
Abstract
Current methods of gene transfer for heart disease include injection into heart muscle or intracoronary coronary delivery, approaches that typically provide limited expression and are cumbersome to apply. To circumvent these problems, we selected a transgene, insulin-like growth factor-I (IGF-I), which may, in theory, have favorable effects on heart function when secreted from a remote site. We examined the feasibility and efficacy of skeletal muscle injection of adeno-associated virus 5 encoding IGF-I under Tet regulation (AAV5.IGFI-tet) to treat heart failure. Myocardial infarction (MI) was induced in rats by coronary occlusion; 1 week later, rats with impaired left ventricular (LV) function received 2×1012 genome copies (GC) of AAV5.IGFI-tet in the anterior tibialis muscle, and 4 weeks later, were randomly assigned to receive doxycycline in drinking water to activate IGF-I expression (IGF-On; n=10), or not to receive doxycycline (IGF-Off; n=10). Ten weeks after MI (5 weeks after activation of IGF-I expression), LV size and function were assessed by echocardiography and physiological studies. IGF-On rats showed reduced LV end-systolic dimension (p=0.03) and increased LV ejection fraction (p=0.02). In addition, IGF-On rats showed, before and during dobutamine infusion, increases in cardiac output (p=0.02), stroke work (p=0.0001), LV + dP/dt (p<0.0001), LV relaxation (LV – dP/dt; p=0.03), and systolic arterial blood pressure (p=0.0003). Mean arterial pressure and systemic vascular resistance were unchanged. Activation of IGF-I expression reduced cardiac fibrosis (p=0.048), apoptosis (p<0.0001), and caspase-3/7 activity (p=0.04). Serum IGF-I was increased 5 weeks after transgene activation (p=0.008). These data indicate that skeletal muscle injection of AAV5.IGFI-tet enables tetracycline-activated expression, increases serum IGF-I levels, and improves function of the failing heart.
Introduction
Current methods of gene transfer for heart disease include intramuscular injection into heart muscle or intracoronary delivery, approaches that provide limited expression or are cumbersome to apply. A simple skeletal muscle injection of a vector encoding a paracrine-type transgene with beneficial cardiac effects might circumvent these problems, enabling patients with severe CHF to be treated in an outpatient setting. To develop and test such an approach was the purpose of the present study.
The ideal vector for such a paracrine-based gene transfer strategy would include the following: (1) a long-term expression vector suitable to treat chronic CHF; (2) a transgene with multiple beneficial cardiac effects that is secreted into the systemic circulation after skeletal muscle injection; and (3) regulated expression enabling (a) turning on and off transgene expression quickly in case of undesirable effects, which would be important in clinical settings, and (b) activation of transgene expression in the presence of severe CHF to test its efficacy in a stringent manner. The vector best suited to fulfill these goals is the adeno-associated virus (AAV), which provides long-term expression after skeletal muscle delivery (Xiao et al., 2000). The gene we selected for the proposed studies was insulin-like growth factor-I (IGF-I), which has pleiotropic effects on the heart that are likely to benefit patients with CHF. IGF-I has positive inotropic effects (Cittadini et al., 1998), reduces cardiac myocyte apoptosis (Buerke et al., 1995), promotes cardiac myocyte growth (Serneri et al., 2001), and is angiogenic (Duerr et al., 1996). Last, the tetracycline (Tet) regulation system would provide a suitable means to control transgene expression (Yu et al., 1996; Gao et al., 2002). Using a long-term regulated expression vector in CHF would enable us to activate IGF-I expression when CHF is severe, providing a stringent test of efficacy that embodies the challenge faced in clinical gene therapy. We therefore tested the hypothesis that skeletal muscle delivery of an AAV5 vector encoding IGF-I, under Tet regulation (AAV5.IGFI-tet), would increase function of the failing heart.
Materials and Methods
Regulated expression of IGF-I
An AAV5 vector encoding rat IGF-I (type A) under Tet regulation (see Fig. 1A) was constructed for these studies. The coding sequence included a signal peptide for extracellular secretion and an AU1 tag to facilitate detection. To determine the time course of doxycycline-activated IGF-I expression, cultured neonatal rat cardiac myocytes underwent gene transfer with AAV5.IGFI-tet (104 GC/cell, 2 days). Subsequently, doxycycline (2 μg/ml) was added to the medium, and IGF-I mRNA expression was quantified sequentially by real-time RT-PCR before and after doxycycline removal, which was achieved by multiple phosphate-buffered saline (PBS) washes. To detect transgene IGF-I protein expression, cultured neonatal rat cardiac myocytes were incubated with AAV5.IGFI-tet (104 GC/cell) and activated with doxycycline (2 μg/ml, 3 days), with subsequent detection of IGF-I in medium by Western blotting with anti-AU1 antibody (Covance, Princeton, NJ). In addition, Akt activation was evaluated in cell lysates by Western blotting with anti-Akt and anti-Akt-P antibodies (Cell Signaling Technology, Danvers, MA). Validation of transgene expression after intramuscular injection was performed with AAV5 encoding enhanced green fluorescent protein (EGFP) (see Fig. 2A).
