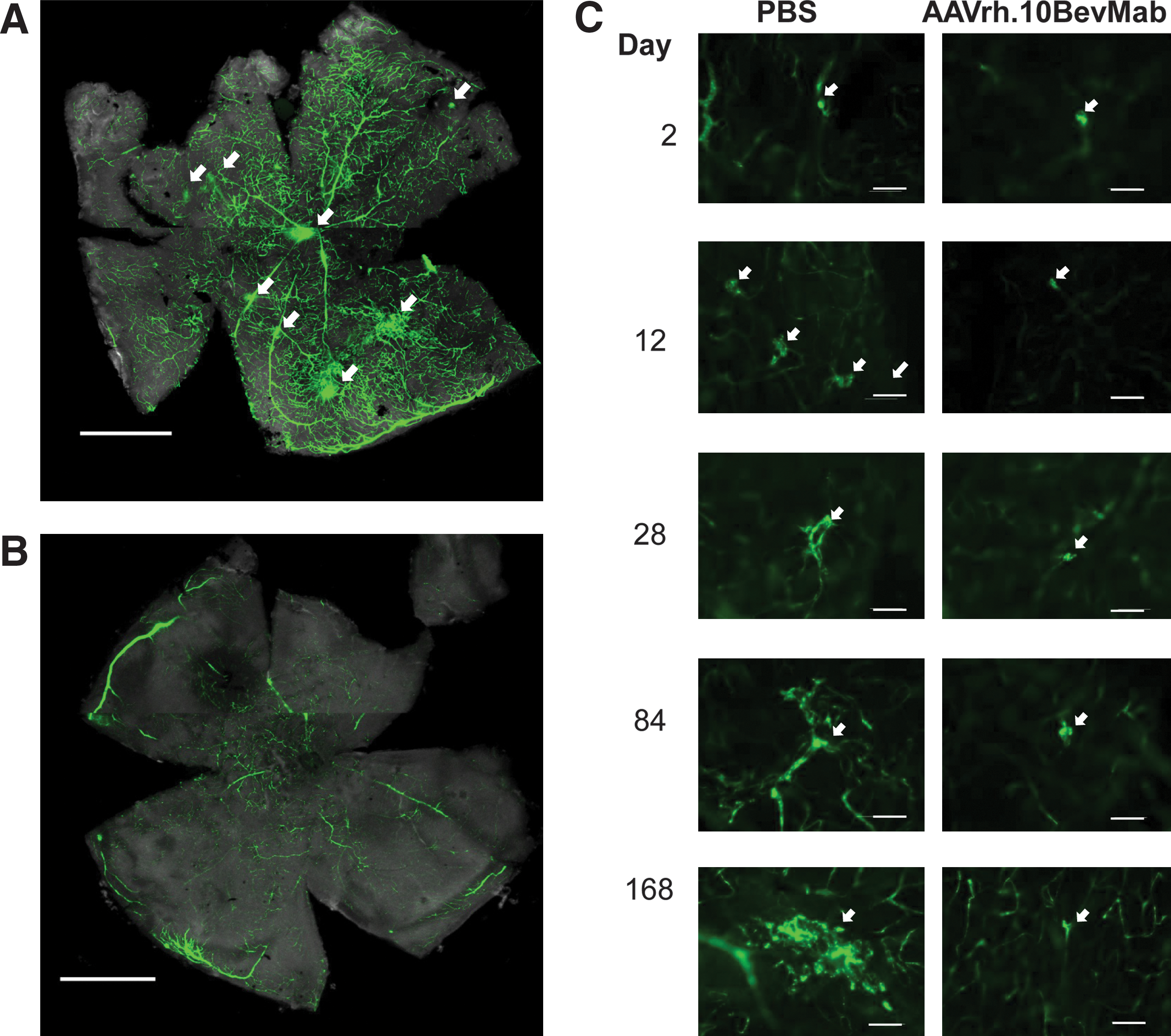
Ability of AAVrh.10BevMab to suppress neovascularization in rho/VEGF transgenic mice with overexpression of VEGF in photoreceptors. At postnatal day 14, rho/VEGF mice were given an intravitreal injection of 1 μl of PBS in one eye and 1 μl containing 1010 gc AAVrh.10BevMab in the other eye. At 2, 14, 28, 84, and 168 days post injection, the total area of subretinal neovascularization per eye was quantified from retinal flatmount.
