
Abstract
Six- to 8-kb mini-dystrophin genes are promising candidates for Duchenne muscular dystrophy (DMD) gene therapy. Several dual adeno-associated virus (AAV) mini-dystrophin vectors have been tested in dystrophin-deficient mice. Despite the encouraging preclinical results, none of the existing dual AAV vectors can restore sarcolemmal neuronal nitric oxide synthase (nNOS) expression. Localization of nNOS to the sarcolemma may greatly improve the therapeutic outcome in DMD (Lai, Y., Thomas, G.D., Yue, Y., et al. [2009]. J. Clin. Invest. 119, 624–635). In this study, we developed a series of dual AAV expression vectors to express a synthetic minigene that carries the nNOS localization domain. To help validate dual vector reconstitution, we also included a FLAG tag and a GFP reporter at different ends of the minigene. These dual AAV vectors were packaged in Y445F tyrosine mutant AAV-6 and tested in dystrophin-null mdx4cv mice by direct muscle injection. All dual vectors expressed GFP/FLAG-tagged mini-dystrophin and restored sarcolemmal nNOS. However, the reconstitution efficiency was significantly different among different sets. The dual vector set YZ27/YZ22 yielded the highest transduction efficiency (∼90%). Further development of this set dual vector may lead to more effective DMD gene therapy.
Introduction
Deletion is the most common mutation in DMD (Flanigan et al., 2009). Interestingly, the phenotype is much milder in some patients that carry large in-frame rod domain deletion (Gillard et al., 1989; Koenig et al., 1989; England et al., 1990; Beggs et al., 1991). Based on these clinical observations, a series of 6- to 8-kb synthetic mini-dystrophin genes have been generated (Clemens et al., 1995; Harper et al., 2002). These abbreviated genes represent excellent candidates for DMD gene replacement therapy. Unfortunately, they do not carry the nNOS-binding domain and cannot recruit nNOS to the sarcolemma. To improve the current mini-dystrophin genes, we developed the second generation minigenes that contain the R16/17 nNOS binding domain (Lai et al., 2009). Studies in transgenic mice suggest that the novel R16/17 minigenes are therapeutically superior to the ones that cannot restore sarcolemmal nNOS expression (Lai et al., 2009).
To translate findings from transgenic mice to gene therapy, we engineered one of our newly developed R16/17 minigenes (ΔR2–15/ΔR18–19) into a series of dual adeno-associated virus (AAV) vectors (Duan, 2006, 2011). To validate expression, we also fused a FLAG tag and a GFP tag to the minigene. AAV is a very small single-stranded DNA virus that has a maximal packaging capacity of 5 kb (Lai et al., 2010). In dual AAV vectors, the expression cassette was split into two parts and packaged separately. Transgene expression was reconstituted through various recombination pathways after co-infection (Duan et al., 2006). The gene split technique has allowed efficient delivery of therapeutic genes that are larger than 5 kb (Lai et al., 2005; Ghosh et al., 2008; Lostal et al., 2010; Odom et al., 2011). In this study, four independent sets of mini-dystrophin dual vectors were packaged in the Y445F tyrosine mutant AAV-6 virus (Zhong et al., 2008; Qiao et al., 2010). The transduction efficiency was compared and nNOS restoration was confirmed in dystrophin-deficient mdx4cv mice.
Materials and Methods
Animals
All animal experiments were approved by the Animal Care and Use Committee of the University of Missouri and were in accordance with National Institutes of Health guidelines. The original breeding pairs of BL6 and B6Ros.Cg-Dmdmdx-4Cv/J (mdx4cv) mice were purchased from The Jackson Laboratory. Experimental mice were generated by in-house breeding. Only adult male mice were used in the study. All mice were housed in specific-pathogen-free animal care facilities and kept under a 12-hr light (25 lux)/12-hr dark cycle with free access to food and water.
