
Abstract
MicroRNAs (miRNAs) compose a relatively new discipline in biomedical research, and many physiological processes in disease have been associated with changes in miRNA expression. Several studies report that miRNAs participate in biological processes such as the control of secondary injury in several disease models. Recently, we identified novel miRNAs that were abnormally up-regulated in a traumatic spinal cord injury (SCI). In the current study, we focused on miR20a, which causes continuing motor neuron degeneration when overexpressed in SCI lesions. Blocking miR20a in SCI animals led to neural cell survival and eventual neurogenesis with rescued expression of the key target gene, neurogenin 1 (Ngn1). Infusion of siNgn1 resulted in functional deficit in the hindlimbs caused by aggressive secondary injury and actively enhanced the inflammation involved in secondary injury progression. The events involving miR20a underlie motor neuron and myelin destruction and pathophysiology and ultimately block regeneration in injured spinal cords. Inhibition of miR20a expression effectively induced definitive motor neuron survival and neurogenesis, and SCI animals showed improved functional deficit. In this study, we showed that abnormal expression of miR20a induces secondary injury, which suggests that miR20a could be a potential target for therapeutic intervention following SCI.
Introduction
The understanding of miRNA has expanded rapidly, but controlling miRNA levels as a therapeutic strategy remains a challenge. The identification of specific genes would further our understanding of the mechanisms underlying secondary injury progression and tissue degeneration linked to various diseases. Based on a specific miRNA's target gene information, RNA-based technologies have potential as a therapeutic strategy. Antisense oligonucleotides targeting a mature miRNA sequence, or miRNA inhibitors, including cholesterol-conjugated 2′-O-methyl oligonucleotide (antagomir), locked-nucleotide modified oligonucleotide, and 2′-O-methoxyethyl-phosphorothioate, can significantly lower the tissue content of disease-promoting miRNAs. Thus, miRNA mimics can serve to evaluate the level of miRNAs (Martinez et al., 2002; Krutzfeldt et al., 2005; Jackson et al., 2006). Aberrant miRNA expression might become a novel therapeutic target for the treatment of various diseases that involve miRNA, including cardiovascular disease and traumatic injury.
When tissue and/or organs are lost as a result of either infection or trauma, a series of regenerative stages are activated. First, healing of the epithelium is followed by generation of precursor cells for replacement of tissue-specific cells. Next, precursor cells differentiate and migrate to loci of cell loss, which involves numerous interacting signaling pathways that integrate and coordinate complicated transcription and post-translational pathways of specific and appropriate target genes. Recent research has reported (Beck et al., 2006) that miRNAs are essential to control both cellular proliferation and differentiation. Application of miRNAs to regenerative scenarios has been applied in numerous species, for example in zebrafish (Thatcher et al., 2008) and in humans in which 20%–30% of the genome is regulated by interaction with miRNAs (Krichevsky, 2007). A number of miRNAs have been reported in the mammalian nervous system in relation to disorders of neural lineage and development (Bak et al., 2008; Kosik, 2006; Miska et al., 2004), neurodegenerative diseases (Bilen et al., 2006; Kim et al., 2007; Schaefer et al., 2007), and fragile X syndrome (Bak et al., 2008).
The spinal cord of adult mammals has a limited capacity for regeneration following traumatic injury, and many studies have provided valuable information related to the molecular and cellular mechanisms and consequences of secondary injuries, including pathology (Xu et al., 2001; Bilen et al., 2006; Kang et al., 2006). Recent study of miRNAs has introduced the concept of a novel type of regulatory control over gene expression during development and pathological progression. Several miRNAs exhibit expression patterns that are specific to a tissue or to injury development, and some have also been reported to be associated with specific diseases such as cancer, cardiovascular disease, and chronic hepatitis C virus infection (Lu et al., 2005; von Rooji et al., 2006; Zhao et al., 2007). Therapeutic trials aimed at silencing miRNAs in vivo have been conducted (Lu et al., 2005; Marcucci et al., 2008). Several studies have reported that the proneural basic helix-loop-helix transcription factors neurogenin 1 (Ngn1) and Ngn2 contribute to neurogenesis in the hippocampus and olfactory receptor cells during embryonic development (Morrison, 2001; Bertrand et al., 2002; Wu et al., 2003; Berninger et al., 2007). On the other hand, the proneural gene Ngn1 gives rise to daughter cells that undergo terminal differentiation into postmitotic Ncam-expressing olfactory receptor cells (Wu et al. 2003). Ngn1 is also involved in cell specification during central nervous system development (E.J. Kim et al., 2011; Quiñones et al., 2010).
In this study, we report novel findings about miR20a in lesions from injured spinal cord tissue in mice. We also identify the roles of miR20a in traumatic injury inducing pathophysiology and neuroprotection within injured spinal cord by altering miR20a expression. Our microarray-mediated miRNA profiling enabled identification of highly conserved miRNAs that have significantly different expression at various time points after spinal cord injury (SCI) in animals. Among these, miR20a, which was overexpressed in injured spinal cord lesions, was further identified as a main factor in apoptotic cell death and inhibition of various regenerative activities following traumatic SCI.
Material and Methods
miRNA microarray and data analysis
Recently, we analyzed miRNAs that had differential expression in lesions of injured spinal cord tissue. miRNA assay probes corresponding to annotated mouse miRNA sequences (miRBase) were used to identify 265 differentially expressed miRNAs. For assay specificity, all probes were compared to avoid sequence similarity between probes. Before hybridization to the miRNA assay product, the identity of each bead was detected, and identical arrays were used to report the results of similar assays (GoldenGate Genotyping Assay, DASL Gene Expression Assay, GoldenGate Methylation Assay). Arrays were scanned and automatic image registration and intensity extraction software was used to determine intensity for each miRNA bead type. In this study, all microarray data were described following Minimal Information About A Microarray Experiment (MIAME) guidelines (Chen et al., 2008).