The subretinal neovascular buds were quantified by three investigators blinded to treatment group. As an example of the individual data from each observer, at 84 days post injection, the data from each of the observers showed significant reduced area of subretinal neovascular buds in retinas of AAVrh.10BevMab-injected eyes compared with eyes injected with PBS (Fig. 5A). The interobserver variability in quantifying the neovascular buds was not significant at the multiple test correction threshold (Table 1). Data from the three observers was first averaged for each eye, and then the average and standard error for each condition and time point were plotted (Fig. 5B). Consistent with the fluorescence microscopy results, at 2 days post injection there was no significant reduction in the area of subretinal neovascular buds for AAVrh.10BevMab-injected eyes, but from 14 to 168 days after therapy, eyes injected with AAVrh.10BevMab had significantly less area of subretinal neovascular buds compared with retinas from eyes injected with PBS (Fig. 5B). The reduction ratio—calculated as [(mean neovascular bud area in PBS-injected eye at indicated time point) – (neovascular bud area in AAVrh.10BevMab-injected eye at indicated time point)]/(mean neovascular bud area in PBS-injected eye)—showed no reduction at 2 days post injection, but significant reduction at later time points: 14 days (49%) to 168 days (90%) (Fig. 5C). Thus, a single intravitreal administration of AAVrh.10BevMab can persistently suppress subretinal neovascularization in this model.
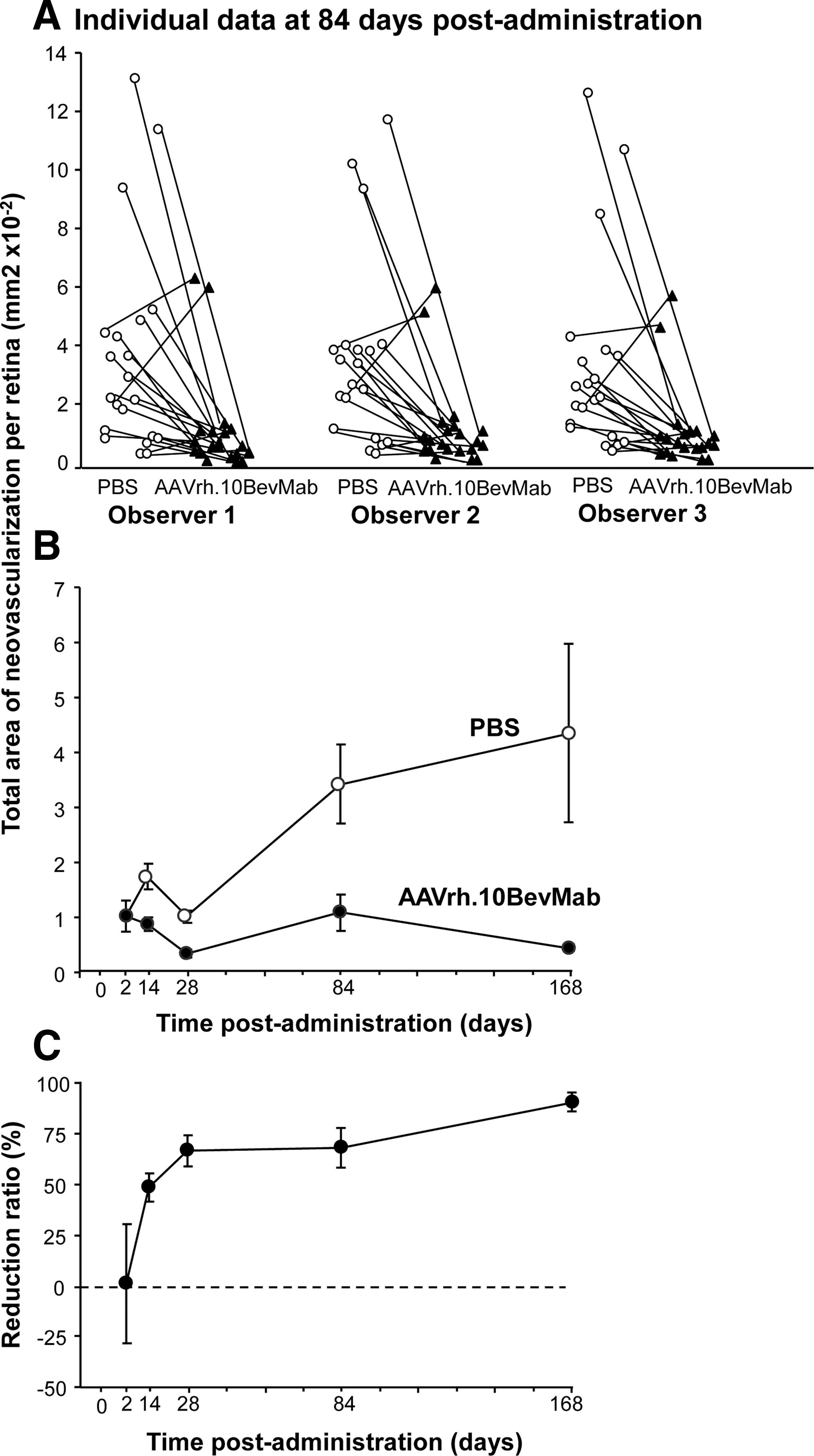
AAVrh.10BevMab-mediated suppression of neovascularization compared with PBS. At postnatal day 14, homozygous rho/VEGF mice were injected intravitreally with 1 μl of PBS to one eye and 1010 gc of AAVrh.10BevMab in 1 μl to the other eye. At 2, 14, 28, 84, and 168 days post therapy, the total area of subretinal neovascularization per eye was quantified by retinal flatmount.