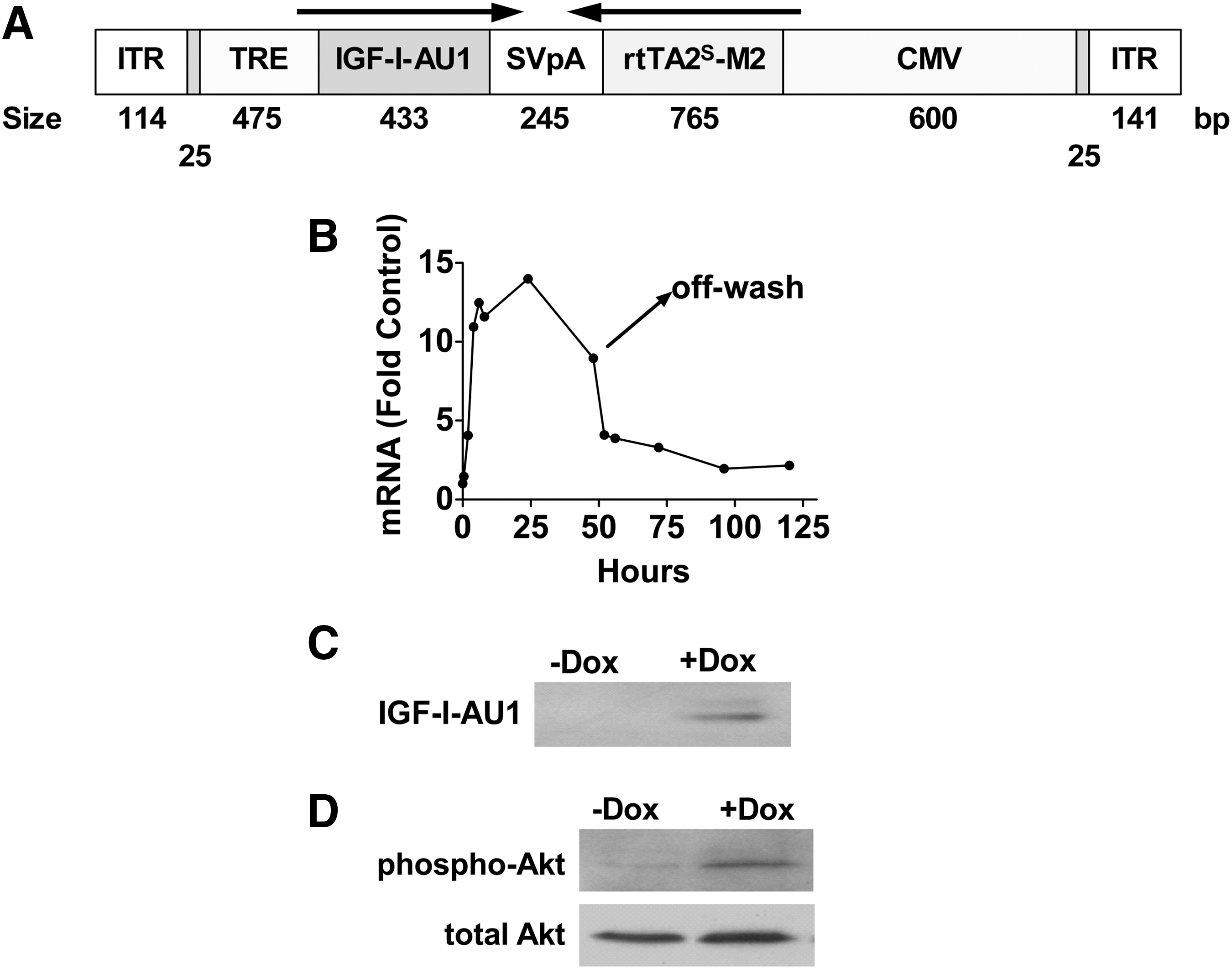
AAV5.IGFI-tet vector map and regulated expression in cultured cardiac myocytes.
