
Abstract
Malignant gliomas remain refractory to treatment despite advances in chemotherapy and surgical techniques. Conditionally replicating adenoviral vector (CRAd) could kill the tumor cells by selectively replicating in neoplastic cells, which represents a novel strategy for tumor therapy. Although CRAd with a 24-bp deletion in CR2 of the E1 region (CRAd5-D24) has been shown to have a better therapeutic effect over the other types of CRAd vectors, the current CRAd5-D24 still has some shortcomings for an efficient therapy of gliomas. In this study, we developed for the first time a novel vector CRAd5/11-D24.TRAIL/arresten by the following strategies: (1) modify CRAd5-D24 with Ad5/11 chimeric fiber to improve its infection efficiency for glioblastoma; and (2) insert two transgene expression cassettes into the E3 region and the region between the fiber and E4, respectively, for an enhanced therapeutic effect. The results indicated that the CRAd5/11-D24.TRAIL/arresten achieved nearly complete inhibition of glioma growth in nude mice possibly by increased antiangiogenesis and enhanced tumor apoptosis. The vector is the first reported E1A D24-deleted, Ad5/11 chimeric, and dual-armed oncolytic virus that shows markedly improved antitumor activities compared with the conventional oncolytic viruses. This novel antitumor agent should be evaluated further in future preclinical and clinical studies.
Introduction
CRAd is emerging as a promising tool in cancer therapy. It can infect cancer cells, multiply selectively within them, cause cell death, as well as release progeny viral particles that infect neighbor cancer cells. One of the common strategies used to design oncolytic adenoviruses is to modify E1A protein. CRAd with a 24-bp deletion in the CR2 of the E1 region (CRAd5-D24) replicates selectively in cells with abnormalities of the Rb pathway and consequent G1-S checkpoint, which was found to be present in over 30% of malignant gliomas (Jiang et al., 2009). Previous studies have demonstrated the feasibility and safety of administering CRAd5-D24 to cancer patients (Zhang et al., 2007; Kimball et al., 2010).
CRAd cell entry generally is dependent on the coxsackie-adenovirus receptor (CAR). CAR is the natural receptor for adenovirus, but is typically underexpressed in many tumor cells, such as glioma (Ulasov et al., 2006). Diminished viral cell-surface receptors may have contributed substantially to the limited clinical responses noted with adenovirus type 5 (Ad5)-based tumor gene therapy. To overcome this limitation, adenovirus can be engineered to infect cells using alternative receptors that are overexpressed at higher levels in tumors compared with surrounding healthy tissues (Ganesh et al., 2009). CD46 is overexpressed in malignant brain tumors. A retargeted vector that uses CD46 as a cellular receptor increased the level of glioma transduction by the modified virus (Nandi et al., 2009). It has been demonstrated that replacing the Ad5 shaft and knob with those of Ad11, which recognize the highly abundant CD46 receptor on tumor cells, significantly improved viral infectivity to human cells (Yu et al., 2007; Gustafsson et al., 2010).
Although virotherapy has made several progressions in its selectivity by modification of the E1 region and target ability by modification of capsids, including alterations of fiber, hexon, penton base, or protein IX, the efficacy of virotherapy still needs to be further improved. Gene therapy combined with virotherapy can be the strategy of choice. As the oncolytic viruses can target and replicate in tumors by several thousandfold, the transgenes inserted in the CRAd can also be expressed at significantly increased levels in the target cells (Xiao et al., 2010). TRAIL belongs to the tumor necrosis factor (TNF) cytokine family and is capable of inducing apoptosis in a variety of cancer cells, while producing negligible effects on normal cells (Siegelin et al., 2009). Arresten is a 26-kDa antiangiogenic fragment from the α1 chain of type IV collagen, which can inhibit endothelial cell proliferation, migration, tube formation, and Matrigel neovascularization (Nyberg et al., 2008).
In this study, we developed a novel CRAd with a chimeric Ad5/11 fiber, CRAd5/11-D24, which carries TRAIL and arresten genes, with the aim of developing a new therapeutic strategy for malignant glioma. This novel CRAd vector contains two expression cassettes. One is the phosphoglycerate kinase (PGK)–mini-cytomegalovirus (mini-CMV) expression cassette carrying TRAIL and the enhanced green fluorescent protein (eGFP) gene, which is placed between the fiber and E4 region. The other is a CMV-arresten-SV40 cassette expressing arresten transgene, which is inserted in the E3 region. The results showed that the novel oncolytic vector CRAd5/11-D24.TRAIL/arresten had significantly improved antitumor activities compared with the control vectors in vitro and in vivo.
Materials and Methods
Cells and culture conditions
The human malignant glioma cell lines U87, U251, and A172 and human embryonic kidney (HEK) cell line 293 were purchased from American Type Culture Collection (ATCC, Manassas, VA). The firefly luciferase-expressing glioma cell line U87-luc was developed by our laboratory. All cell lines were maintained in a humidified 37°C atmosphere of 95% air and 5% CO2 and cultured in Dulbecco's modified Eagle's medium supplemented with 10% heat-inactivated fetal calf serum, 100 U/ml penicillin, 100 μg/ml streptomycin, and 200 μg/ml L-glutamine.
