
Abstract
Libraries based on the insertion of random peptide ligands into the capsid of adeno-associated virus type 2 (AAV2) have been widely used to improve the efficiency and selectivity of the AAV vector system. However, so far only libraries of 7-mer peptide ligands have been inserted at one well-characterized capsid position. Here, we expanded the combinatorial AAV2 display system to a panel of novel AAV libraries, displaying peptides of 5, 7, 12, 19, or 26 amino acids in length at capsid position 588 or displaying 7-mer peptides at position 453, the most prominently exposed region of the viral capsid. Library selections on two unrelated cell types—human coronary artery endothelial cells and rat cardiomyoblasts—revealed the isolation of cell type–characteristic peptides of different lengths mediating strongly improved target-cell transduction, except for the 26-mer peptide ligands. Characterization of vector selectivity by transduction of nontarget cells and comparative gene-transduction analysis using a panel of 44 human tumor cell lines revealed that insertion of different-length peptides allows targeting of distinct cellular receptors for cell entry with similar efficiency, but with different selectivity. The application of such novel AAV2 libraries broadens the spectrum of targetable receptors by capsid-modified AAV vectors and provides the opportunity to choose the best suited targeting ligand for a certain application from a number of different candidates.
Introduction
Many in vitro and in vivo studies have shown that the 12 distinct human and nonhuman primate AAV serotypes (AAV1–12) sequenced to date (Atchison et al., 1965; Hoggan et al., 1966; Parks et al., 1967; Bantel-Schaal and zur Hausen, 1984; Rutledge et al., 1998; Gao et al., 2002, 2004; Mori et al., 2004; Schmidt et al., 2008a,b) exhibit different tissue and cell tropisms. Therefore, pseudotyping of AAV2 vector genomes with capsids of different AAV variants or the generation of mosaic capsids can enable recombinant AAV (rAAV) delivery to previously refractory cell types (Rabinowitz et al., 2004; Choi et al., 2005; Gigout et al., 2005; Wu et al., 2006). However, the use of pseudotyped or mosaic vectors per se does not imply a higher cell-type specificity nor does it help in overcoming preexisting immunity against the parental serotypes. Improved gene-transfer efficiency and specificity can be achieved through rational vector targeting, for example, via bispecific adaptor molecules or chemical modifications of the AAV capsid. Yet such vector modifications are often associated with reduced titers, decreased vector stability, or additional immunogenicity elicited by these conjugates (Bartlett et al., 1999; Ponnazhagan et al., 2002; Arnold et al., 2006). Direct targeting approaches, e.g., through genetic modifications of the viral capsid by incorporation of targeting ligands (Girod et al., 1999; Wu et al., 2000; Loiler et al., 2003), can circumvent these problems, but will always remain limited to the use of already known, defined targeting peptides.
A further step toward tailored viral vectors for gene transfer has been achieved by library-based approaches. In a first set of experiments, cell-binding motifs derived from peptide libraries displayed on the surface of M13 phages have been inserted preferentially in the threefold spike region of the AAV2 capsid or in other capsid positions tolerating peptide insertions (Grifman et al., 2001; Nicklin et al., 2001; Work et al., 2004, 2006). Although this approach was successful in some instances, insertion of cell type–specific binding peptides isolated by phage display into the AAV capsid changed the specificity and affinity of peptide binding in many cases, presumably by steric and electrostatic interactions with surrounding capsid protein structures. In addition, peptides derived from phage display libraries are selected exclusively for cell binding, but do not necessarily mediate efficient cell entry or intracellular processing of the vector. The development of AAV display peptide libraries overcame these problems by allowing the selection of targeting peptides directly in the context of the viral capsid (Müller et al., 2003; Perabo et al., 2003). Such library screenings not only imply an inherent selection for proper conformation of the peptide ligands within the capsid structure and efficient cell binding, but also warrant successful internalization and intracellular processing of selected library clones.