Induction of SCI and inhibitor administration
Adult female ICR mice weighing 30 g (∼6 weeks of age) were housed in a controlled environment and provided with standard animal chow and water. Animal care was carried out in compliance with Korean regulations regarding the protection of animals used for experimental and other scientific purposes. The animals were subjected to traumatic SCI via a modified version of the protocol described by Kang et al. (2006). Briefly, rongeurs were used to remove the neural spines of the T9 and T10 thoracic vertebrae and to carry out a laminectomy to expose the outer surface of the spinal dura mater. The dura was opened using microscissors attached to a stereotactic apparatus, and the tip of the scissors were inserted 3.5 mm below the dorsal surface of the cord. The microscissors were then closed twice to provide a consistent total transection of the dorsal and ventral corticospinal tracts and was confirmed by experiment. The mice were then randomly assigned into two groups: 30 SCI mice treated with miR20a inhibitor and 10 SCI mice that received scrambled RNA (controls). For inhibitor administration, RNA oligomer was injected directly into the lesion site 2 days after injury. The control (vehicle) group received the same concentration of scrambled RNA. A total of 40 animals were utilized in these experiments.
Spinal cord–derived neural progenitor cell culture and neurogenesis induction
Spinal cord-derived neural progenitor cells (NPCs) were isolated from 5- to 6-week-old, adult ICR mice. The adult mouse spinal cord tissues were placed in 4°C Hank's Balanced Salt Solution (HBSS) without Ca2+ or Mg2+ and washed extensively with equal volumes of HBSS. The tissues were then minced into small pieces (1 mm3) and enzyme digested at 37°C for 30 min with 0.025% trypsin. Enzyme activity was neutralized with Dulbecco's modified Eagle's medium (DMEM), containing 10% fetal bovine serum (FBS) and then centrifuged (425×g for 5 min) to obtain a high density of cells. The cultured NPC pellet obtained by centrifugation was incubated overnight at 37°C in 5% CO2 in DMEM containing 10% FBS. Culture media was changed at 48 hr of culture and then every fourth day thereafter. After passage 2 or 3, we evaluated differentiation potency of cultured cells. To evaluate the neural lineage differentiation potency of cultured NPCs, cells were placed in differentiation medium. Cultured NPCs formed spherical clumps of cells in suspension culture media and free-floating neurospheres in neural induction culture media. The neurospheres were then transferred to a petri dish and cultured in neurobasal medium (Invitrogen) supplemented with B27 (Invitrogen), 20 ng/mL basic fibroblast growth factor (bFGF), and 20 ng/mL brain-derived neurotrophic factor (BDNF) (Sigma) for 3 days. The culture density of the spheroid bodies was maintained at 10–20 cells/cm2 to prevent self-aggregation. For neuronal differentiation induction, neurospheres derived from NPCs were plated on a Poly-D-Lysine (PDL)-laminin double-coated cover slip. During differentiation, 60% of the media was replaced every 4 days. All data shown are representative of at least three independent experiments. To detect differentiated neuronal cells, we performed immunocytochemical analysis using neuronal marker primary antibodies.
Luciferase assay
The oligonucleotides Ngn1-Fw-wt and Ngn1-Rev-wt were ligated to form a fragment containing the miR20a binding site on the Ngn1 3′ untranslated region (UTR) predicted by TargetScan (
Whole mount in situ analysis
In situ hybridization of miR20a and Ngn1 was performed on injured or normal spinal cord tissue. Several kinds of gene oligonucleotide probes were synthesized (Dharmacon RNA Technologies). Tissue slides were air-dried and prepared for 15 min in saline citrate (pH 7.0) containing 0.1 mg/mL proteinase K (Sigma Aldrich) at room temperature. Sections were washed in 1 × sodium chloride–sodium citrate buffer (SSC) for 5 to 10 min. Labeled oligonucleotides were added (30 pM) to the hybridization buffer solution, and the slides were covered with Parafilm™, incubated in a humid chamber, and finally air-dried. Controls in which specific probes were omitted resulted in no positive staining signal. Finally, we detected the specimens using a Leica fluorescence microscope (Leica Microsystems). These studies were repeated at least three times (Im et al., in press).
Inhibition assays
To confirm the function of the Jak2 signaling pathways during proliferation, NPCs were seeded in 10-cm dishes at a density of 6×105 and cultured in DMEM containing 2% FBS for 8 hr at 37°C in a CO2 incubator. NPCs were exposed to the Jak2 inhibitor AG490 (20 μM; Biomol) or left untreated. The self-renewal activity of cells was analyzed via cell counting and Western blotting.
Small interfering RNA silencing
To knock-down miR20a and Ngn1, a synthesized small interfering RNA (siRNA) duplex was obtained from Dharmacon RNA Technologies. For transient transfection, about 60% confluent cells in six wells were transfected with 10 μM siRNA using Lipofectamine (Invitrogen). Cells were allowed to stabilize for 48 hr before being used in experiments. Cells were harvested after 24 hr for RNA isolation. Silencer Negative Control siRNA (catalog number 4611; Ambion, Inc.) was utilized as a control for nonspecific gene silencing. The transfection of siRNAs was conducted using DharmaFECT siRNA transfection reagents, in accordance with the manufacturer's instructions (Dharmacon RNA Technologies). Two complementary hairpin siRNA template oligonucleotides harboring the 21-nt target sequences of mouse Ngn1 were used for transient transfection of 50 nM siRNA. For that, three separate Ngn1 siRNAs (Silencer® predesigned siRNAs; Ambion) and scrambled siRNAs with the same nucleotide content were assessed (Jang et al., 2012).
For knock-down of miR20a in vivo, mice were anesthetized and immobilized with a stereotaxic instrument. Experimental trials were first conducted to verify consistent, total transection of the corticospinal tracts on both sides (Jee et al., 2010). One day after the operation, antisense miR20a (20 μM stock solution) was diluted with DharmaFECT (Dharmacon RNA Technologies.), and a total of 5 μL (2.5 μL of DharmaFECT and 2.5 μL of antisense miR20a or scrambled RNA) was injected with a 25-μL Hamilton syringe 2 days after SCI. Each mouse was anesthetized with avertin, its dorsal spinal cord was opened with a surgical knife, and the vertebral bone was removed with surgical scissors and rongeurs. Antisense miR20a was injected at a final concentration of 10 μM.