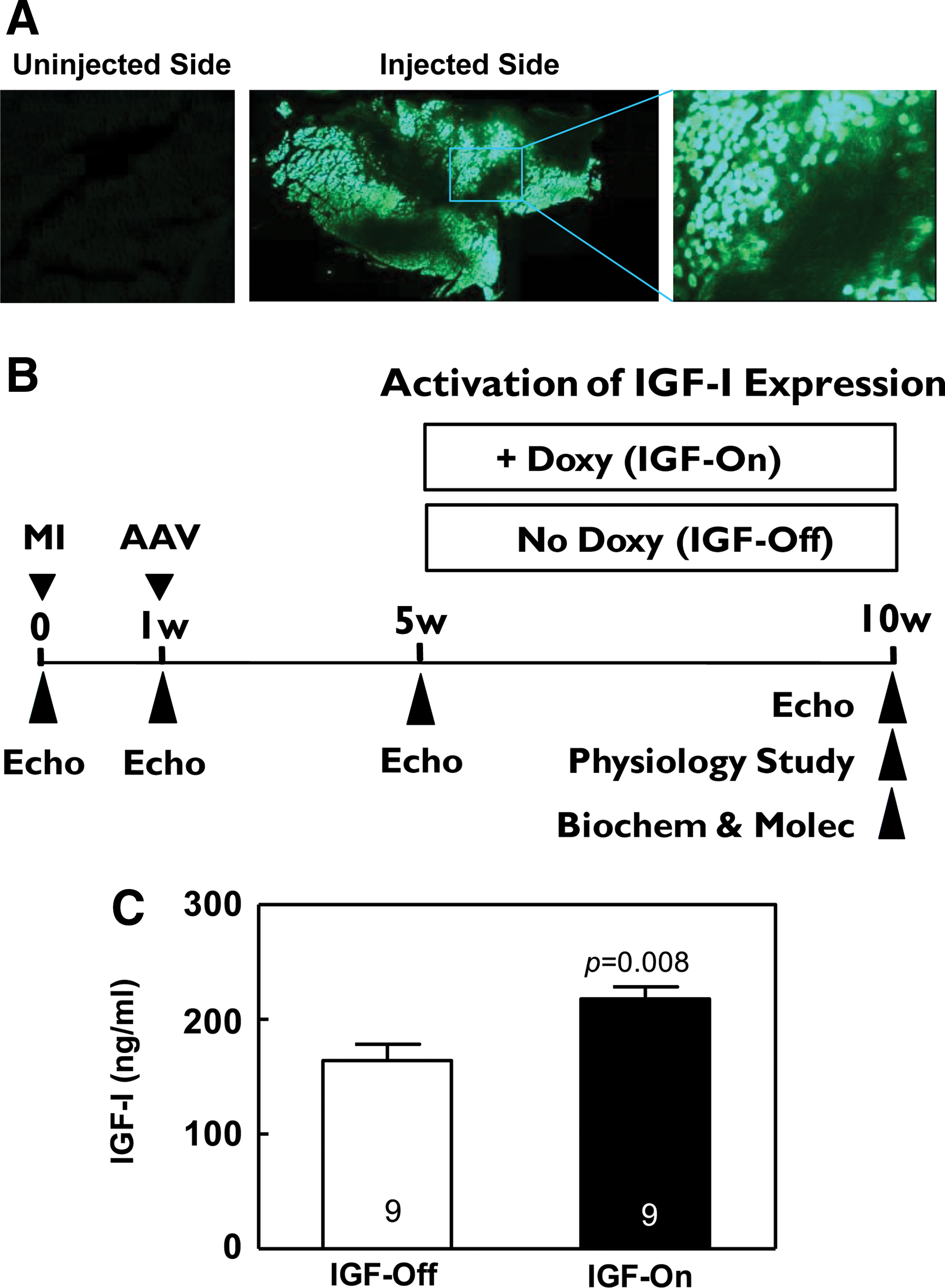
AAV5.IGFI-tet gene transfer in vivo.
Calcium signaling proteins
Viable (noninfarcted) left ventricular (LV) samples underwent Western blotting to assess key proteins involved in Ca2+ handling, including the following: phospholamban (PLN; Affinity BioReagents, Golden, CO), phospho-PLN (Upstate Biotechnologies, Lake Placid, NY), calsequestrin (Novus Biologicals, Littleton, CO), and SERCA2a (Affinity BioReagents), using previously published methods (Tang et al., 2010).