AAV production and gene delivery
The proviral cis plasmids were engineered using standard molecular biology methods. The complete expression cassette included the cytomegalovirus (CMV) promoter, GFP/FLAG fused ΔR2–15/ΔR18–19 minigene and SV40 poly-adenylation signal. The Y445F tyrosine modified AAV-6 was generated and purified using a previously reported protocol (Zhong et al., 2008; Qiao et al., 2010). Dual AAV vectors were delivered in pairs to the anterior tibialis muscle as we described before (Ghosh et al., 2006). Each co-infection consisted of 1.2×1010 vg particles/vector/muscle. AAV transduction was evaluated at 4 weeks post-injection.
Morphology studies
GFP was visualized under the FITC channel in a fluorescence microscope. Dystrophin expression was examined with Dys 3, an epitope and species-specific mouse monoclonal antibody against human dystrophin H1 (1:20; Novocastra, Newcastle, UK) (Yue et al., 2003; Lai et al., 2005; Lai et al., 2009). nNOS was detected with a polyclonal antibody (1:2000; Santa Cruz Biotechnology). In situ NOS activity staining was performed according our published protocol (Lai et al., 2009; Li et al., 2010, 2011). Photomicrographs were taken with a Qimage Retiga 1300 camera using a Nikon E800 fluorescence microscope.
Western blot
Whole muscle lysate was obtained from frozen limb muscles as we described before (Li et al., 2011). Dystrophin was detected with a monoclonal antibody against the dystrophin C-terminal domain (Dys2, 1:100, clone Dy8/6C5, IgG1; Novocastra). GFP was determined with a monoclonal antibody against GFP (1:2000, clone 3E6, IgG2a; Invitrogen). The FLAG tag was revealed using the anti-FLAG M2 antibody (1:1000; Sigma). β-actin was detected with a monoclonal antibody (1:5000; Sigma).
Statistical analysis
Data are presented as mean±standard error of mean. Statistical analysis was performed with the SPSS software. Statistical significance was determined by one-way ANOVA followed by Bonferroni post hoc analysis. Difference was considered significant when p<0.05.
Results
Design and construction of the dual AAV vectors
The 6-kb ΔR2–15/ΔR18–19 mini-dystrophin gene is engineered from the full-length human dystrophin coding sequence. It carries the major dystrophin functional domains including the N-terminal actin-binding domain, R16/17 nNOS binding domain, cysteine-rich domain, and C-terminal domain (Fig. 1A).