ATP measurement
Total concentration of protein was determined using the Protein Assay Kit (Bio-Rad) following the manufacturer's instructions. Cells were resuspended in buffer containing 150 mmol/L KCl and 0.1% (w/v) bovine serum albumin at a concentration of 1 mg protein/mL of buffer. ATP synthesis was initiated by the addition of 250 μL of the cell suspension to ADP-containing substrate buffer. Cells were incubated at 37°C for 10 min. The quantity of ATP was measured in a luminometer (Berthold Detection Systems) with the ATP Bioluminescence Assay Kit (Roche Diagnostics) following the manufacturer's instructions (Shepherd et al., 2006).
Bisulfite modification and sequencing of genomic DNA
For bisulfite DNA sequencing, bisulfite conversion was conducted using the EZ DNA Methylation–Gold kit (Zymo Research) following the manufacturer's instructions. The bisulfite-modified DNA was then immediately utilized for polymerase chain reaction (PCR) or stored at or below −20°C. The converted DNA was amplified via PCR or designed with MethPrimer (
CHIP analysis
The monoclonal antibodies anti-Ngn1 and pSTAT3 were obtained from Santa Cruz Biotechnology and rabbit IgG (PP64B) antibody was obtained from Upstate. Cells were harvested and chemically crosslinked with 1% formaldehyde (Sigma), and fixation was quenched by 2.5 M glycine at room temperature. Washed cell pellets were resuspended in pre-IP dilution buffer containing 4% IGEPAL and 1 mM phenylmethanesulfonyl fluoride (PMSF). Cells were sonicated and fragments ranged in size from 200 to 1000 bp. Sonicated cell extracts equivalent to 2×106 cells were used in subsequent immunoprecipitations. Samples were pre-cleared with protein G Dynabeads (Dynal) in 1000 mL of dilution buffer and 5 mL of Upstate protease inhibitor cocktail II. Cell extracts were incubated with 1 mg of antibody overnight (B.S. Kim et al., 2011).
Electrophysiological recording
Electrophysiological evaluation of injured spinal cord tissue (treated with simiR20a or scrambled RNA) was performed before, immediately after, and 3 weeks after sciatic axotomy. For electrophysiological recording, under anesthesia, the mouse's left sciatic nerve and fourth digital nerve were exposed. The evoked action potential in responding to the stimuli (1 msec, 500 mV) in the ipsilateral sciatic nerve was recorded using Powerlab-800 system (ADInstruments,
Statistical analysis
Data were described as the mean±SEM from five or more independent evaluations. Statistically significance during functional recovery among different groups was measured by repeated evaluation of the ANOVA test.
Results
Differential expression of miR20a in SCI lesions
We identified differentially expressed miRNAs and mRNA at a various time points (0, SCI 24 hr, 48 hr, 72 hr, 1 week) via DNA microarray and in situ analysis (Fig. 1A). From among these, we selected several miRNAs targeting specific neuronal, functional protein, and up-regulation factors. Finally, we focused on miR20a in SCI for further study. As shown in Fig. 1, miR20a was prominently elevated in mouse SCI lesions compared with normal spinal cord (Fig. 1B). Moreover, to evaluate whether miR20a can directly regulate Ngn1 expression via binding to its 3′UTR, wild-type and mutated binding sites of miR20a on the 3′UTR region of the Ngn1 gene were cloned into a luciferase reporter plasmid. The cotransfection of pmiR20a-Ngn1-wt with pcDNA-miR20a resulted in decreased luciferase activity (Fig. 1C); miR20a was not able to bind to the mutant construct. In contrast, the transfection of anti-miR20a resulted in restored luciferase activity to control levels (Fig. 1C). Additionally, most of the target genes of miR20a were differentially expressed in SCI lesions, including Ngn1, Ngn2, Gab3, Pax6, SMAD1, DCX, Tuj, FGF8, and SALL1, which were prominently down-regulated at a various time intervals after traumatic injury (Fig. 1D). Moreover, our immunohistochemical analysis of injured spinal cord tissue against GFAP, Nestin, and NF160 showed that miR20a expression was restricted in Nestin-positive NPC and NF160-positive motor neurons at the subcellular level (Fig. 1E).