Heart failure model and AAV5 gene transfer
(See Fig. 2B.) Animal use and care were done in accordance with institutional and National Institutes of Health (NIH, Bethesda, MD) guidelines. Proximal coronary occlusion in male Sprague-Dawley rats (200–250 g; Harlan Laboratories, Indianapolis, IN) was used to induce anterior wall myocardial infarction (MI), and subsequent CHF as previously described (Swaney et al., 2007). One week later, survivors with sufficiently large MIs (determined by echocardiography) were selected for gene transfer. AAV5.IGFI-tet (2×1012 GC) was injected into four sites (5×1011 GC in each of four injections) in the unilateral tibialis anterior muscle under light anesthesia (1.5% isoflurane). Four weeks after gene transfer, before activation of IGF-I expression, those with an LV ejection fraction (EF) less than 45% (normal, >75%) were enrolled. Rats were randomly assigned to receive normal drinking water (IGF-Off) or water with doxycycline at 200 mg/liter (IGF-On). Blood samples were collected before and 5 weeks after activation of IGF-I expression. The primary end point of the study was LV function 5 weeks after activation of IGF-I expression. Data were acquired and analyzed without knowledge of group identity.
Echocardiography
Echocardiography was performed in anesthetized rats (1.5% isoflurane) before enrollment, and at 1, 5, and 10 weeks after MI as previously described (Swaney et al., 2007).
Physiological studies
Anesthetized rats (sodium pentobarbital, 100 mg/kg, intraperitoneal) underwent studies with a 2F conductance-micromanometer catheter (SPR838; Millar Instruments, Houston, TX) inserted via the right carotid artery across the aortic valve and into the LV chamber as previously described (Tang et al., 2010). Measurements included the following: heart rate, LV pressure development (LV + dP/dt) and relaxation (LV – dP/dt), mean and phasic arterial blood pressure, and stroke volume. Cardiac output and systemic vascular resistances were then calculated. Dobutamine was infused (5 μg/kg/min, intravenous, 5 min) to measure LV response to β-adrenergic receptor stimulation and to evaluate the reserve function of the heart.
Necropsy and tissue collection
Hearts were collected after diastolic arrest (0.2 M KCl). A short axis midwall LV ring was formalin-fixed and paraffin-embedded. The border zone (the viable LV free wall bordering the infarct) and the remote zone (the interventricular septum) were collected, frozen in liquid nitrogen, and stored at −80°C for quantitative RT-PCR, Western blotting, and caspase-3/7 activity assays. Anterior tibialis muscles were also collected and stored at −80°C for detecting AAV5.IGF-I-tet DNA and IGF-I mRNA.
Cardiac fibrosis and apoptosis
Dewaxed sections (5 μm) were rehydrated, stained with picrosirius red, dehydrated, and mounted in Permount as previous described (Tang et al., 2010). Collagen fractional area in the septum was quantified from picrosirius red-stained sections, using NIH ImageJ software. Cardiac myocyte apoptosis was evaluated by terminal deoxynucleotidyltransferase dUTP nick end labeling (TUNEL) staining as previously described (Tang et al., 2010).
PCR and RT-PCR
AAV5 virus was detected by quantitative real-time PCR (TaqMan) using the following primers: forward, GGCATTATGCCCAGTACATGAC; reverse, CATCACCATGGTAATAGCG; detection probe, tetrachloro-6-carboxyfluorescein-TGGGACTTTCCTACTTGGCAGTACATCTACGTATT. Transgene IGF-I mRNA expression was measured by quantitative RT-PCR using transgene-specific primers (forward, CAAGACTCAGAAGGAAGTACACTGGAA; reverse, AATGTACCGGTAGGTGTCCATTCT; detection probe, tetrachloro-6-carboxyfluorescein-AACACAAGTAGAGGAAGTGAAGGAAACAAGACCTA). Activation of fetal gene program was measured as previously described (Tang et al., 2010).
Serum IGF-I
Free IGF-I in serum was measured with a Duoset IGF-I ELISA kit (R&D Systems, Minneapolis, MN) according to the manufacturer's instructions.