Construction of recombinant adenoviral vectors
The human Ad5 and Ad11 fiber gene sequences were acquired from NCBI GenBank (accession nos. M18369.1 and AB162822). The shuttle vector plasmid for a chimeric fiber Ad5/11 vector, pShuttle-Ad5/11F-E4, was constructed as described below. First, we synthesized a gene that includes Ad5 tail (522–608 bp), Ad11 shaft and knob sequence in pcDNA 3.1(+) (Sangon, Shanghai, China). Unique NdeI and SpeI restriction sites were introduced separately in its 5′ and 3′ end. The pcDNA 3.1(+)/Ad5/11F digested by NdeI and SpeI was ligated with an NdeI/SpeI-linearized pBS shuttle vector (Xia et al., 2000) at a 4:1 (vol/vol) ratio. The positive clones that displaced Ad11 shaft and knob were screened by enzyme digestion and DNA sequencing, named pBS/Ad5/11F. Second, we constructed a single expression cassette that includes a CMV promoter, an eGFP gene, and a SV40 polyA. Meanwhile, a double-expression cassette was constructed such that the PGK drives the soluble TRAIL and a mini-CMV promotes the eGFP gene. The two cassettes were digested by BamHI and SfuI and ligated with the pBS/Ad5/11F separately. Otherwise, we constructed an E3-deletion shuttle vector used for introducing the arresten gene. For the generation of the final adenovirus vector, the adenovirus backbone with the deletion of 24 bp in CR2 of the E1 region digested with SwaI and the pShuttle-E3 digested with PacI were cotransformed into Escherichia coli BJ5183 cells. The positive resultant plasmid digested with SwaI was cotransformed into E. coli BJ5183 cells along with the PacI-linearized pBS/Ad5/11F containing a cassette for dual expression of eGFP and TRAIL, and the positive clones were screened by enzyme digestion and DNA sequencing. The resulting plasmid was designated pAd5/11-D24.TRAIL/arresten. The CRAd5/11-D24.TRAIL/arresten virus was produced by transfecting PacI-digested pAd5/11-D24.TRAIL/arresten vector into HEK293 cells. The virus was propagated in HEK293 cells and purified by cesium chloride gradient methods. The titers were detected by spectrophotometry at an absorbance (A) of 260 nm. The viruses CRAd5-D24, CRAd5/11-D24, CRAd5/11-D24.TRAIL, and CRAd5/11-D24.arresten were produced using a similar strategy, as mentioned above for the production of CRAd5/11-D24.TRAIL/arresten (Fig. 1).
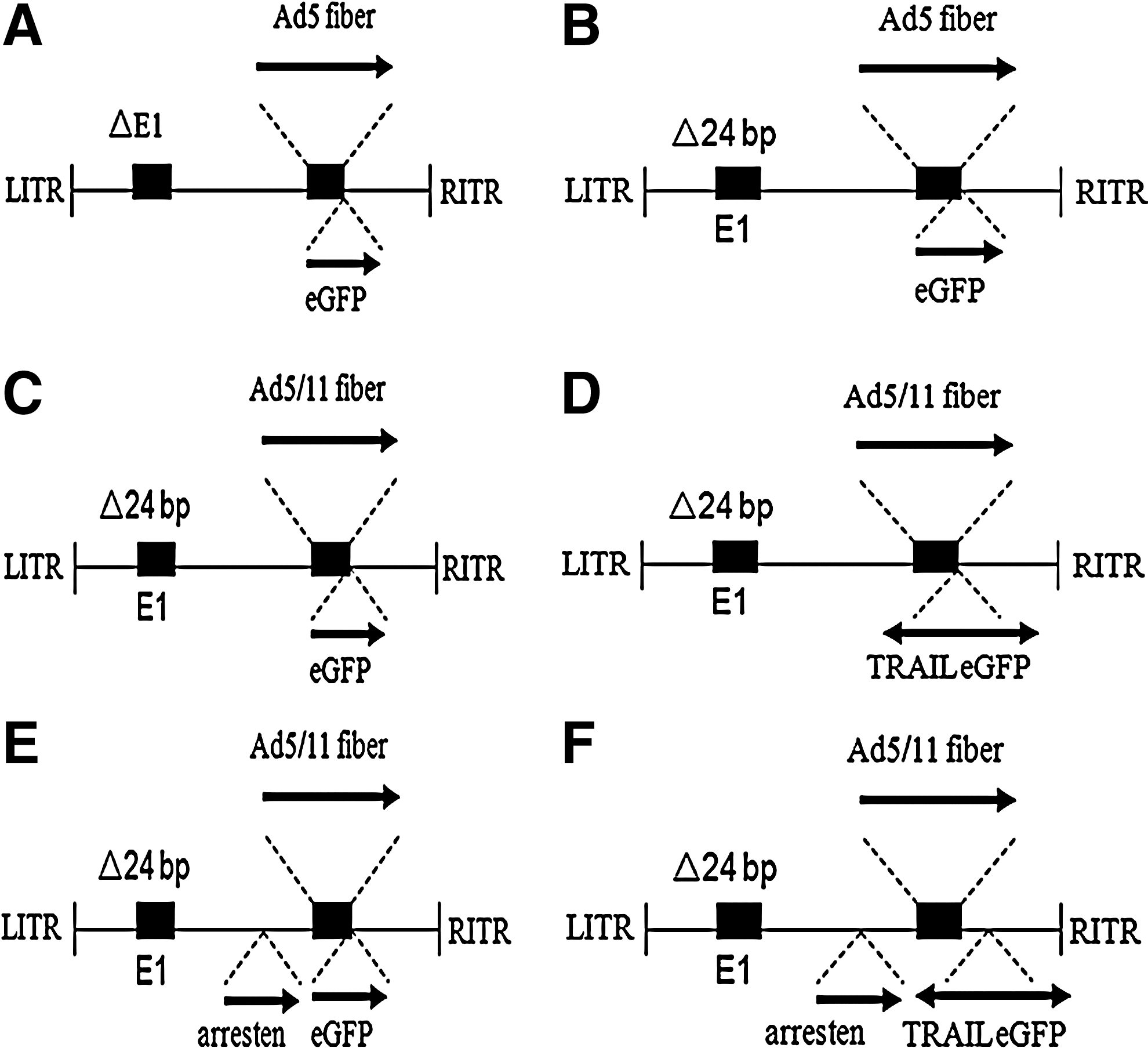
Schematic map of the adenoviral vectors:
Cytopathic assay
U87, U251, and A172 tumor cells were plated at subconfluent levels in a 24-well plate (Corning, NY) and infected with Ad5-eGFP, CRAd5-D24, CRAd5/11-D24, CRAd5/11-D24.TRAIL, CRAd5/11-D24.arresten, and CRAd5/11-D24.TRAIL/arresten at various multiplicities of infection (MOIs). At 7 days after infection, the medium was removed and cells were exposed to 2% crystal violet in 20% methanol for 15 min. Then plates were washed with distilled water and documented as photographs.