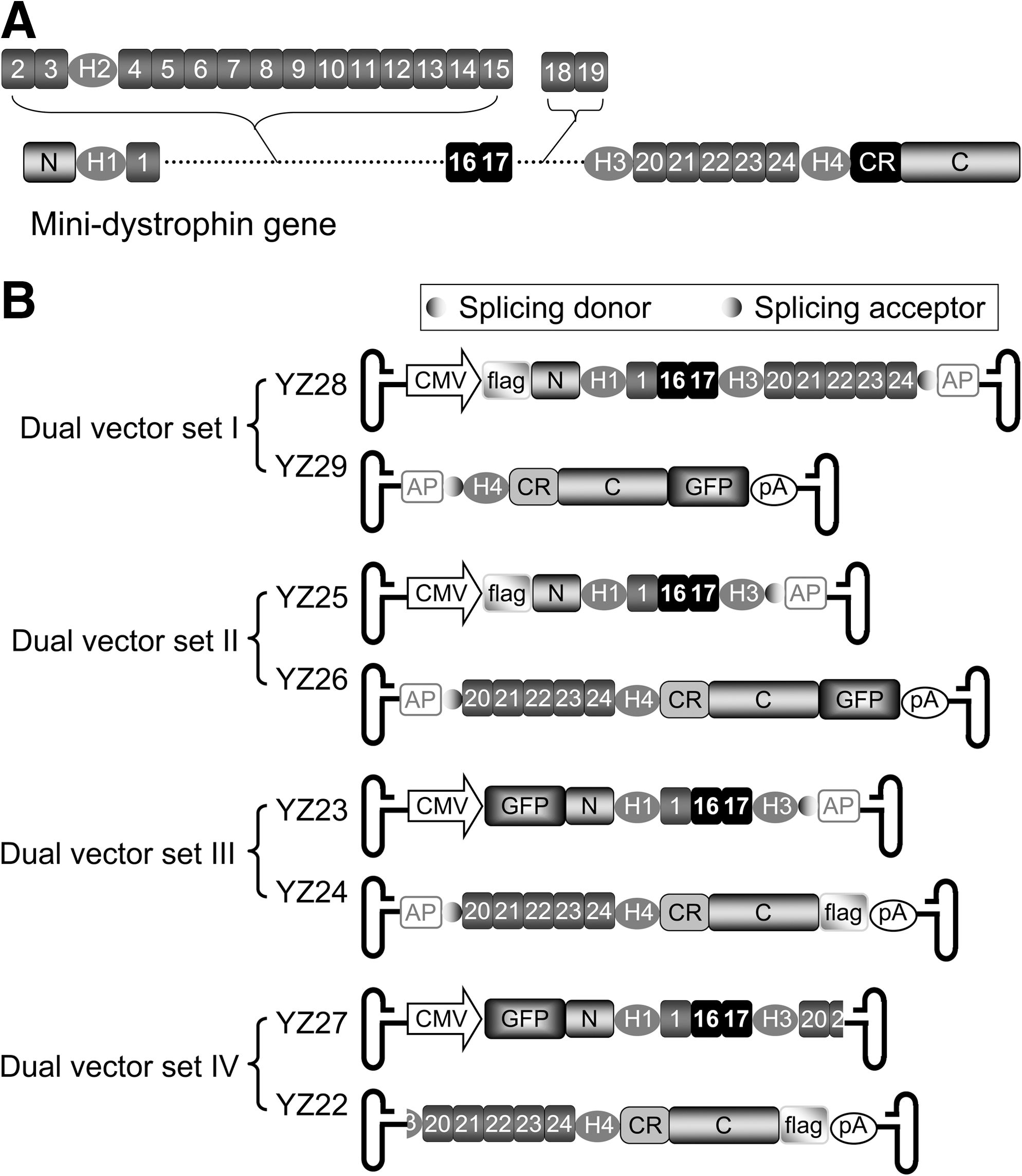
Schematic outline of the mini-dystrophin gene structure and dual adeno-associated virus (AAV) constructs.
Four different sets of the dual AAV vectors were designed to express the ΔR2–15/ΔR18–19 nNOS recruiting minigene (Fig. 1). To facilitate detection, a FLAG tag and a GFP gene were separately fused in frame to either the N-terminal or C-terminal end of the minigene (Fig. 1B) (Li et al., 2006; Rodino-Klapac et al., 2010). The CMV promoter and SV40 pA sequence were used for transcriptional regulation in all dual vectors. Dual vector sets I to III were hybrid dual AAV vectors in which an approximately 0.3-kb highly recombinogenic DNA fragment from the human placental alkaline phosphatase (AP) gene was used to assist interviral genome recombination (Ghosh et al., 2008, 2011). The synthetic intron splicing signals (a donor and an acceptor) were also engineered in each vector set. These splicing signals were used to remove the AP gene fragment and the viral inverted terminal repeat junction in the reconstituted viral genome in the hybrid vectors. Specifically, dual vector set I consisted of YZ28 and YZ29 (Fig. 1B). In this set, the ΔR2–15/ΔR18–19 minigene was split between spectrin-like repeat 24 and hinge 4. The FLAG tag was fused in front of the dystrophin N-terminal domain and the GFP was fused at the C-terminal end. The minigene was split between hinge 3 and spectrin-like repeat 20 in dual vector set II (YZ25 and YZ26) and III (YZ23 and YZ24). The difference between sets II and III was the location of the fused tags. In set II the FLAG tag was fused to the N-terminal end and the GFP gene was fused to the C-terminal end. In set III, the position of these tags was switched (Fig. 1B).
Dual vector set IV (YZ27 and YZ22) was built as the overlapping vectors (Duan et al., 2001; Ghosh and Duan 2007; Odom et al., 2011). Specifically, an approximately 0.4-kb region (including part of hinge 3, whole repeat 20, and part of repeat 21) was shared by both vectors (Fig. 1B). The GFP signal was engineered at the N-terminus in YZ27 and the FLAG tag was engineered at the C-terminus in YZ22 (Fig. 1B).