Evaluation of angiogenesis
Capillary density
Platelet endothelial cell adhesion molecule (PECAM; CD31) immunohistochemistry was performed to identify endothelial cells in sections of LV (5 μm), which were deparaffinized, rehydrated, treated with trypsin (0.05%, 10 min) for antigen retrieval, and treated with H2O2 to block endogenous peroxidase activity. After incubation with PECAM antibody (BD Biosciences, San Jose, CA) overnight, LV sections were washed extensively and incubated with horseradish peroxidase-conjugated anti-mouse IgG (Dako North America, Carpinteria, CA). Stable 3,3′-diaminobenzidine (DAB) solution (Invitrogen, Carlsbad, CA) was then added to LV sections to visualize PECAM staining. Capillary density was calculated as the number of capillaries per square millimeter of viable LV wall.
Genome-wide gene expression profiling
Microarray analysis was performed on Rat Whole Genome OneArray (version ROA1.1; Phalanx Biotech Group, Belmont, CA), using purified RNA samples (RNA integrity number, >7.5) from viable LV, skeletal muscle, and liver. The Phalanx Rat OneArray ROA1.1 covers 24,358 rat transcript probes corresponding to 19,565 rat genes and 980 experimental control probes. High hybridization performance was achieved through isothermal ∼60-mer sense-strand probe design (
Statistical analysis
Results are shown as means±SE. Differences in group means were tested by Student t test (two-tailed) or two-way analysis of variance with Bonferroni t testing. The null hypothesis was rejected when p<0.05.
Results
Regulated IGF-I expression in cultured cardiac myocytes
(See Fig. 1B–D.) To evaluate the time course of regulated IGF-I expression, cultured neonatal rat cardiac myocytes underwent gene transfer with AAV5.IGFI-tet and activation with doxycycline. Expression of IGF-I mRNA was increased (vs. unstimulated) by 1.5-fold within 30 min, and reached a peak of 14-fold elevation by 24 hr. At 48 hr, IGF-I mRNA was somewhat less (10-fold), reflecting doxycycline degradation. To turn off IGF-I expression doxycycline was removed, using four sequential PBS washes (“off-wash”). IGF-I mRNA rapidly decreased after doxycycline withdrawal (Fig. 1B). AAV5.IGFI-tet gene transfer in cultured cardiac myocytes provided doxycycline-dependent IGF-I protein expression in the medium (Fig. 1C), which was associated with increased phosphorylation of Akt (Fig. 1D), a known target for IGF-I in cardiac myocytes.
Transgene expression after AAV5-mediated gene transfer in vivo
Three weeks after intramuscular injection of AAV5.EGFP, we found extensive EGFP expression in anterior tibialis muscle in the injected side, but not in the uninjected side (Fig. 2A). We next performed intramuscular AAV5.IGFI-tet injection in rats after MI (Fig. 2B), and 9 weeks later confirmed the substantial presence of AAV5.IGFI-tet DNA (8.2±1.6×104 copies/μg genomic DNA, n=6) in the injected anterior tibialis, which also showed marked IGF-I mRNA expression after 5 weeks of doxycycline administration (qRT-PCR: 8.0±1.5×104 copies/μg total RNA, n=6). Last, and most importantly, increased free IGF-I in serum was documented 9 weeks after gene transfer (IGF-Off, 164±14 ng/ml; IGF-On, 217±11 ng/ml; n=9, both groups; p=0.008; Fig. 2C). IGF serum levels in IGF-Off rats were not different from those of normal rats that had not received gene transfer (normal rats, 151±8 ng/ml, n=11; IGF-Off, 164±14 ng/ml, n=9; p=0.45), confirming minimal doxycycline-independent transgene expression.
Echocardiography and physiological studies
(See Tables 1 and 2.) Five weeks after MI, rats showed reduced LV ejection fraction (IGF-Off, pre-MI: 78±3%; 5 weeks post-MI: 41±3%, n=9; p<0.001; IGF-On, pre-MI: 79±2%; 5 weeks post-MI: 37±4%, n=10; p<0.001), and severely reduced basal LV + dP/dt (normal LV + dP/dt, 10,853±917 mmHg/sec, n=4). Activation of IGF-I expression was associated with a 9% absolute and 22% relative increase in LV ejection fraction (p=0.02; Table 1). The IGF-Off group, by contrast, showed a 5% relative decline in LV ejection fraction over the same period. Physiological assessment showed substantial increases in LV contractile function (LV + dP/dt) after activation of IGF-I expression (p<0.0001; Table 2). Basal LV + dP/dt was increased 50%, and dobutamine-stimulated LV + dP/dt was increased 81% (p<0.0001), indicating that activation of IGF-I expression improves LV contractility in failing rat hearts. Furthermore, activation of IGF-I increased cardiac relaxation as reflected by increased rate of LV pressure decay (p=0.03; Table 2). Last, activation of IGF-I resulted in substantial increases in systolic arterial pressure (p=0.0003), cardiac output (p=0.02), and stroke work (p=0.0001) both before and during dobutamine infusion, in the absence of changes in systemic vascular resistance (p=0.49), confirming a direct cardiac effect.