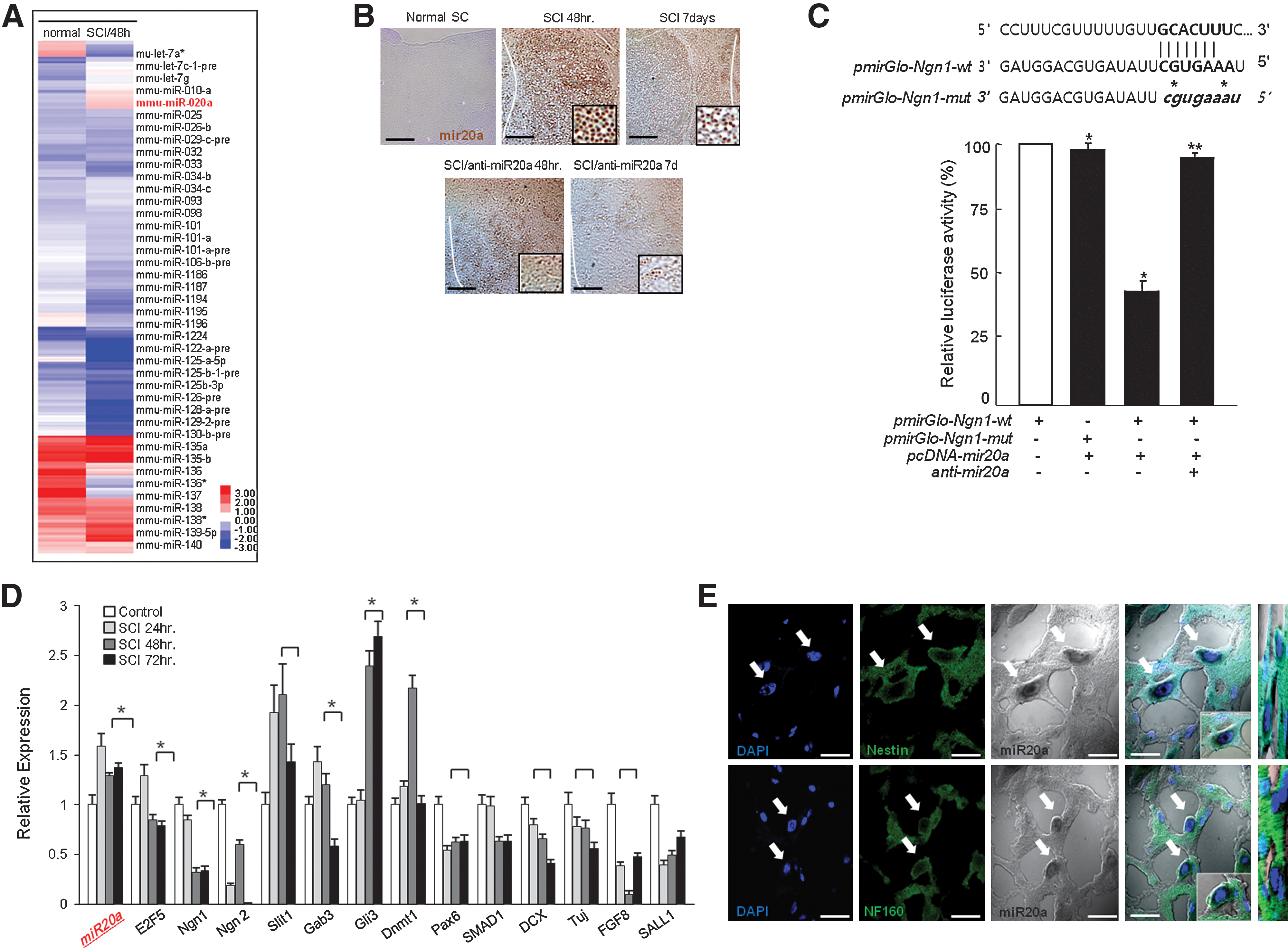
Prominent miR20a expression and down-regulation of target gene expression in traumatic spinal cord injury (SCI).
Major role of miR20a and inhibition of miR20a rescues secondary injury of spinal cord along with functional recovery
In the study of the function of miR20a in SCI lesions, we induced SCI through miR20a infusion in normal spinal cord (Fig. 2A). We directly confirmed miR20a induced apoptotic neural cell death in spinal cord tissue after 2 days of miR20a infusion along with aggressive inflammation (Fig. 2A; Supplementary Fig. S1; Supplementary Data are available online at
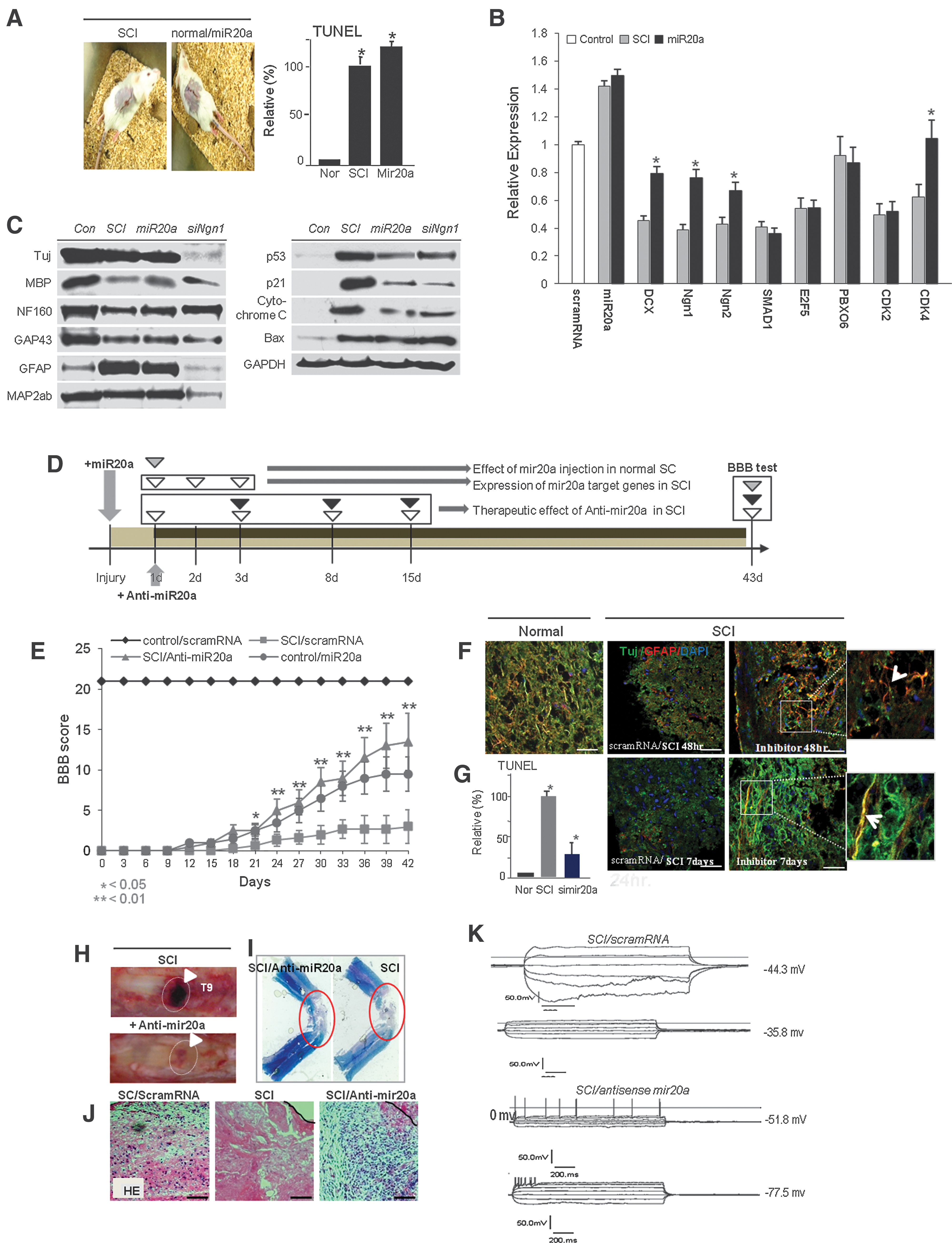
Therapeutic application and evaluation of traumatic SCI by effective inhibition of miR20a expression.