Dual vector set IV resulted in superior transduction in mdx4cv muscle
To compare the relative transduction efficiency of four pairs of dual vectors, we injected them to the anterior tibialis muscle of 4-week-old male mdx4cv mice (Fig. 2). Each set of vectors was delivered as pairs at a dose of 1.2×1010 vg particles/vector/muscle (n=5 muscles/vector set). Dual vector reconstitution was evaluated 4 weeks after gene transfer.
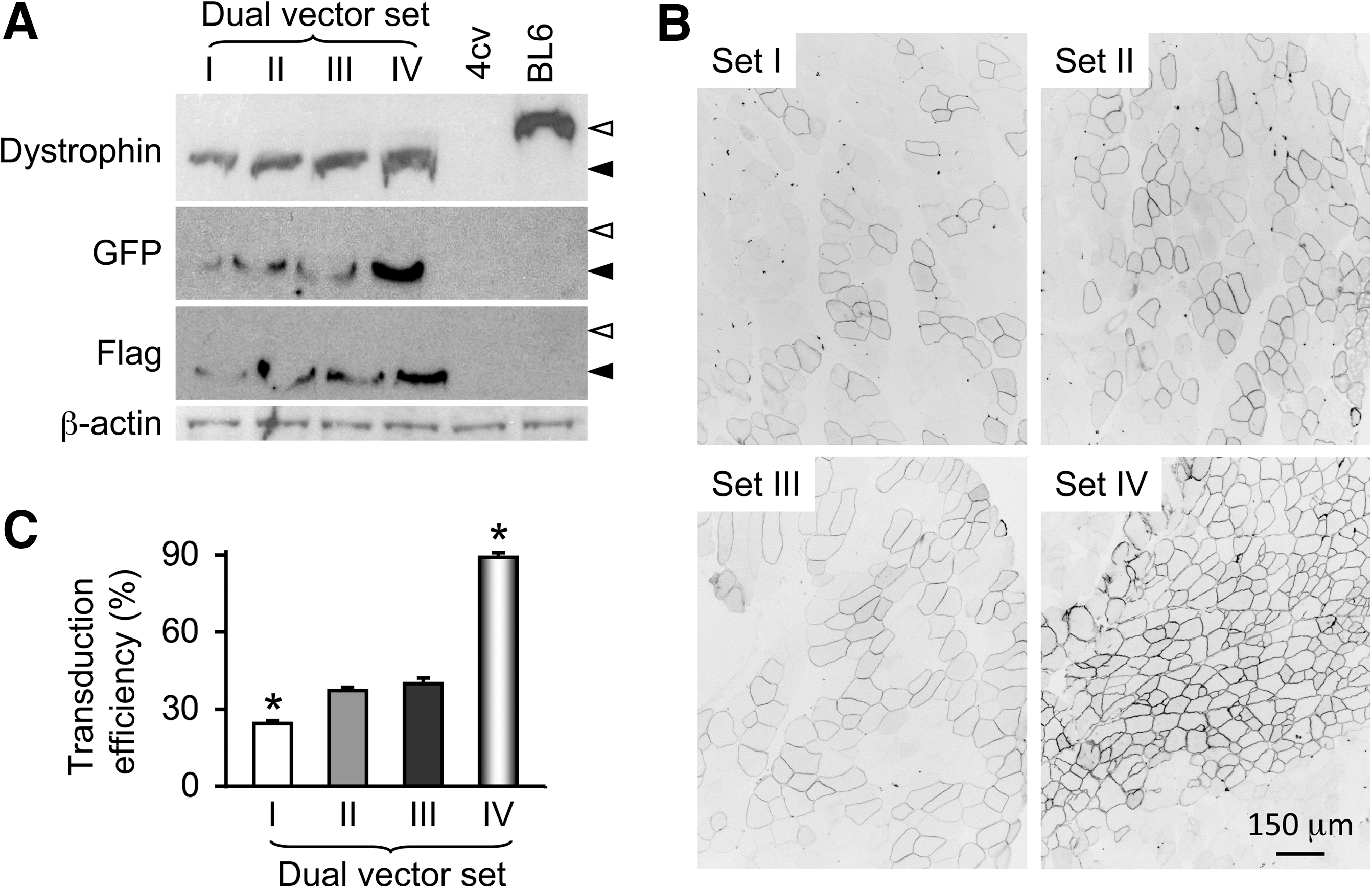
Comparative evaluation of four different sets of the mini-dystrophin dual AAV vectors in mdx4cv mice.