HR, heart rate; EDD, end-diastolic diameter; ESD, end-systolic diameter; LVEF, left ventricular ejections fraction.
Values represent means±SE; p values were determined by Student t test, comparing changes for each variable between groups (two-tailed, unpaired).
HR, heart rate; MAP, mean arterial pressure; LV, left ventricular; SVR, systemic vascular resistance in Wood units (MAP/CO; mmHg·sec/ml).
Data denote means±SE. Group size as indicated except (1) n=9 (both groups) for CO and SVR; and (2) body weight: n=7 for IGF-Off. Probability values, from two-way ANOVA, showing IGF-I effect.
LV mass, apoptosis, and fibrosis
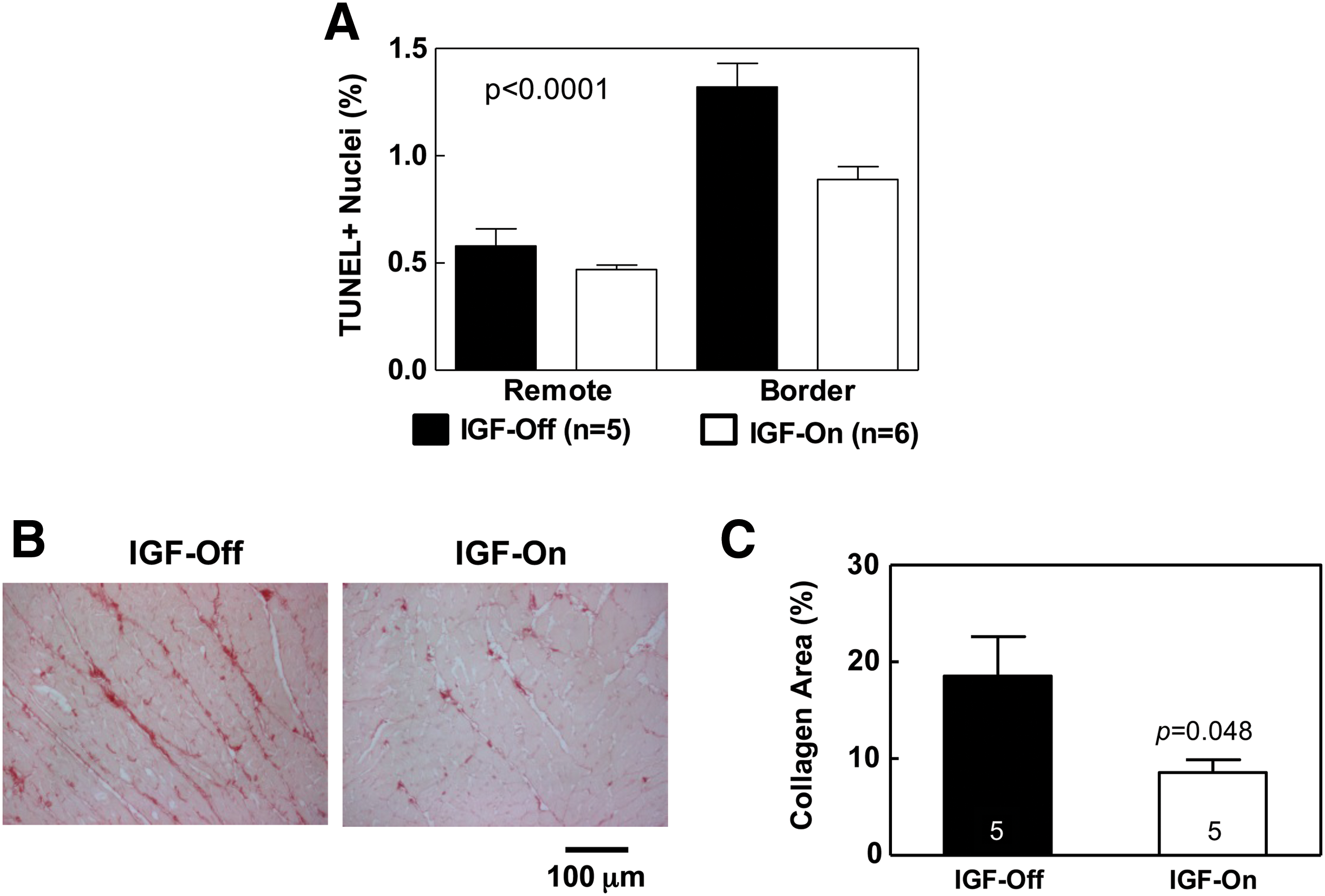
There were no group differences in LV weight (IGF-Off: 0.98±0.03 g, n=7; IGF-On: 1.02±0.04 g, n=10), body weight (IGF-Off: 419±15 g, n=7; IGF-On: 425±8 g, n=10), or LV-to-body weight ratio (IGF-Off: 2.43±0.03 g/kg, n=7; IGF-On: 2.31±0.06 g/kg, n=10). Activation of IGF-I expression was associated with reductions in cardiac myocyte apoptosis (p<0.0001; Fig. 3A), and caspase-3/7 activity in the border region, but Bcl2 expression was unchanged. Activation of IGF-I expression reduced LV fibrosis—a 54% reduction of collagen fractional area was observed (Fig. 3B and C).