Cell viability assay
Cells were plated on 96-well plates at 5×103/well, 1 day prior to virus infection. Cells were then infected with different viruses at the concentration of 10 MOI or with PBS only. Every 24 hr later, the cell survival rate was evaluated by a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. After incubation at 37°C for 4 hr, all media and MTT solutions were gently removed, and 150 μl of dimethyl sulfoxide was added to each well and mixed thoroughly on a concentrating table for 10 min. The absorbance from the plates was read at 570 nm with a Multiskan MK3 (Thermo Electron Corporation, Madison, WI).
Analysis of adenovirus transduction
Cells were plated overnight at a density of 5×105/well in a 60-mm plate. For about 24 hr, Ad5-eGFP, CRAd5-D24, CRAd5/11-D24, CRAd5/11-D24.TRAIL, CRAd5/11-D24.arresten, and CRAd5/11-D24.TRAIL/arresten were applied to the U87 cells at the dosage of 500 viral particles/cell. For quantification of eGFP-positive cells, photographs were taken at 24 hr post infection with a 5×objective (3-sec exposure time). The infection efficiency was quantified by counting eGFP-positive cells.
Animal experiments
Animal experiments were performed according to the National Institutes of Health Guide for the Care and Use of Laboratory Animals. Four- to 6-week-old female BALB/c nude mice were obtained from the Animal Research Committee of the Institute of Biochemistry and Cell Biology (Shanghai, China). The xenograft tumor model was established by subcutaneously injecting 2×106 U87-luc cells in a 150-μl volume into the right hind flank of the mice. Tumor volumes were measured and calculated as length×width2/2. When the tumors reached a volume of approximately 70–100 mm3, animals were randomly assigned to treatment groups and received every other day four injections of 5×108 plaque-forming units (PFU) of virus each time. The tumor volumes of the seven mice in each group were measured every 2 days for about 1 month.
Bioluminescence imaging and analysis
Real-time tumor growth was monitored by optical imaging using a Xenogen-IVIS cooled CCD optical system (Xenogen, Alameda, CA). Mice were anesthetized with 3% isoflurane after intraperitoneal (i.p.) injection with 75 mg/kg body weight D-luciferin firefly (Xenogen). Five minutes after injection of the D-luciferin, images were acquired for 1 sec using Living Image analysis and acquisition software (Xenogen). A photographic image was taken onto which the pseudocolor image representing the spatial distribution of photon counts was projected. For bioluminescence imaging plots, photon flux was calculated for each mouse by using a square region of interest encompassing the right hind flank of the mouse in a side lying position.
Histology and immunohistochemistry
Tumors obtained from mice treated with different viruses were immediately fixed in formalin (4% formaldehyde–PBS) and subsequently embedded in paraffin. Two-millimeter-thick sections were embedded in paraffin and stained with hematoxylin and eosin. Tumor sections were assayed by immunohistochemical analysis with rat anti-mouse CD31 antibodies (1:500 dilution; BD Biosciences Pharmingen, San Diego, CA). The TUNEL (TdT-mediated dUTP nick end labeling) assay was performed according to the apoptosis detection kit manual (Keygen Biotech, Nanjing, China).
Statistical analysis
Statistical analysis was performed using the Statistical Package for the Social Sciences (SPSS) software (version 13; Chicago, IL). Differences between each control and experimental groups were analyzed by using one-way analysis of variance between groups (ANOVA/LSD), and p<0.05 was considered statistically significant.
Results
Cytotoxic effect of CRAd5/11-D24 against glioma cells in vitro
Previous studies suggested that a chimeric adenoviral vector Ad5/11 showed improved infective capability and was considered a promising gene-therapy vector candidate (Yu et al., 2007). In this study, to evaluate the cytotoxicity of the fiber-modified oncolytic adenoviruses in glioma cells, U87, U251, and A172 were infected with CRAd5-D24, CRAd5/11-D24, or replication-deficiency adenovirus Ad5-eGFP at MOIs of 0, 0.1, 0.5, 1, 5, and 10. Cells were stained with crystal violet 7 days after infection. Significant cytotoxicity was observed in U87 and A172 cells infected with CRAd5/11-D24, compared with the cells infected with CRAd5-D24 or Ad5-eGFP (Fig. 2). The cytopathic effect of CRAd5/11-D24 was about 100 times greater than that of Ad5-eGFP in the U87 and A172 cell lines.