Knock-down of miR20a expression effectively controls neuroprotection of motor neurons and myelin and regeneration in vitro and in vivo
Inhibition of miR20a expression resulted in recovery of major target gene expression 48 hr and 7 days after infusion of simiR20a (Fig. 3A), which prominently protected apoptotic neuronal cell death by caspase 3 activation and ATP down-regulation in SCI animals (Fig. 3B). At the protein level, expression of most of the neuronal markers, including Tuj, MAP2ab, NF160, MBP, and GFAP, and the neuronal regeneration marker GAP43 were prominently up-regulated 48 hr and 7 days after miR20a inhibition in SCI (Fig. 3C; Supplementary Fig. S2). Inhibition of miR20a expression resulted in neuronal cell survival and also down-regulated apoptotic cell death signals such as Bax and cytochrome C expression along with increased Bcl2 after 48 hr and 7 days of simiR20a infusion in SCI (Fig. 3D; Supplementary Fig. S3). As shown in Fig. 3E, in cultured NPCs, TuJ, GFAP, MAP2ab, NF160, and expression of related transcription factors were recovered after miR20a/simiR20a treatment at the protein level. We also confirmed that antisense miR20a infusion resulted in reactivation of STAT3/Jak2/ERK1/2 and Oct4 and Sox2 expression along with increased PI3K/Akt phosphorylation (Fig. 3E). miR20a introduced into NPCs attenuated self-renewal activity, which was rescued by inhibition of miR20a expression (Fig. 3E, F, and G). Along with that, Telomerase reverse transcriptase (TERT) activity and telomere length was more or less decreased, but miR20a/simiR20a (rescue) treatment resulted in rescued TERT activity and elongated telomere length (Fig. 3H, I). Compared to control or miR20a-treated cells, miR20a/simiR20a-treated NPCs showed that nuclear localization and DNA binding activity of Ngn1 protein was induced in the promoter regions of Oct4, STAT3, and GAP43 genes, with an increase in transcriptional expression (Fig. 3J; Supplementary Fig. S4). Among Ngn1-regulated genes, the regulatory region of Oct4 was significantly demethylated after miR20a/simiR20a treatment in NPCs compared to NPCs treated with scrambled RNA or miR20a (Fig. 3K). Figure 3L shows the functional role of miR20a in injured spinal cord for neuronal cell survival and regeneration (Fig. 3L).

Inhibition of miR20a expression effectively induces neuroprotection and regeneration in traumatic SCI.
Critical role of Ngn1 for neuroprotection in traumatic SCI lesion
As one of the key target genes of miR20a, Ngn1 has a critical role in maintenance of cell survival, self-renewal, and neurogenesis in normal and injured spinal cords. Following our gene expression analysis, we found that siNgn1 infusion in normal mouse spinal cord induced SCI with very similar behavior to the traumatic SCI model and showed completely paralysis after 3 day of siNgn1 infusion in the spinal cord (Fig. 4A). In contrast, the HBSS-infused control animal had normal behavior, including regular coordinated walking. When we detected Ngn1 expression by in situ hybridization injured spinal cord, it was dramatically depleted but normal, and SCI animal tissue in which miR20a was inhibited showed overexpression of nuclear Ngn1 (Fig. 4B). In vivo injured spinal cord microenvironment also demonstrated that knock-down of Ngn1 expression effectively down-regulated Ngn1, Ngn2, DCX, Tuj, SMAD, Nestin, and MAP2ab expression after 48 hr of SCI induction (Fig. 4C and D). Coordinatively, siNgn1 effectively inhibited regenerative marker GAP43; neuronal markers TuJ, GFAP, MBP, and MAP2ab; and NF160 protein expression (Fig. 4D). Inhibition of Ngn1 by siNgn1 infusion in normal spinal cord effectively enhanced cytotoxic effects of motor neurons and myelin and also constituted a pathologic microenvironment with increased levels of interleukin (IL)-6, tumor necrosis factor (TNF)α, IL-1β, and COX2 expression in normal spinal cord (Fig. 4E). ED1-positive microglia cells were effectively increased after infusion of siNgn1 expression (Fig. 4E–G). We confirmed that inhibition of Ngn1 was associated with an induction of caspase 3 activation and iNOS, eNOS, and COX2 overexpression that effectively enhanced motor neuron and myelin degeneration following traditional secondary injury activation (Fig. 4G and H). Expression of direct cell death–inducing factors, Bax and cytochrome C, was highly up-regulated along with increased caspase 3 activity in miR20a- and siNgn1-infused spinal cord tissue (Fig. 4G, H).