Calcium signaling proteins
No group differences were seen in LV protein content of PLN, phospho-PLN, calsequestrin, or SERCA2a.
Evaluation of angiogenesis
Capillary density. PECAM (CD31) immunohistochemical studies performed on viable transmural LV samples showed no difference in capillary density 5 weeks after activation of IGF-I expression (IGF-Off: 867±154 capillaries/mm2, n=5; IGF-On: 950±177 capillaries/mm2, n=5; p=0.73).
Genome-wide gene expression profiling
Microarray analysis was performed to screen the effects of activation of IGF-I expression on expression of angiogenesis-related genes. Using purified RNA samples from viable LV, skeletal muscle, and liver, we found that <1.1% of the 24,358 transcripts covered by Rat OneArray showed >2-fold change (an a priori criteria for statistical significance) after activation of IGF-I expression. No change in mRNA expression of the following angiogenesis-related genes was found: vascular endothelial growth factors a–c (VEGFa, VEGFb, and VEGFc), PECAM, angiopoietin-1, angiopoietin-2, and platelet-derived growth factors a–d (PDGFa, PDGFb, PDGFc, and PDGFd). We also confirmed these results by quantitative real-time RT-PCR.
Discussion
The key finding of this study was that regulated expression of IGF-I via gene transfer in skeletal muscle increases function of the failing heart. Although a number of transgenes have been associated with increased LV function, novel features underscore the importance of our findings. First, gene transfer was performed by a simple intramuscular injection, circumventing impediments imposed by cardiac-targeted approaches, and enabling easier translation to clinical application. Second, the studies were conducted in a manner replicating those present in clinical settings: Heart failure was present at the time of transgene activation. Last, transgene expression was regulated, enabling the turning on and off of expression as needed, a desirable safety feature for clinical application.
The evidence for increased LV function came from multiple sources: increased ejection fraction by echocardiography, increased rate of LV pressure development and decay, and increased maximal systolic blood pressure, data that were collected in blinded studies. A potential problem vis-à-vis data interpretation derives from the potential vasodilator effects of IGF-I, which would be predicted to enhance LV function indirectly. However, there was no group difference in systemic vascular resistance after activation of IGF-I expression, despite striking increases in basal and dobutamine-stimulated cardiac output, stroke work, blood pressure, and LV + dP/dt. These data establish a direct cardiac effect of IGF-I gene expression on cardiac contractile function of the failing heart, independent of systemic vascular resistance. It is important to emphasize that these favorable effects of activation of IGF-I expression were evident in the basal (unstimulated) state, but also in response to β-adrenergic receptor stimulation with dobutamine, indicating the heart's contractile reserve had also substantially increased after therapy.
These beneficial effects on function of the failing heart were associated with sustained increased free IGF-I in serum after a one-time injection of AAV5.IGFI-tet in the tibialis anterior muscle. Additional benefits included reduced apoptosis in the periinfarct region, and decreased cardiac fibrosis. A previous study showed sustained elevation in serum growth hormone after skeletal muscle delivery of an AAV vector under rapamycin regulation (Rivera et al., 1999). However, the present study went further, by demonstrating that paracrine-based gene transfer can effectively treat CHF.