Western blot with a dystrophin C-terminal domain specific antibody revealed an approximately 250-kD band in all infected muscles (Fig. 2A). This is the expected size of the GFP/FLAG-fused mini-dystrophin protein. Expression of the fusion protein was further confirmed by Western blots using GFP-specific and FLAG-specific antibodies (Fig. 2A). Interestingly, substantial differences appeared in the expression level from each set of dual vectors. Set IV vectors (YZ27 and YZ22) yielded the strongest band and the set I vectors (YZ28 and YZ29) yielded the weakest band (Fig. 2A). Muscles infected with sets II (YZ25 and YZ26) and III (YZ23 and YZ24) showed bands of intermediate intensity (Fig. 2A).
To quantify transduction efficiency, we counted the percentage of GFP-positive myofibers (Fig. 2B, C). Consistent with the Western blot results, GFP quantification revealed a significantly higher transduction efficiency of the set IV vectors (YZ27 and YZ22) (Fig. 2C). The number of GFP-positive cells was significantly lower in muscles infected by set I vectors (YZ28 and YZ29) (Fig. 2B, C).
Dual vector–mediated mini-dystrophin expression restored membrane-associated nNOS expression
Serial muscle sections were examined for GFP, human mini-dystrophin, and nNOS expression. Figure 3 shows representative results from muscles infected with set IV vectors (YZ27 and YZ22). Direct GFP fluorescence imaging, indirect immunofluorescence staining with a human dystrophin monoclonal antibody and a polyclonal nNOS antibody all showed the same staining pattern (Fig. 3). In situ NOS activity staining further confirmed correct localization of nNOS on the sarcolemma.

Dual AAV vectors restored sarcolemmal nNOS expression. Left and middle panels, representative serial muscle cross-section photomicrographs from dual AAV vector set IV–infected mdx4cv muscle. Top left panel, GFP fluorescence image; top middle panel, indirect immunofluorescence staining with a human dystrophin-specific antibody; bottom left panel, immunofluorescence staining with a polyclonal nNOS antibody; bottom middle, a histochemistry image of NOS activity staining. Right panels, representative NOS activity staining in normal BL6 and dystrophin-null mdx4cv muscles. Scale bar applies to all images. Asterisk marks the same myofiber in serial sections. Black asterisk, a myofiber transduced by the dual AAV vector; gray asterisk, a myofiber that was not transduced by AAV.
Muscles infected by other three sets of dual vectors were also examined for GFP, mini-dystrophin, and nNOS expression. Although the overall transduction efficiency was significantly lower (Fig. 2), membrane-associated nNOS expression was confirmed in every case (data not shown).
Discussion
Membrane localization of nNOS plays an important role in DMD. In normal muscle, nNOS is activated to produce nitric oxide (NO) during contraction. Rapid diffusion of NO to the surrounding vasculature counteracts α-adrenergic vasoconstriction and prevents contraction-induced functional ischemia (Thomas et al., 1998; Grange et al., 2001; Lai et al., 2009). In dystrophin-deficient muscle, total cellular nNOS is decreased. Residual nNOS is mislocalized to the cytosol (Brenman et al., 1995; Li et al., 2011). Delocalized nNOS results in nitrosative stress and muscle force reduction (Li et al., 2011). For these reasons, normalizing nNOS expression has been considered as an important goal in the development of novel treatments for DMD.
Replacing the mutated dystrophin gene may lead to a breakthrough in DMD therapy. A startling advancement over the last decade is the development of abbreviated but yet functional synthetic dystrophin genes. The microgene and minigene are two types of the minimized dystrophin genes currently under evaluation. The microgene carries only ∼30% of the full-length coding sequence. The minigene carries more genetic information (50%–60% of the full-length coding sequence). However, it has been technically more challenging to deliver the minigene. For example, the microgene is less than 4 kb and can fit into a single AAV particle. However, the 6- to 8-kb minigene exceeds the AAV packaging capacity and requires dual vector technology for delivery (Lai et al., 2010).