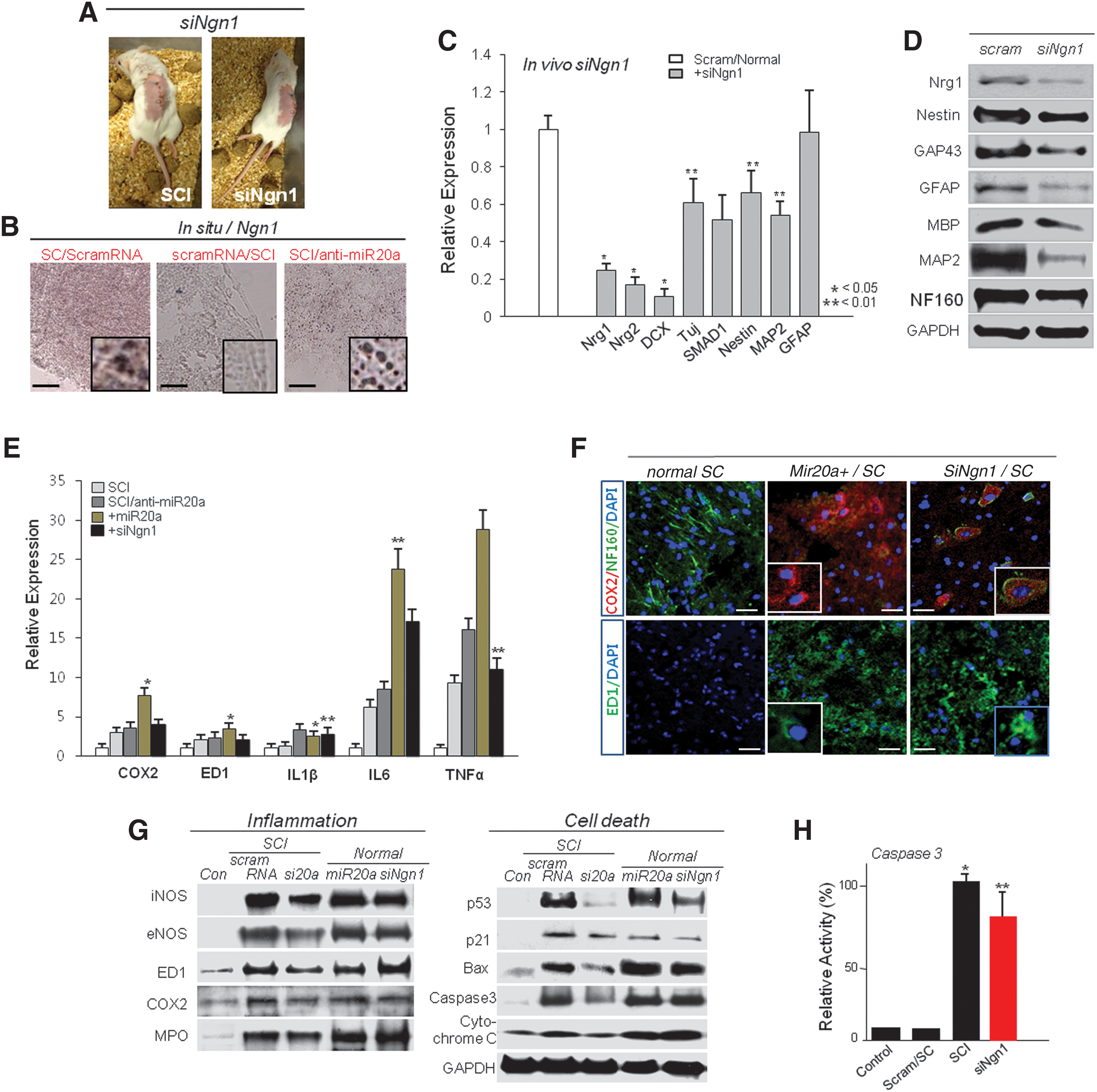
Silencing of the functional target gene, Ngn1, effectively induced inflammation that mediated secondary injury.
Additionally, when we inhibited Ngn1 expression in cultured NPCs, Oct4, Sox2, and Nanog, as well as CDK2, CDK4, and Runx3 expression were prominently down-regulated along with DCX, SMAD1, and GAP43 and expression of several neuronal markers at the transcriptional and protein levels (Fig. 5A and B). Also knock-down of Ngn1 expression completely blocked TuJ, GFAP, MBP, and MAP2ab expression at the protein level (Fig. 5B). SiNgn1 effectively induced apoptotic cell death of cultured NPCs with TUNEL-positive signature (Fig. 5C). When we evaluated the functional roles of Ngn1 in NPC self-renewal, we induced inhibition of Ngn1 expression in NPCs. As a result, self-renewal activity of NPCs was prominently decreased (Fig. 5D). Evaluation of the neurogenic activity, including neurosphere formation in siNgn1 or miR20a-treated or normal NPCs and miR20a- and siNgn1-treated cells, showed significantly diminished neurosphere formation, with depleted neurogenesis (Fig. 5E). We also showed that inhibition of miR20a significantly induced Ngn1-mediated NPC self-renewal, survival, and neurogenesis in SCI lesions via Jak/STAT3 phosphorylation (Fig. 5F; Supplementary Fig. S4). Further, AG490 treatment in NPCs induced down-regulation of Jak/STAT3, PI3K/Akt, and MAPK phosphorylation but also Oct4, Sox2, Nanog, and c-myc down-regulation along with p53 increase (Fig. 5G).

Silencing of the functional target gene, Ngn1, effectively induces attenuation of self-renewal and differentiation in cultured NPCs.
Neural regeneration by exogenic Ngn1 for functional recovery in SCI
To examine the functional effect of Ngn1 in SCI, we analyzed direct regulation of expression of genes for inflammation-inducing factors, motor neurons, and myelin regeneration for functional behavior in Ngn1-infused SCI animals (Fig. 6). Treatment with exogenous Ngn1 ameliorated the traumatic damage otherwise observed after traumatic SCI. After 7 days of Ngn1 infusion into the SCI lesion, there were significantly increased mature neuronal markers, TuJ, NF160, MAP2ab, and MBP-positive myelin in the levels of transcripts and proteins (Fig. 6B, C). When we evaluated expression of inflammation-inducing factors before and after Ngn1 infusion in SCI animals, Ngn1 infusion did not affect expression (data not shown). Additionally, 4 weeks after traumatic SCI, Ngn1-infused animals recovered from paralysis, consistently supported their own weight during planar stepping, and had a predominantly rotated paw position during walking. In contrast, injured animals injected only with scrambled RNA had limited joint locomotion (Fig. 6D). Moreover, exogenic siNgn1 infusion in SCI tissue resulted in significantly down-regulated Ngn1, Ngn2, DCX, Nestin, TuJ, and MAP2ab expression compared to siNgn1 untreated SCI tissue at the transcription level (Supplementary Fig. S5).