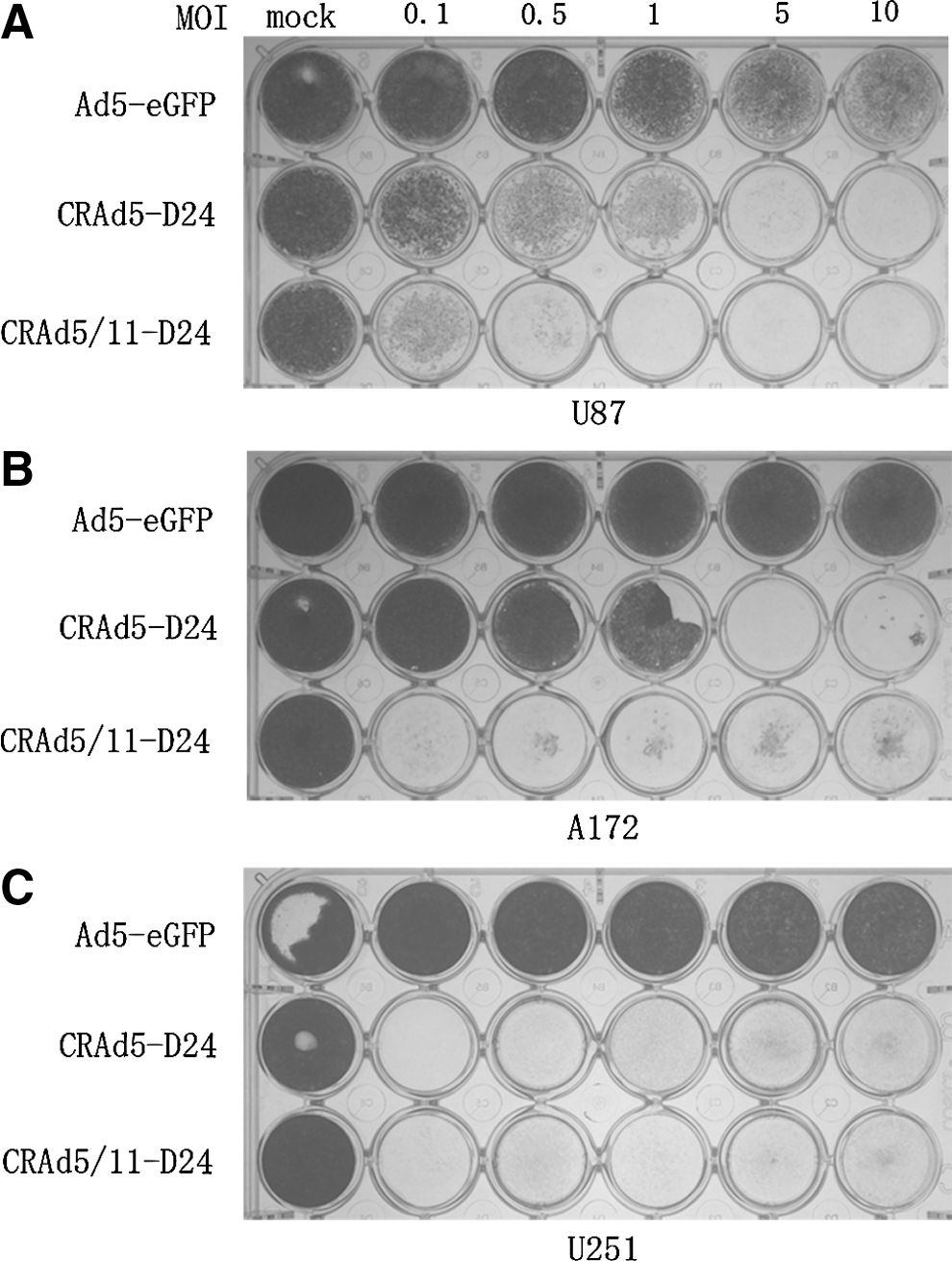
Cytopathic effects of Ad5-eGFP, CRAd5-D24, and CRAd5/11-D24 on
To further evaluate the kinetics of cytotoxicity induced by CRAd5/11-D24, glioma cells (U87, U251, and A172) were plated in 96-well plates and infected with Ad5-eGFP, CRAd5-D24, and CRAd5/11-D24 at 10 MOI. Tumor cell viability was determined by MTT assay. The data showed that the virus could induce time-dependent cytotoxicity in all three tumor cell lines. The cytotoxic effect of the adenovirus with chimeric fiber 5/11 modification in U87 and A172 cells was more apparent than that of CRAd5-D24 and Ad5-eGFP (Fig. 3).

Kinetics of cell killing by Ad5-eGFP, CRAd5-D24, and CRAd5/11-D24 in
To confirm that the improved cytotoxic effect of Ad5/11 fiber chimeric CRAd in glioma cells was due mainly to a better transduction of target cells, Ad5-eGFP, CRAd5-D24, CRAd5/11-D24, CRAd5/11-D24.TRAIL, CRAd5/11-D24.arresten, and CRAd5/11-D24.TRAIL/arresten were applied to U87 cells that express a high level of CD46, the receptor of Ad11. The results showed that the percentage of eGFP-positive cells in CRAd5/11-D24 groups was higher than that in the control groups Ad5-eGFP and CRAd5-D24 (Fig. 4). In contrast, the infectivity of CRAd5/11-D24 showed little difference from that of Ad5-eGFP and CRAd5-D24 in HEK293 cells and U251 cells, both of which have a high level of CAR and a relatively low level of CD46. In addition, the modification of the adenoviral vector did not affect the virus titer (data not shown).