Despite the methodology difficulties, our group and Dr. Jeffery Chamberlain's group have independently developed several different versions of dual AAV vectors to express the ΔH2-R19 minigene (Lai et al., 2005; Ghosh et al., 2008; Odom et al., 2011). The ΔH2-R19 minigene is an optimized version of a minigene originally derived from human patients (England et al., 1990; Harper et al., 2002). Lai et al. (2005) developed a set of the trans-splicing vectors by splitting the ΔH2-R19 minigene at the junction of exons 60 and 61 (Lai et al., 2005). The transduction efficiency of this dual vector set reached that of a single AAV vector (Lai et al., 2005). Ghosh et al. (2008) generated a set of the trans-splicing vectors and a set of the hybrid dual vectors by splitting the ΔH2-R19 minigene between exons 55 and 56. In the hybrid vectors, a 0.87-kb fragment from the middle section of the AP gene was used to enhance recombination. Interestingly, the hybrid set resulted in significantly higher transduction. Odom et al. (2011) achieved efficient reconstitution by dividing the ΔH2-R19 minigene into two fragments. R20 was shared in this set of the overlapping vector as the common sequence for homologous recombination.
Transgenic expression of the ΔH2-R19 minigene has resulted in 100% recovery of the specific muscle strength by in vitro force measurements (Harper et al., 2002). However, this minigene cannot recruit nNOS to the sarcolemma (Lai et al., 2005). To overcome this limitation, we have recently developed the second-generation minigenes that carry the nNOS binding domain (Lai et al., 2009). In this study, we designed and constructed four different sets of dual AAV vectors to express the nNOS-recruiting ΔR2-R15/ΔR18–19 minigene and we also compared their transduction efficiency in mdx4cv mice (Figs. 1 and 2). To simplify the screening process and validate transgene reconstitution, a GFP tag and a FLAG tag were engineered in-frame at either the 5′-end or the 3′-end of the ΔR2–15/R18–19 minigene (Fig. 1B). For efficient delivery, all vectors were packaged in the Y445F tyrosine mutant AAV-6 virus (Zhong et al., 2008; Qiao et al., 2010).
Among four sets of dual vectors, three sets (set I to III) were built using the synthetic splicing signals and a 0.3-kb AP gene fragment for transgene reconstitution (Ghosh et al., 2008, 2011). In the set IV vectors, the full expression cassette was expected to reconstitute via homologous recombination (Duan et al., 2001; Ghosh et al., 2006; Odom et al., 2011). While mini-dystrophin expression and sarcolemmal nNOS localization was detected in every dual vector infected muscle, set IV resulted in the highest transduction efficiency (Figs. 2 and 3). This is interesting but not surprising. We have previously demonstrated that dual AAV vector–mediated transduction is highly dependent on the vector design (Lai et al., 2005; Ghosh et al., 2006). In the case of the trans-splicing vectors, the location of the gene-splitting site appears to have a decisive effect. When an ideal site is chosen, one may achieve robust expression similar to that of a single AAV vector (Lai et al., 2005). In the case of the overlapping vectors, the exact sequence used for homologous recombination determines the outcome. A longer fragment may not necessarily lead to better reconstitution (Ghosh et al., 2011). The results here, as well as those published by Odom et al. (2011) suggest that certain regions of the dystrophin coding sequence are highly recombionogenic.
In summary, we reported for the first time the construction of a series of dual AAV mini-dystrophin vectors that restored sarcolemmal nNOS expression. We further demonstrated that the set IV (YZ27 and YZ22) was especially promising. Future physiological assays (such as muscle force, blood perfusion, and exercise capacity) in murine and canine models may further reveal the therapeutic benefit of the nNOS restoring mini-dystrophin gene dual AAV vectors.
Footnotes
Acknowledgments
The study was supported by grants from the National Institutes of Health (DD) AR-49419, NS-62934, HL-91883, and the Muscular Dystrophy Association (DD). We thank Thomas McDonald, Keqing Zhang, and Yongping Yue for excellent technical assistance. We thank Yi Lai for helpful discussion.
Author Disclosure Statement
No competing financial interests exist.