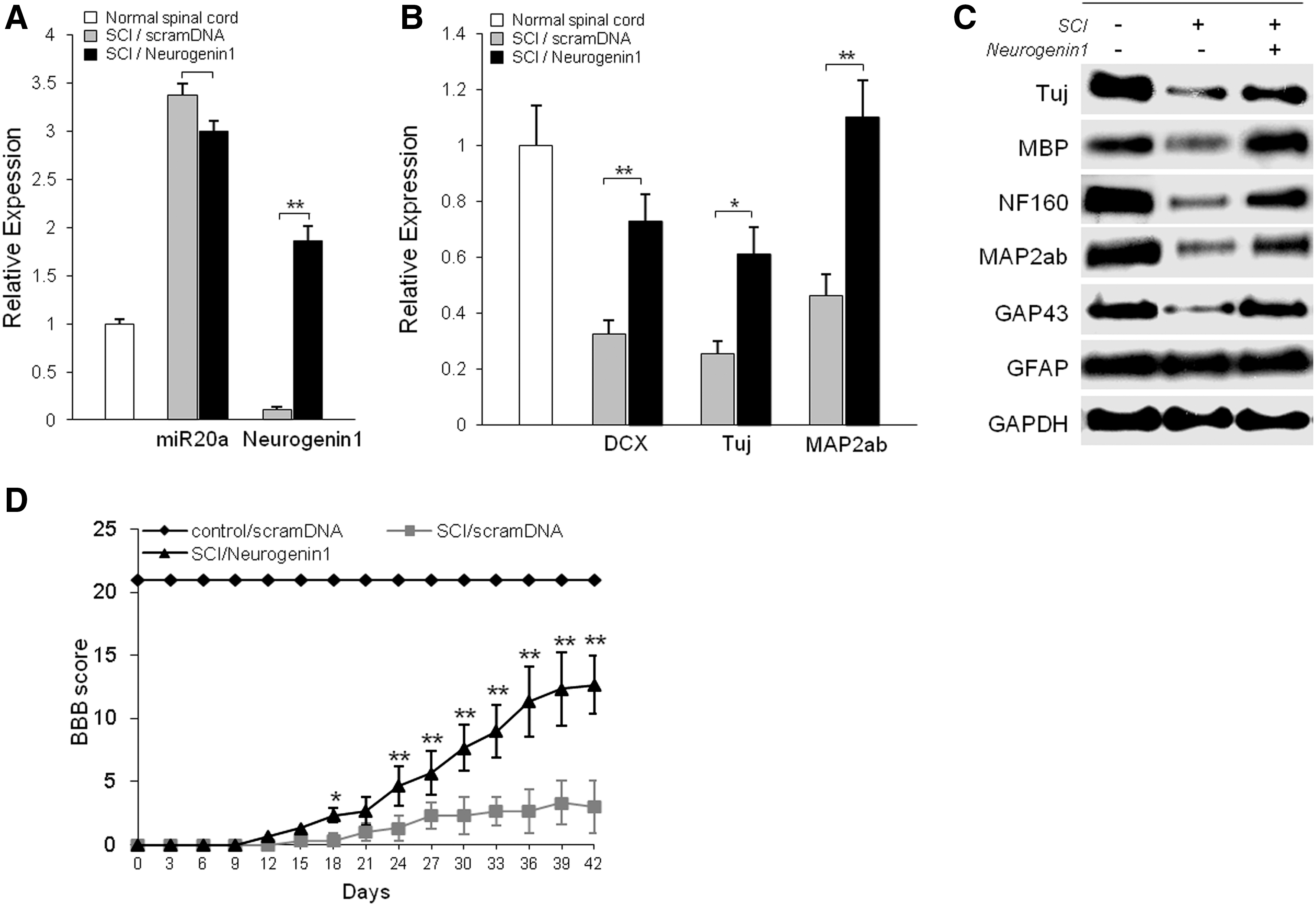
Rescue of Ngn1 expression in SCI significantly increases neuroregeneration of motor neurons.
Discussion
Noncoding miRNA has newly discovered roles in the biology of several diseases. These noncoding small RNAs serve as modulators of genes involved in almost all biological pathways, such as development, cell differentiation, cell self-renewal, cell migration, and cell death. Global gene regulation by miRNAs occurs at the post-transcriptional level by either specific inhibition of translation or by induction of mRNA cleavage (Chekulaeva and Filipowicz, 2009). Moreover, miRNAs not only regulate gene expression at the post-transcriptional level, but they are also capable of chromatin modification (Bala et al., 2009). As in the current study, many cellular and physiological processes in disease have been associated with changes in miRNA expression (van Rooij et al., 2006; Zhao et al., 2007).
SCI treatment is a major medical issue. Great efforts have been made to improve the functional deficits, including paralysis (Schaefer et al., 2007). Several therapeutic approaches to alter the microenvironment of injured spinal cord and/or to stimulate endogeneous repair mechanisms are being actively explored. Importantly, during secondary injury progression, demyelination, which induces neuronal cell death, gradually increased after onset of trauma. These notable physiological changes may continue for years after injury and include strong suppression of spontaneously occurring neuronal precursor neurogenesis, finally leading to paralysis. Each of these processes contributes to deficits in nerve conduction (Xu et al., 2001). A majority of the research has focused on chemical agents for neuroprotection against secondary tissue damage in SCI, but it remains controversial. We found that direct administration of simiR20a following traumatic injury of mouse spinal cord significantly improved recovery of hindlimb function, as measured by modified BBB; motor performance in our model of lower thoracic SCI is largely due to loss of white matter axonal tracts (Blight, 1983; Scott et al., 2005).