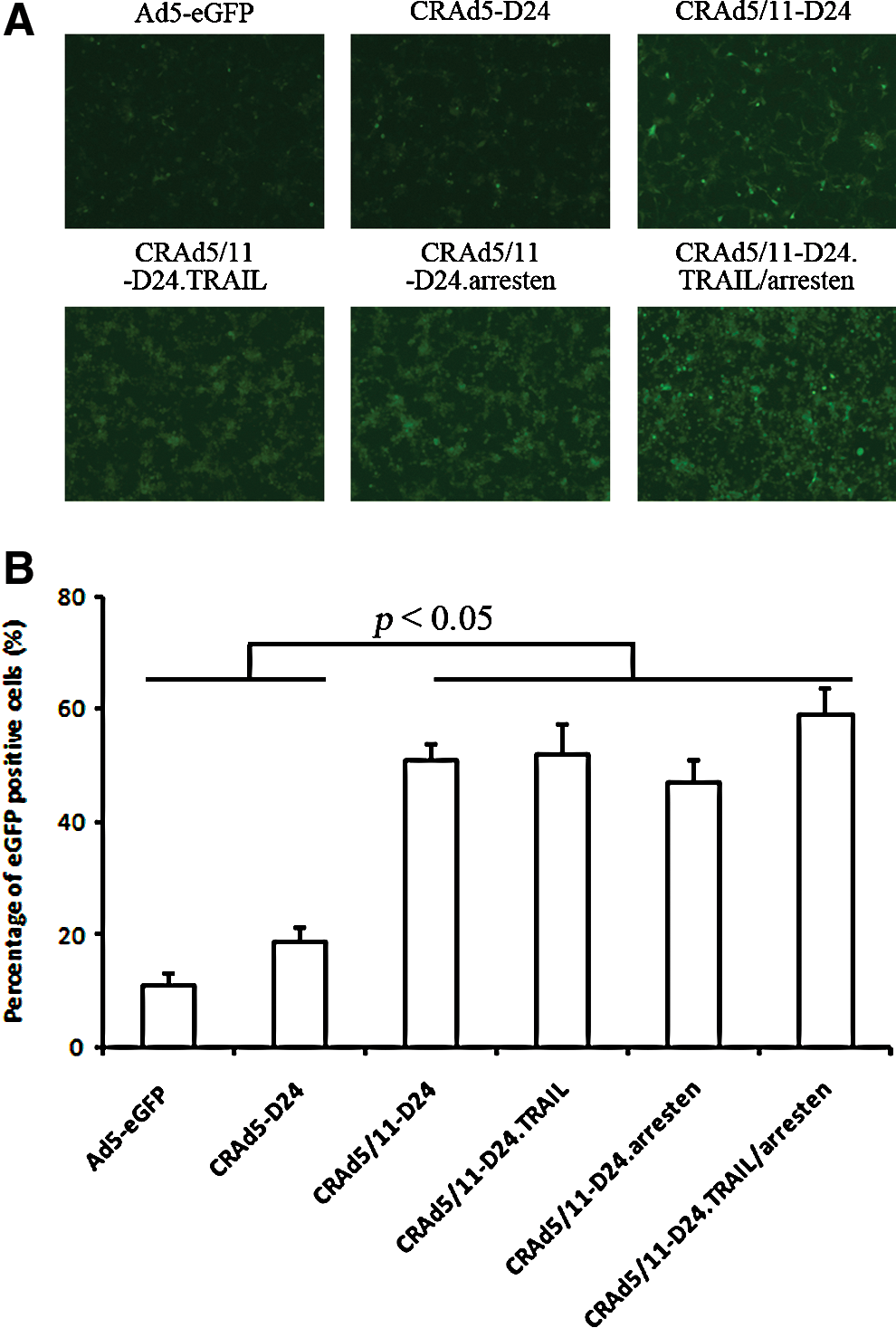
Comparison of transduction efficiency of Ad5-eGFP, CRAd5-D24, CRAd5/11-D24, CRAd5/11-D24.TRAIL, CRAd5/11-D24.arresten, and CRAd5/11-D24.TRAIL/arresten in the U87 cell line.
Therapeutic effect of CRAd5/11-D24.TRAIL/arresten in vivo
We next used a glioma xenograft model in nude mice to test the therapeutic effect of CRAd5/11-D24.TRAIL/arresten in vivo. The human U87-luc glioma cell line was injected into the right flank of nu/nu mice. This cell line could be used to visualize the therapeutic effects of our treatments, because it stably expresses luciferase, which allows noninvasive estimates of tumor burden in vivo. Once the tumors reached a size of 70–100 mm3 (approximately 7 days later), mice were randomly divided into six groups and then treated, respectively, with intratumoral injection of Ad5-eGFP, CRAd5-D24, CRAd5/11-D24, CRAd5/11-D24.TRAIL, CRAd5/11-D24.arresten, or CRAd5/11-D24.TRAIL/arresten (5×108 PFU). Mice were then imaged after i.p. injection of D-luciferin on days 7, 19, and 30. A panel of representative images is shown in Fig. 5A. Low bioluminescent signals of U87-luc cells were detected as early as 7 days after i.p. injection, and were greatly increased by day 19. At the last imaging time point (30 days), the lowest bioluminescent signals were detected in the group treated by TRAIL and arresten gene–based CRAd5/11-D24 compared with the groups treated with nonreplicative Ad5-eGFP or other CRAd virus, indicating a stronger inhibition of tumor growth in the group exposed to the gene-virotherapy manner. Antitumor ability of the viruses was also assessed by measuring the tumor volume. In the control mice that received replicative-deficiency adenovirus Ad5-eGFP, tumor volume increased progressively over the course of the experiment. Compared with the control virus Ad5-eGFP, tumor growth was suppressed in groups treated with CRAds. In addition, the antitumor efficacy of the CRAds with chimeric fiber modification was stronger compared with that of the unmodified CRAd, CRAd5-D24 (p<0.05). Furthermore, the group treated with CRAd5/11-D24.TRAIL/arresten showed the strongest antitumor activity among all of the treatment groups (Fig. 5B). To make these observations valid, we confirmed that the oncolytic adenoviruses CRAd5-D24, CRAd5/11-D24, CRAd5/11-D24.TRAIL, CRAd5/11-D24.arresten, and CRAd5/11-D24.TRAIL/arresten have a similar replication ability in U87 cells. Furthermore, the expression of transgene TRAIL or arresten was equally efficient with oncolytic adenoviruses CRAd5/11-D24.TRAIL, CRAd5/11-D24.arresten, and CRAd5/11-D24.TRAIL/arresten in U87 cells (data not shown).