AAV2 display peptide libraries have been successfully applied for the selection of vectors both in vitro and in vivo, yielding unique peptide motifs that mediate enhanced (but not always specific) gene transfer to various cell types (Waterkamp et al., 2006; Michelfelder et al., 2007, 2009; Ying et al., 2010). More recently, the combination of AAV peptide display libraries with directed evolution approaches, based on the fragmentation and staggered resynthesis of cap genes from different AAV serotypes by error-prone PCR (Maheshri et al., 2006), were used to exploit the advantageous intracellular processing abilities of different AAV serotypes together with receptor targeting via inserted peptide ligands (Grimm et al., 2008).
In previous studies, random AAV2 display peptide libraries were generated by the incorporation of peptides of seven amino acids (aa) in length into the AAV2 capsid at aa positions 588 or 587 (Müller et al., 2003; Perabo et al., 2003; Waterkamp et al., 2006; Michelfelder et al., 2007, 2009; Ying et al., 2010), but thus far the optimal and maximal length of inserted peptides regarding structural constraints has not been determined. Here we describe the construction of libraries displaying peptides of diverse length, ranging from 5 to 26 aa in length at aa 588, or displaying a 7-mer peptide library at aa 453, the most prominently exposed position of the AAV2 capsid, which might impact both the accessibility for capsid–receptor interactions, as well as the kind of receptors amenable for binding. Selection of these libraries yielded divergent peptide ligands enabling superior transduction of otherwise refractory primary human coronary artery endothelial cells (HCAEC). Peptides of different length selected on the identical cell type demonstrated divergent degrees of transduction efficiency and specificity and enabled the targeting of different cellular receptors. Therefore, the reported approaches expand the capacity of the AAV display system for the selection of the optimally targeted gene-therapy vector and the best suited ligand length for a given application.
Materials and Methods
Cells and cell culture
H9C2 (rat cardiomyoblasts) HeLa and 293T cells were cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal calf serum (FCS), 2 mM L-glutamine, and 100 μg/ml penicillin/streptomycin. Primary HCAEC (PromoCell, Heidelberg, Germany) were maintained in Endothelial Cell Growth Medium MV supplemented with SupplementMix and 100 μg/ml penicillin/streptomycin. Human tumor cell lines from the NCI-60 panel were maintained in RPMI 1640 supplemented with 10% FCS, 2 mM L-glutamine, and 100 μg/ml penicillin/streptomycin. All cells were constantly kept in a humidified atmosphere at 37°C and 5% CO2.
Generation of AAV2 display peptide libraries
Random AAV2 display peptide libraries were produced in a three-step process as described earlier (Waterkamp et al., 2006). For the production of AAV2 libraries displaying peptides of diverse length, (NNK)n, random oligonucleotides encoding n=5-, 7-, 12-, 19-, or 26-aa inserts at nucleotide position 3,967 of the AAV2 genome were synthesized. The following degenerated oligonucleotides, flanked by two incompatible BglI restriction sites on each end, were used for directional in-frame cloning into the linearized library backbone plasmid pMT187-0-3-SfiI (Müller et al., 2003): 5′-CAGTCGGCCAGAGAGGC(NNK)nGCCCAGGCGGCTGACGAG-3′. To minimize the number of triplets theoretically encoding stop codons, the (NNK) insert was chosen in which “N” denotes the four nucleotides A, T, C, and G, and “K” stands for nucleotides G or T. The diversity of the resulting AAV2 plasmid libraries was determined by the number of individual colonies obtained from plating representative aliquots of transformed ElectroMAX electrocompetent bacteria (Invitrogen, Karlsruhe, Germany). The generated plasmid libraries were subsequently used for the production of AAV2 transfer shuttle and virus libraries.