Although studies of IGF-I gene transfer for the treatment of CHF are rare, IGF-I protein delivery before myocardial ischemia and reperfusion in rats decreased creatine kinase release and reduced apoptosis (Buerke et al., 1995), and combined IGF-I and growth hormone peptides increased cardiac contractility (Duerr et al., 1996). Although there is a paucity of placebo-controlled clinical studies, the acute hemodynamic effects of 4-hr infusions of IGF-I protein in human subjects with CHF (n=8), showed increased cardiac output, decreased vascular resistance, and reduced right atrial and wedge pressures (Donath et al., 1998). However, chronic administration of IGF-I protein has not been evaluated in patients with CHF.
IGF-I protein administration to rats appears to be associated with LV hypertrophy and increased heart function after MI (Duerr et al., 1995). In cultured cardiac myocytes, IGF-I protein increases SERCA2a protein content and sarcoplasmic reticulum (SR) calcium uptake (Kim et al., 2008), which would be anticipated to increase heart function. In the present study we found that IGF-I gene transfer was associated with substantial increases in LV function and cardiac reserve, in the absence of LV hypertrophy or changes in SERCA2a protein expression. We did not assess SR calcium uptake, but found no group differences in LV contents of PLN, phospho-PLN, calsequestrin, or SERCA2a. IGF-I protein has been shown to have beneficial effects on heart function by increasing myofilament sensitivity to Ca2+ in isolated isovolumic aequorin-loaded hearts and in ferret papillary muscles (Cittadini et al., 1998). Whether such a mechanism explains increased function of failing hearts seen in the present study is not known and is a focus of ongoing studies.
In isolated neonatal rat cardiac myocytes, doxycycline activation of IGF-I expression was associated with increased Akt phosphorylation (Fig. 1D). However, we found no group differences in phospho-Akt and total Akt protein levels in LV samples from IGF-Off and IGF-On rats 5 weeks after activation of IGF-I expression (data not shown). These findings suggest that IGF-I expression is not associated with prolonged Akt activation, perhaps reflecting a negative feedback of Akt activation on IGF-I receptor expression (Glass, 2010). Because prolonged Akt activation may be deleterious to the heart (Shiojima et al., 2005), shorter term Akt activation may be preferred.
IGF-I has been reported to possess mild angiogenic effects in some settings, but capillary density in noninfarcted areas of the LV showed no group differences in capillary number or in expression of angiogenic genes. We conclude that the mechanism underlying improved LV function associated with activation of IGF-I expression was unrelated to angiogenesis in this model.
Clinical epidemiological studies report a correlation between increases in serum IGF-I concentration (2-fold elevations) and prostate and premenopausal breast cancer. Although a direct causal effect of IGF-I on these tumors has not been demonstrated clinically, it seems prudent that IGF-I expression should be regulated if used, a goal we achieved by Tet regulation. The high mortality of human subjects with CHF (50% mortality in 4–5 years) lessens the likelihood that IGF-I gene therapy would be used for decades, which would reduce cancer risk. Moreover, the modest increase in serum IGF-I seen in the present study (38% increase), although adequate to increase function of the failing heart, may not be sufficiently high to increase cancer risk. Even so, it will be important to balance risk and benefit if these preclinical data are translated to clinical practice.
Limitations of the study: Translation of these results to clinical trials will require overcoming immune response to vector and to bacterial and virus components of the Tet regulation system. Intramuscular delivery, which works well in rodents, is beset with difficulties in nonhuman primates. In that regard, we have shown that intravenous delivery of AAV5.IGFI provides high levels of free IGF-I in serum. Identification of an AAV serotype best suited for intravenous delivery but with little preexisting immunity in humans will be essential. Unaddressed in the present study were (1) whether the Tet regulation system will be effective after serial bouts of doxycycline activation; and (2) whether IGF-I expression was associated with anti-IGF-I antibodies, although because serum levels of IGF-I were sustained 9 weeks after vector delivery, this seems unlikely.
In conclusion, activation of IGF-I expression after a one-time skeletal muscle injection of AAV5.IGFI-tet improved function of the failing heart. This was associated with increased circulating free IGF-I levels and reduced cardiac apoptosis and fibrosis. These results provide a rationale to conduct additional studies in larger animals, to explore alternative AAV serotypes, other regulated expression systems, and alternative routes of delivery to translate these findings to clinical application.
Footnotes
Acknowledgments
The authors thank Matthew Spellman for technical assistance. This work was supported by grants from the NIH (5P01HL066941, HL081741, HL088426); by a Merit grant from the Department of Veterans Affairs; and by American Heart Association Beginning Grant-In-Aid Awards 0765064Y and 0865147F, and Grant-In-Aid Award 11GRNT7610059.
Author Disclosure Statement
There are no financial conflicts of interest to disclose.