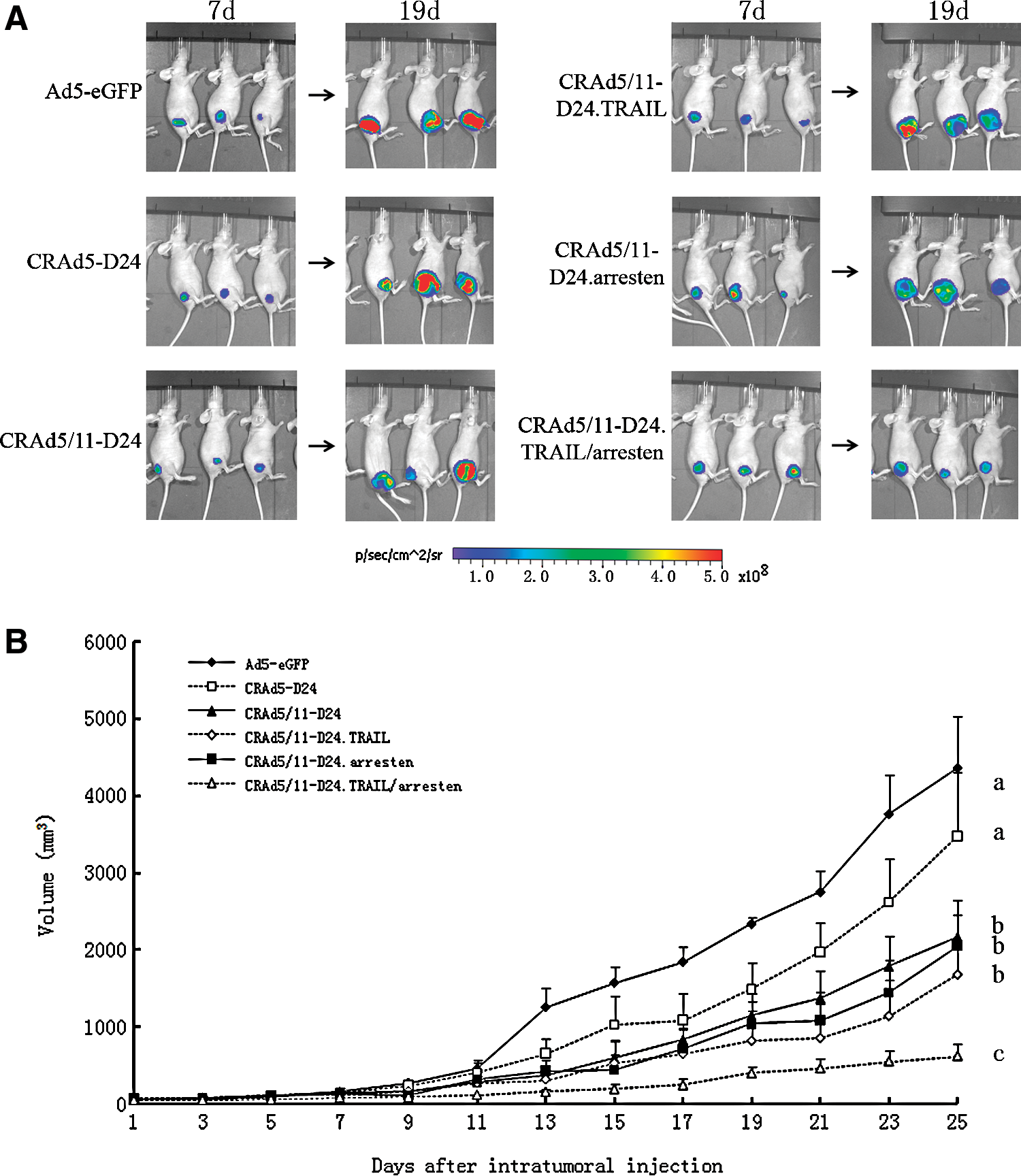
Treatment monitoring in a model of glioblastoma xenograft. Tumor models were established by injecting 2×106 U87-luc cells into the right flank of female nude mice. When tumors reached the size of 70–100 mm3, the mice were treated four times every other day by intratumoral injection of 50 μl of PBS or Ad5-eGFP, CRAd5-D24, CRAd5/11-D24, CRAd5/11-D24.TRAIL, CRAd5/11-D24.arresten, or CRAd5/11-D24.TRAIL/arresten at 5×108 PFU of virus in a 50-μl volume.
Morphologic evaluation of the inhibitory effect of CRAd5/11-D24.TRAIL/arresten on tumor growth
To understand the mechanisms underlying the antitumor effect of the novel CRAd5/11-D24.TRAIL/arresten, vessel growth and apoptosis were analyzed using CD31 and TUNEL staining in tumors treated with different viruses. As shown in Fig. 6A, the vessel growth in tumors from the groups treated with CRAd5/11-D24.arresten or CRAd5/11-D24.TRAIL/arresten was significantly suppressed compared with that from the other groups. Relative to Ad5-eGFP (positive control=100%) group, the average percentage of microvessel density (MVD) from five randomly selected visual fields in tumors treated with CRAd5-D24, CRAd5/11-D24, CRAd5/11-D24.TRAIL, CRAd5/11-D24.arresten, and CRAd5/11-D24.TRAIL/arresten was 86.3±4.8%, 79±9.3%, 71.2±4.9%, 44±5.7%, and 37.6±5.0%, respectively (Fig. 6B). In addition, as shown in Fig. 7, the tumors in the groups of CRAd5/11-D24.TRAIL and CRAd5/11-D24.TRAIL/arresten displayed much stronger apoptosis induction than those in the other groups. The average percentage of TUNEL-positive cells from five randomly selected visual fields in the Ad5-eGFP, CRAd5-D24, CRAd5/11-D24, CRAd5/11-D24.TRAIL, CRAd5/11-D24.arresten, and CRAd5/11-D24.TRAIL/arresten groups was 1.0±0.2%, 1.6±0.5%, 1.5±0.3%, 10±2.4%, 0.8±0.2%, and 11.2±1.8%, respectively.