For the generation of an AAV2 display library with a novel peptide insertion site at glycine 453 of the VP protein, a synthetic nucleotide sequence of 797 bp comprising the AAV2 cap gene sequence with two incompatible SfiI restriction sites at nucleotide position 3,559 was designed (GeneArt, Regensburg, Germany). This synthetic sequence was subsequently cloned via BsiWI and XcmI restriction into the AAV2 cap gene sequence of a pBluescript-based plasmid. The generated library backbone plasmid, pBS-ITRmut, contained the AAV2 genome flanked by inverted terminal repeats (ITRs) and two SfiI restriction sites separated by a 15-bp oligonucleotide stuffer to allow peptide insertions at aa position 453. Library production was carried out in a three-step process as described before (Waterkamp et al., 2006). Random oligonucleotides encoding a 7-aa insertion and flanked by incompatible BglI restriction sites, 5′-CAGTCGGCCCAAGTGGC(NNK)7GCCGGAACGGCTGACGAG-3′, were used for directional in-frame cloning into the linearized library backbone plasmid pBS-ITRmut-SfiI, and the resulting plasmid library was subsequently used for the production of AAV2 transfer shuttles and the final virus library. PCR amplification of virus DNA followed by agarose gel electrophoresis was applied to test all AAV2 transfer shuttle and virus libraries for contaminations by wild-type (wt) AAV2 using the following primer sets: 5′-GGTTCTCATCTTTGGGAAGCAAG-3′ and 5′-TGATGAGAATCTGTGGAGGAG-3′ (inserts at position 588) and 5′-GACGTCTTCATGGTGCC-3′ and 5′-GTGGTTCCACTTGGAG-3′ (inserts at position 453).
Sequencing of AAV2 libraries
AAV2 plasmid and virus libraries were analyzed by DNA sequencing to verify the presence of random oligonucleotide inserts and to analyze the aa distribution of the encoded peptide ligands. For this purpose, each AAV2 plasmid library, as well as viral DNA purified from each virus library (DNeasy Blood & Tissue kit; Qiagen, Hilden, Germany), was amplified by PCR using the distinct primer pairs 5′-ACCTCCAGAGAGGCCAGAGAG-3′/5′-CTGAAGGTACACATCTCTGTC-3′ and 5′-TACTTAAGCAGAACAAACGGC-3′/5′-CTGAAGCCTTGACTGCGTGGC-3′ for the analysis of inserts at aa position 588 and 453, respectively. The PCR products were subsequently ligated into vector pCR2.1-TOPO using TOPO TA Cloning Kit (Invitrogen) and transformed in chemical competent DH5α bacteria. Stab cultures of single transformants were analyzed by high-throughput sequencing using primer M13R (GATC Biotech, Konstanz, Germany).
Selection of AAV2 random display peptide libraries
For screening, 2.5×106 H9C2 cells or 1×106 HCAEC were infected with AAV2 library particles at multiplicities of infection (MOIs) of 1,000 or 10,000 capsids/cell, respectively, in the first selection round. After 4–6 hr of incubation at 37°C, cells were washed and superinfected with adenovirus serotype 5 (Ad5) at 5,000 plaque-forming units (pfu)/cell (H9C2) or 20 pfu/cell (HCAEC) to enable AAV replication. Successfully infecting and replicating AAV2 library clones were harvested from target cells after 48–72 hr by three cycles of repeated freezing and thawing and titrated by AAV2 capsid ELISA (Progen, Heidelberg, Germany). Recovered, preselected viruses were applied to freshly seeded cells at 50% lower MOI in each subsequent selection round. Library coselection was performed using a mixture of equal amounts of capsid particles from each library (e.g., 2,000 capsids/cell of each library displaying peptides of 5, 7, 12, 19, or 26 aa in length for selection on HCAEC) in the first selection round. After each selection round, viral DNA was extracted from cell lysates using DNeasy Blood & Tissue kit (Qiagen). Viral DNA comprising the respective library insert region of the cap gene was amplified by PCR as described above. PCR products were ligated into pCR2.1-TOPO (Invitrogen) and sequenced using the standard primer M13R.
Production of AAV2 reporter vectors
rAAV2 vectors displaying a selected peptide motif on the capsid surface and harboring a luciferase (Luc)- or β-galactosidase (LacZ)-encoding reporter gene cassette were used for gene-transduction analysis of selected library clones. Recombinant reporter vectors were produced by triple transfection of 293T cells with (a) the respective cap gene-modified plasmids, pMT187xx2-“insert” or pMT187xx2mut-“insert”, encoding peptide insertions at aa position 588 or 453 of the VP protein but lacking ITR sequences necessary for genome packaging, (b) pDGΔVP (Dubielzig et al., 1999) providing adeno helper genes, and (c) pTR-UF2-CMV-Luc or pTR-UF2-CMV-LacZ, derivatives of the plasmid pUF2 (Zolotukhin et al., 1996) harboring the luciferase or β-galactosidase genes under control of the cytomegalovirus (CMV) promoter and flanked by ITRs for packaging.