Observer means and standard deviations were calculated after summing over mice. The effects of treatment, observer, and mouse were assessed using permutations after fitting a two-factor and three-factor ANOVA model.
NV, neovascularization.
p<0.05, but not significant after a multiple test correction.
Significant test results.
Discussion
Intraocular anti-VEGF therapy with bevacizumab or its Fab fragment ranibizumab, the standard worldwide therapy for the treatment of AMD and DR, requires monthly intravitreal administration to maintain optimal visual outcomes (Avery et al., 2006; Brown et al., 2006; Rosenfeld et al., 2006; Regillo et al., 2008; Elman et al., 2010; Folk and Stone, 2010; Gulkilik et al., 2010; Mitchell et al., 2010; Nicholson and Schachat, 2010; Schmidt-Erfurth et al., 2010; Waisbourd et al., 2010). To circumvent the discomfort to the patient of monthly intravitreal injections, the economic burden of this expensive therapy, and the uncommon, but serious, ocular complication rate associated with repeated intravitreal administration, we have devised an AAV-mediated gene-therapy strategy to deliver persistent therapeutic levels of bevacizumab to the eye using a single administration. Using AAVrh.10BevMab, a rhesus-derived AAV coding for bevacizumab, the data demonstrate that AAVrh.10BevMab expresses intact functional bevacizumab that, with a single intravitreal administration, provides effective therapy in a murine model of VEGF-medicated ocular neovascularization for at least 24 weeks, the longest time point evaluated.
Anti-VEGF therapy for ocular neovascularization
DR, the primary cause of permanent blindness in adults aged 20–65 years, and AMD, the main cause of irreversible blindness in those over age 65, are predominantly VEGF-mediated pathological processes (Aiello et al., 1994; Lu and Adamis, 2006; Ferrara, 2010). Up-regulation of VEGF within the eye leads to retinal neovascularization in DR and choroidal neovascularization in exudative AMD (Aiello et al., 1994; Lu and Adamis, 2006; Ferrara, 2010). This pathological neovascularization results in increased vascular permeability producing retina edema, vascular fragility leading to hemorrhage, and fibrovascular proliferation and scarring—all ultimately culminating in severe vision loss. Suppression of this VEGF-driven proangiogenic cycle has revolutionized the treatment of DR and AMD (Schlingemann and Witmer, 2009; Ferrara, 2010; Nicholson and Schachat, 2010; Waisbourd et al., 2010). Numerous clinical trials have definitively demonstrated that, when compared with previously available therapies, intravitreal administration of anti-VEGF agents (either the full-length antibody bevacizumab or its Fab fragment ranibizumab) considerably improves visual outcome in these patients (Avery et al., 2006; Brown et al., 2006; Rosenfeld et al., 2006; Regillo et al., 2008; Schlingemann and Witmer, 2009; Elman et al., 2010; Ferrara, 2010; Folk and Stone, 2010; Gulkilik et al., 2010; Mitchell et al., 2010; Nicholson and Schachat, 2010; Schmidt-Erfurth et al., 2010; Waisbourd et al., 2010). For example, in the two pivotal ranibizumab trials in patients with exudative AMD (ANCHOR and MARINIA), on average, all patients in the standard-of-care group lost vision at the 1- and 2-year time points. This is in dramatic contrast to those patients receiving monthly intraocular anti-VEGF injections, where 95% of patients maintained, and 30–40% improved, their vision at the 1- and 2-year time points (Brown et al., 2006; Rosenfeld et al., 2006; Mitchell et al., 2010). Thus, anti-VEGF therapy has become the standard of care for these ocular disorders.