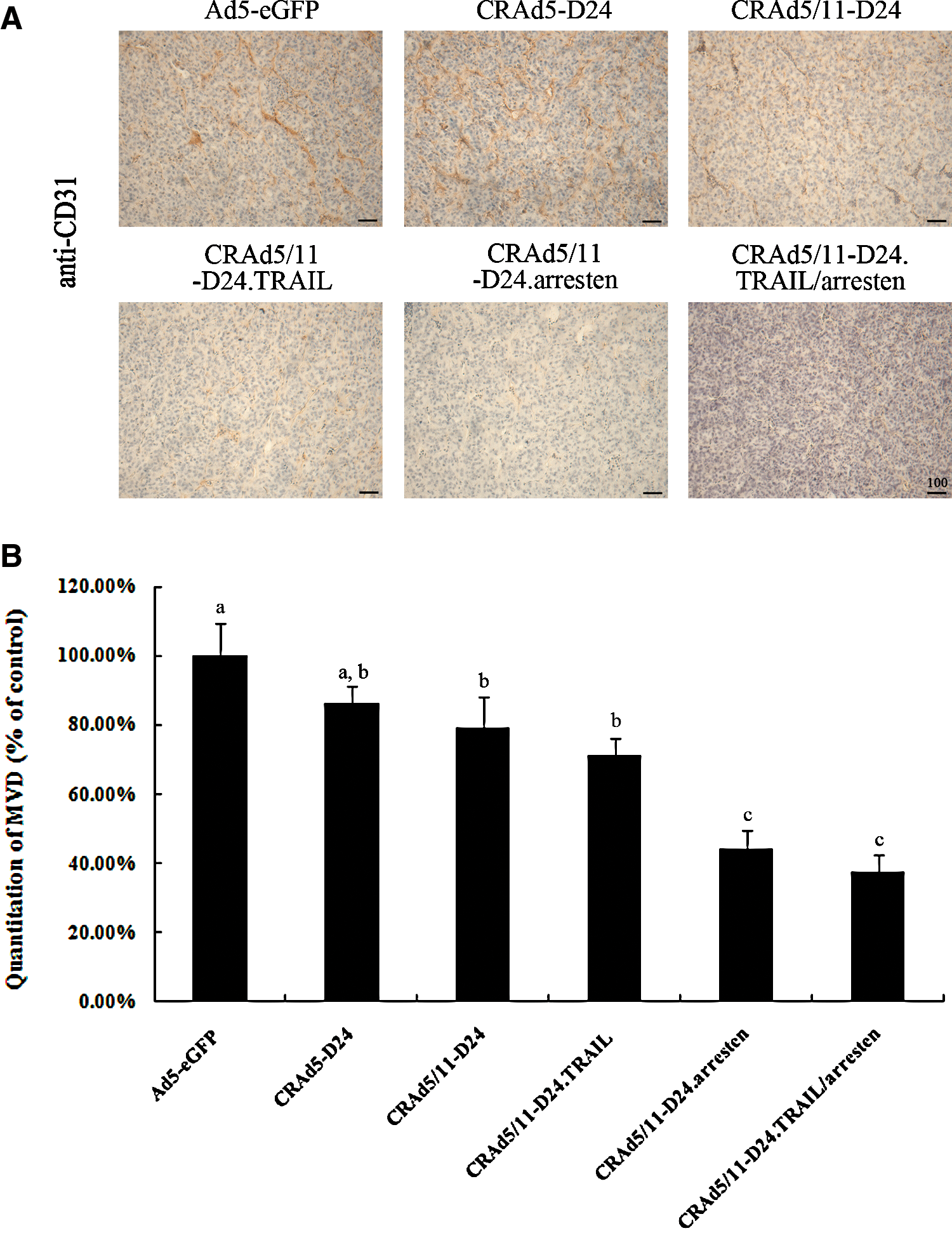
Pathological evaluation and immunohistochemical staining. Tumors from the treated mice were harvested 25 days after intratumoral viral infection.
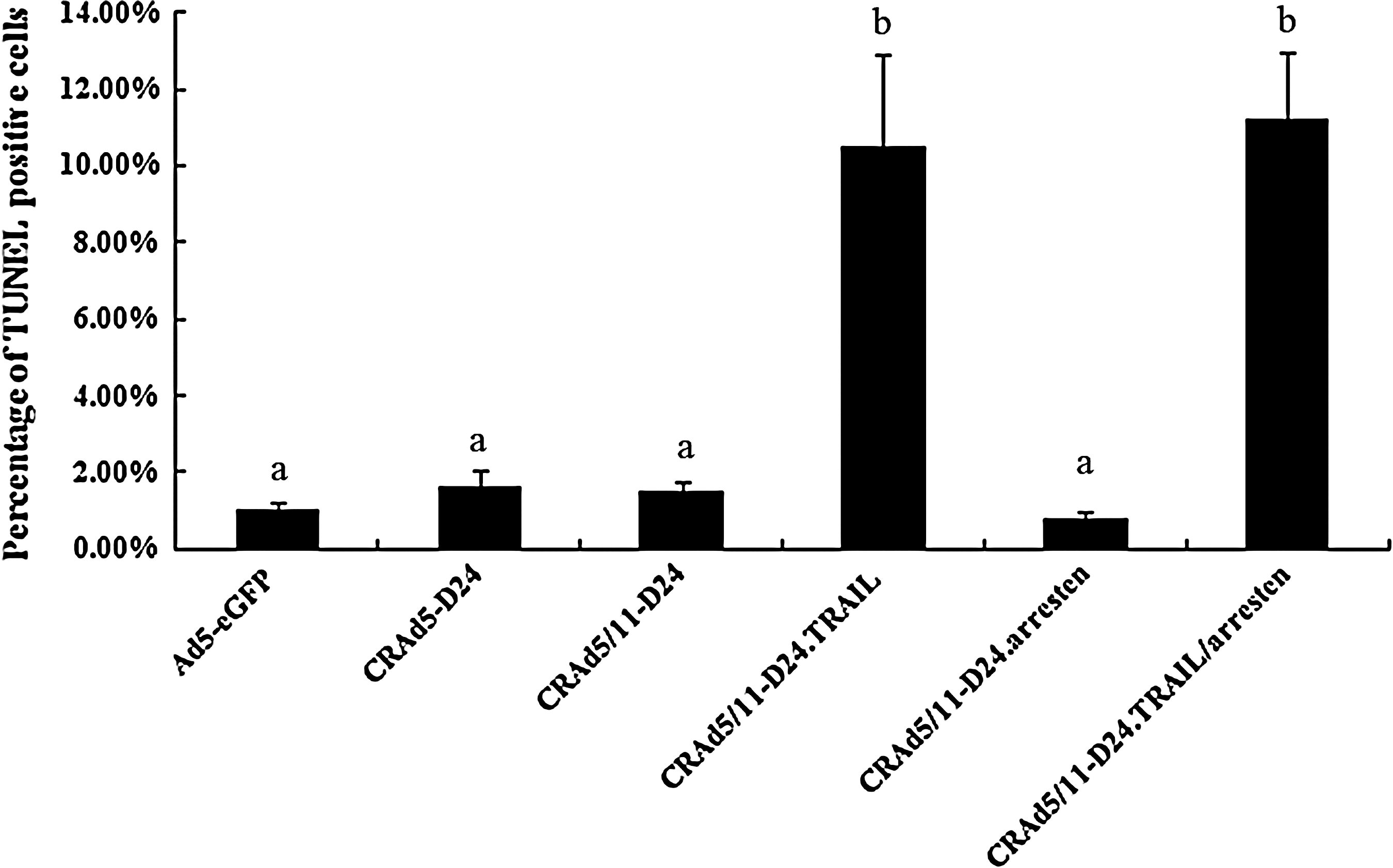
TUNEL analysis of the tumors from the CRAd-treated mice. Tumors were harvested 25 days after the injection of Ad5-eGFP, CRAd5-D24, CRAd5/11-D24, CRAd5/11-D24.TRAIL, CRAd5/11-D24.arresten, or CRAd5/11-D24.TRAIL/arresten. Sections were stained with TUNEL to reveal the apoptotic activity in the tissue. TUNEL-positive cells were quantified and presented as means±SEM. The bars that do not share a letter designation were determined to be significantly different by one-way ANOVA/LSD comparison (p<0.05).
Discussion
In this study, we successfully generated a novel oncolytic vector with 5/11 chimeric adenovirus fiber and 24-bp deletion in the CR2 of the E1 region, which additionally carries two genes, arresten and TRAIL, using two separate expression cassettes inserted into different regions of the viral vector. The data for the first time showed that this dually armed CRAd with E1A 24-bp deletion and 5/11 chimeric adenovirus fiber possessed a markedly elevated tumor-suppressing capacity against malignant gliomas.
The E1A-mutant oncolytic adenovirus carries a 24-bp (from 923 bp to 946 bp) deletion in the E1A region that is responsible for binding Rb protein. Many reports have demonstrated that this type of E1A-restricted adenoviruses could attain efficient antitumor activity in both in vitro and in vivo experiments (Jiang et al., 2005; Gomez-Manzano et al., 2006) and reveal very low toxicity to normal tissues (Page et al., 2007). The previous studies of Ulasov et al. (2006, 2007a) showed that a chimeric Ad5/3 vector rendered increased glioma transduction by binding to CD46. The adenovirus serotypes 3, 11, and 35 all belong to the species B adenovirus serotype and use CD46 as a receptor (Fleischli et al., 2007). Similarly, the novel Ad5/11 fiber chimeric CRAd constructed in this study was demonstrated to possess stronger tumor lysis capacity than the control viruses, suggesting that this fiber-chimeric CRAd can be an attractive vector for the treatment of malignant gliomas.