Our study indicates that inhibition of miR20a is therapeutically beneficial in that it prevents inflammation-mediated secondary injury, including neuron and myelin degeneration, while also simulating endogeneous NPC self-renewal and neurogenesis. simiR20a-treated animals maintained a large volume of white matter at the injury lesion, thus reducing the overall size of the lesion and leading to significantly decreased blood clotting (Fig. 2). For confirmation of the protective effect of simiR20a, we showed that the inhibition of miR20a expression resulted in limited cellular apoptosis shown by decreased caspase 3 and 9, together with a reduction in the number of TUNEL-positive cells (Figs. 2 and 3). Caspases are known to be important intracellular signaling molecules that promote apoptosis after traumatic SCI. Caspase 3 is known to directly declare Poly ADP ribose polymerase (PARP), which blocks DNA repair but also preserves the cellular pools of ATP that may be necessary for programmed cell death (LaPlaca et al., 2001). For therapeutic recovery, the proneural protein Ngn plays on important role in neurogenesis in the dorsal telencephalon, where it has been shown to commit multipotent progenitors to neuronal development and also inhibit other types of neural differentiation. Moreover, when we inhibited Ngn1 expression in normal mouse spinal cord, Ngn1 knock-down mice showed a traditional SCI phenotype that included paralysis of hindlimbs and attenuated neurogenesis along with down-regulation of the neuronal proteins TuJ, GAP43, MBP, MAP2ab, and NF160. Based on our in vitro culture of NPCs, inhibition of Ngn1 and miR20a expression blocks neurogenic gene expression and also effectively inhibits neurosphere formation and neurogenesis (Fig. 5). Other key target genes of miR20a, specifically Ngn1 and Ngn2, have very important roles in traumatic injury lesion sites because simiR20a-mediated Ngn1 up-regulation enhanced neuronal differentiation and NPC self-renewal and was also involved in mature neuron protection (Fig. 5). We found that miR20a induced SCI-like symptoms that included infiltration of microglial cells that produce various reactive nitrogen species that induce cytotoxic factors. Infusion of miR20a or siNgn1 in spinal cord effectively induced apoptotic neural cell death and also induced infiltration of ED1- or Cox2-positive microglial/macrophage cells and that finally led to production of various inflammatory cytokines (Fig. 4).
A recent report indicated that proneural bHLH (basic helix and loop helix) protein regulates progenitor cell self-renewal and the cells' progression through the neurogenic program throughout the nervous system (Bertrand et al., 2002). Interestingly, our study confirmed that siNgn1 directly induces secondary injury, including aggressive inflammation in the spinal cord. We found that an siNgn1 infusion in normal mouse spinal cord induced symptoms of SCI that were very similar to our traumatic SCI model: complete paralysis and extensive inflammation inducing motor neuron and myelin degeneration were evident 3–5 days after siNgn1 infusion (Fig. 4). In situ hybridization in injured spinal cord showed that Ngn1 expression was dramatically depleted but normal, and injured, miR20a-depleted spinal cord tissue showed very similar overexpression of nuclear Ngn1 (Fig. 4). The key miR20a target gene, Ngn1, has a critical role in the maintenance of cell survival, self-renewal, neurogenesis, and modulation of secondary injury in injured spinal cord. In the in vivo microenvironment of the injured spinal cord the response mirrored that of the cultured NPCs; knockdown of Ngn1 expression prominently down-regulated DCX, Tuj, SMAD, Nestin, and MAP2ab expression 48 hr after of siNgn1 infusion in normal animals (Fig. 4). Moreover, inhibition of Ngn1 effectively down-regulated the regenerative marker GAP43 and the neuronal markers TuJ, GFAP, MBP, MAP2ab, and NF160 along with microglial cell infiltration and apoptotic ligand secretion in the SCI lesions (Fig. 4). We observed concomitant repression of Ngn1 expression after induction of traumatic SCI. Inhibition of Ngn1 by siNgn1 infusion in normal spinal cord effectively enhanced cytotoxic effects of motor neuron and myelin and also constituted pathologic microenvironment that increased IL6, TNFα, IL1β, and COX2 expression in normal spinal cord (Fig. 4). Moreover, ED1-positive microglia cells were effectively increased after infusion of siNgn1 (Fig. 4). Finally, we confirmed that inhibition of Ngn1 was associated with an induction of motor neuron apoptosis similar to traditional secondary injury activation (Fig. 4). Moreover, the target gene of miR20a, STAT3, is very important for self-renewal of endogeneous NPCs in the spinal cord and that induces stemness gene expression after binding on functional gene promoters (Fig. 3). In particular, our study confirmed that STAT3 is involved in TuJ-positive cell self-renewal after traumatic injury (Fig. 5) as shown by inhibition of miR20a expression at the lesion site. Furthermore, Jak2/STAT3 and Raf/ERK1/2 signaling promote cell survival and inhibit cell apoptosis. Among the targets of miR20a, genes underlying Jak/STAT3 phosphorylation processes are the most important during stem cell reprogramming. When we inhibited miR20a expression in vitro and in vivo, not only Jak2/STAT3 activation, but also expression of stemness genes, Oct4, Sox2, and Nanog were recovered along with highly increased BrdU-positive, mitotic cells in SCI lesions. The Jak2/STAT3 phosphorylation signal is transferred to PI3K/Akt activation, which directly up-regulates stemness gene expression in the nucleus and finally induces NPC self-renewal in lesions before neurogenesis induction by Ngn1. For the sake of follow-up NPC reprogramming in vitro, we determined population density of S phase cells before and after simiR20a treatment.
Our study demonstrates in an animal model that inhibition of miR20a in traumatic SCI prominently decreases apoptosis and reduces functional deficits through up-regulated expression of the major target gene, Ngn1. Additionally, functional deficit in miR20a-inhibited or Ngn1-infused SCI animals was significantly ameliorated, as was tissue damage, and hindlimb reflexes were recovered. Finally, a higher percentage of these mice regained motor function compared to untreated SCI mice. Our finding that inhibition of miR20a expression effectively promotes recovery of motor function in an animal model suggests novel drug targets for treating SCI in humans. Therapeutic targeting of miR20a expression in SCI effectively prevented traumatic damage that would otherwise induce endogenous neural cell death and block activation of endogenous neural cell regeneration (Supplementary Fig. S6).
Footnotes
Acknowledgments
This work was supported by the 21st Century Frontier/Stem Cell Research Committee (SC5110) and was also supported by the National Research Foundation of Korea (NRF) grant funded by the Korea Government (MEST, 2010-0020265).
Author Disclosure Statement
The authors declare no conflict of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.