However, the relatively short half-life of intraocularly administered proteins (including bevacizumab and ranibizumab) necessitates frequent and repeated administration to maintain adequate therapeutic levels. When compared in AMD and DR clinical trials, patients treated less frequently (either quarterly or on an “as needed” basis) showed less pronounced benefit of visual outcomes (Regillo et al., 2008; Mitchell et al., 2010; Schmidt-Erfurth et al., 2010). Patients switched from monthly therapy to an “as needed” anti-VEGF therapy regimen had, on average, greater visual decline compared with a monthly treatment regimen (Regillo et al., 2008; Mitchell et al., 2010; Schmidt-Erfurth et al., 2010). Without prolonged VEGF blockage via repeated intraocular injections of currently available anti-VEGF agents, the proangiogenic process is reactivated and visual outcome is compromised. However, these repeated injections come at the high cost of a cumulative risk of visually devastating complications, including intraocular hemorrhage, retinal detachment, and endophthalmitis (Jager et al., 2004; Schmidt-Erfurth et al., 2010). For example, when the anti-VEGF clinical trials are taken together, over a 1–2-year period of repeated intravitreal injections, the per-eye risk of intraocular infection approaches 1% (Brown et al., 2006; Rosenfeld et al., 2006; Elman et al., 2010). With some patients potentially requiring decades-long VEGF suppression, the local complication rate becomes a significant impediment to the adequate treatment of pathological ocular neovascularization.
AAV-mediated anti-VEGF gene therapy
One rational approach to achieve a sustained therapeutic effect following a single intraocular injection is that of AAV-mediated gene transfer. As a compartmentalized, self-contained, easily assessable, relatively small, immune privileged organ, the eye is an ideal site for in vivo gene transfer (Bainbridge et al., 2003). Ocular gene-transfer strategies have been developed for gene-augmentation therapy in recessively inherited disorders (e.g., autosomal recessive and X-linked recessive retinitis pigmentosa, Usher syndrome, X-linked retinoschesis, Leber's congenital amaurosis); for gene silencing in dominantly inherited disorders (e.g., autosomal dominant retinitis pigmentosa, retinoblastoma); and for treatment of ocular pathologic processes (e.g., DR and AMD) (Roy et al., 2010). For example, the clinical trial of rAAV2-CB-hRPE65 in patients with Leber's congenital amaurosis demonstrated the safety and efficacy of ocular gene therapy with clinically quantifiable improvements in visual function up to 1.5 years following a single injection, without any serious ocular or systemic side effects (Simonelli et al., 2010). Given the accumulating evidence for the safety and efficacy of ocular gene therapy, our approach to the long-term suppression of ocular neovascularization in DR and AMD was that of AAV-mediated gene-transfer strategy.
Although all human studies on AAV gene transfer to the eye have involved serotype 2 (Hauswirth et al., 2008; Simonelli et al., 2010), there is ample evidence in animals that other AAV serotypes derived from nonhuman primates provide higher levels of transgene expression (Lebherz et al., 2008). AAVrh.10 is derived from rhesus macaque and is extremely effective in gene transfer to pleura and brain (De et al., 2006; Sondhi et al., 2007). In a previous study, intravitreal injection into the eye of AAVrh.10 expressing enhanced GFP efficiently transduced several cell types of the retina, including cells of the outer plexiform layer, the ganglion cell layer, and the RPE (Giove et al., 2010). In the present study, bevacizumab delivered by AAVrh.10 accumulated exclusively in the RPE. This apparent inconsistency may reflect several factors, including the use of an intracellular (GFP) versus a secreted transgene (bevacizumab) and mouse strain differences. In addition, the receptor for AAVrh.10 is unknown and the specific factors affecting tropism are not defined.
Other gene-therapy approaches for the treatment of pathologic ocular neovascularization have included viral vectors that encode various antiangiogenic agents, including endostatin, angiostatin, pigment epithelial-derived factor, and sFLT01 (Bainbridge et al., 2003; Roy et al., 2010). A phase 1 clinical trial using intravitreal AAV2-sFLT01 in patients with AMD has just recently been completed. Although no dose-limiting toxicities have been publicly reported in this trial, it is not known whether sFLT01 itself has sufficient antiangiogenic activity to provide similar visual outcomes to currently available anti-VEGF therapeutics. Anti-VEGF therapy with bevacizumab, on the other hand, is an existing, well-characterized, therapeutic agent with proven efficacy and an acceptable safety profile following repeated intravitreal injections in patients with DR and AMD (Avery et al., 2006; Brown et al., 2006; Rosenfeld et al., 2006; Regillo et al., 2008; Singerman et al., 2008; Schlingemann and Witmer, 2009; Elman et al., 2010; Ferrara, 2010; Folk and Stone, 2010; Gulkilik et al., 2010; Mitchell et al., 2010; Nicholson and Schachat, 2010; Schmidt-Erfurth et al., 2010; Waisbourd et al., 2010). Therefore, our approach was to use bevacizumab rather than another agent in the AAV-mediated intraocular antiangiogenesis gene therapy and improve the only known shortcoming of this therapy: persistence of action. Toward that end, we engineered an AAV vector that encodes the antibody heavy- and light-chain variable domains of the humanized version of the murine mAb corresponding to bevacizumab (AAVrh.10BevMab).