Gene-virotherapy combines the advantages of both gene therapy and virotherapy. As the novel oncolytic viruses targeted and replicated in tumor more efficiently compared with the control viruses, the transgenes inserted in the CRAd could also be expressed at significantly increased levels in the target cells (Xiao et al., 2010). In this study, we overcame the technical challenges and generated the 5/11 fiber-chimeric CRAd vector that for the first time carries two transgene expression cassettes. One was inserted in the E3 region expressing arresten. The other is a bidirectional expression cassette between the fiber and E4 region that expresses TRAIL and eGFP under the control of the PGK or mini-CMV promoter separately.
TRAIL, recently entered in clinical trials, is a member of the TNF superfamily and a promising candidate for cancer therapy (de Vries et al., 2006). TRAIL induces apoptosis almost specifically in cancer cells, but not in normal cells (LeBlanc and Ashkenazi, 2003). Arresten, a 26-kDa noncollagenous domain of type IV collagen α1 chain, was found to be an efficient inhibitor of angiogenesis and tumor growth in mouse models (Colorado et al., 2000; Long et al., 2008). Arresten decreases angiogenesis by affecting several distinct cell-signaling pathways, one of which involves binding to α1β1 integrin and heparan sulfate proteolycans (Colorado et al., 2000). In this article, we showed that CRAd5/11-D24.arresten exhibited enhanced antiangiogenesis and CRAd5/11-D24.TRAIL exhibited increased apoptotic activity in glioma xenografts. Furthermore, this dual-expression CRAd with the Ad5/11 chimeric fiber modification possessed a remarkably elevated tumor-suppressing capacity against glioma cells compared with traditional CRAds.
In summary, the novel adenovirus, CRAd5/11-D24.TRAIL/arresten, exhibited enhanced tropism, oncolytic capacity, and tumor-suppressing ability in malignant gliomas. The enhanced antitumor ability of this novel fiber-chimeric and dual-expression oncolytic virus warrants further evaluations in future preclinical and clinical studies.
Footnotes
Acknowledgments
This work was supported by the “Foundation for Excellent Doctor Degree Dissertation” (S2009YB04) of Shaanxi Normal University and research grants to H.X. from National Natural Science Foundation of China (nos. 30872993 and 31070137).
Author Disclosure Statement
No competing financial interests exist.