In vitro analysis of our AAVrh.10BevMab vector product showed it to have equivalent VEGF-binding properties to native bevacizumab used clinically. Following a single intravitreal injection, the vector was able to transfect the RPE cells and to produce bevacizumab. Intraocular bevacizumab levels were detected by day 14 and remained elevated up to 24 weeks, the last time point tested. In vivo efficacy of our AAVrh.10BevMab vector was tested in a well-characterized VEGF-mediated ocular neovascularization model that mimics critical features of neovascular AMD. A single intravitreal injection of bevacizumab in this model typically suppresses ocular neovascularization for 2 weeks. Using our AAVrh.10BevMab vector, we were able to meaningfully suppress ocular neovascularization following a single intravitreal injection for up to 24 weeks, the last time point tested.
Ocular gene-therapy strategies have used either intravitreal or subretinal approaches for intraocular delivery of the viral vector (Bainbridge et al., 2003, 2008; Roy et al., 2010; Lukason et al., 2011). In current clinical practice, however, intravitreal injections are used for anti-VEGF treatment of AMD and DR. Intravitreal injections are less invasive than subretinal injections, have a considerably more favorable side-effect profile, and can be performed in an office setting. Subretinal injections, on the other hand, require the patient to undergo a surgical procedure. Hence, our approach for long-term suppression of ocular neovascularization was that of an intravitreal injection route. This was the same strategy used for the AAV2-sFLT01 preclinical and phase 1 studies (ClinicalTrials.gov, 2011; Lukason et al., 2011; MacLachlan et al., 2011). Similar to the AAV2-sFLT01 data, the results of our approach include retinal cell transfection, localized transgene expression, and suppression of neovascularization.
One potential criticism of using AAVrh.10BevMab for ocular gene therapy for AMD and DR is the theoretical deleterious effect of prolonged ocular VEGF suppression. Although these warrant careful surveillance and further investigation, clinical experience with hundreds of thousands of patients who have received continued intraocular anti-VEGF therapy over many years (up to 3 years with the clinically available intraocular anti-VEGF therapies) have yet to show any detrimental consequences of prolonged VEGF blockage (Avery et al., 2006; Brown et al., 2006; Rosenfeld et al., 2006; Regillo et al., 2008; Singerman et al., 2008; Schlingemann and Witmer, 2009; Elman et al., 2010; Ferrara, 2010; Folk and Stone, 2010; Gulkilik et al., 2010; Mitchell et al., 2010; Nicholson and Schachat, 2010; Schmidt-Erfurth et al., 2010; Waisbourd et al., 2010). In fact, the opposite appears to be true, as patients who are not treated with anti-VEGF therapy ultimately end up with significantly worse visual function. Along the same lines, the most recent evidence on long-term ocular VEGF suppression in several animal models has shown no abnormalities of the choriocapillaris and no indication of retinal cell dysfunction (Singerman et al., 2008; Ueno et al., 2008). At this time, most experimental and all clinical evidence indicates that long-term VEGF suppression is preferred in patients with AMD and DM.
This study provides the first report of sustained suppression of ocular neovascularization using a well-characterized, existing therapeutic approach with proven clinical efficacy. With the goal of minimizing dosing intervals and maximizing clinical efficacy, a single administration of AAVrh.10BevMab provides long-term expression of bevacizumab and suppression of VEGF-mediated ocular neovascularization. The results of this study warrant the further investigation of AAVrh.10BevMab as a long-term therapeutic approach for the treatment of AMD and DR.
Footnotes
Acknowledgments
We thank Stacey Chen for help with the analysis and N. Mohamed for help in preparing the manuscript. These studies were supported, in part, by P50 HL084936, T32 HL094284, and UL1-RR024996, and Research to Prevent Blindness, New York, NY.
Author Disclosure Statement
No competing financial interests exist.