Luciferase reporter vectors comprising wt AAV2 capsids were generated by double transfection of (a) pTR-UF2-CMV-Luc, together with (b) the helper plasmid pDP2 (Grimm et al., 2003). Transfections were performed by calcium phosphate precipitation, and vector productions were purified by iodixanol density-gradient centrifugation (Hauswirth et al., 2000).
Titration of AAV library and vector productions
Titers of assembled AAV2 capsid particles (capsids per milliliter) were determined as previously described (Grimm et al., 1999) using an A20 antibody-based AAV2 Titration ELISA (Progen). Genomic titers of AAV2 library and vector productions were determined as the number of viral genome-containing particles (vg/ml) by quantitative real-time PCR as described earlier (Veldwijk et al., 2002). The following combinations of forward and reverse primers, as well as the respective fluorescently labeled probes diluted in TaqMan Universal PCR Mastermix (Applied Biosystems, Darmstadt, Germany), were used: detection of rep: 5′-AAGTCCTCGGCCCAGATAGAC-3′ (rep+), 5′-CAATCACGGCGCACATGT-3′ (rep–), and 6-fam-TGATCGTCACCTCCAACA-MGB (probe); detection of CMV enhancer: 5′-TGCCCAGTACATGACCTTATGG-3′ (cmv+), 5′-GAAATCCCCGTGAGTCAAACC-3′ (cmv–), and 6-fam-AGTCATCGCTATTACCATGG-MGB (probe). The number of infectious, replication-competent AAV particles was determined by a 10-fold serial endpoint titration on HeLa cells, as previously described (Grimm et al., 1999).
Luciferase reporter gene assays
Gene transduction by selected rAAV2 was analyzed by means of luciferase reporter gene expression in target and nontarget cells. For this purpose, 5×103 cells were seeded in each well of a 96-well plate. After 24 hr, cells were transduced with rAAV2-Luc vectors at MOIs of 10,000 vg/cell (HCAEC, H9C2) or 1,000 vg/cell (HeLa) and incubated at 37°C in serum-reduced (5% FCS) culture medium for 96 hr. For competition experiments, HCAEC were transduced with 5,000 genome-containing rAAV2-Luc vector particles per cell for 24 hr in the presence of a 25-fold excess of rAAV2-LacZ vectors, followed by a 24-hr incubation period in complete medium containing no vector particles. Cells were washed with PBS, resuspended in reporter lysis buffer RLB (Promega, Mannheim, Germany), and lysed by incubation overnight at –80°C followed by thawing at 37°C. Reporter gene activity was analyzed in a Lumat LB 9501 luminometer (Berthold, Bad Wildbad, Germany) using Luciferase Assay System substrate (Promega) according to the manufacturer's instructions.
Comparative gene-transduction analysis
The gene-transduction profiles of rAAV2 vectors displaying HCAEC-selected peptides of different length were compared with gene-expression data from cDNA microarrays developed for the NCI-60 panel of human tumor cell lines (Ross et al., 2000) in order to analyze how gene transfer is influenced by peptide modifications of diverse length displayed on the AAV2 capsid. For transduction experiments, 44 adherent tumor cell lines derived from diverse tissues, including brain, blood, bone marrow, breast, colon, kidney, lung, ovary, prostate, and skin, were used. Gene-expression data for these cell lines are available from the Developmental Therapeutics Program (DTP) at the National Cancer Institute Web site (
Statistical analysis
All data are expressed as means±SD. To test for statistical significance, an unpaired Student's t test was applied. Data were considered significant when p<0.05.
Results
Generation of novel AAV2 libraries displaying peptides of diverse lengths and at different capsid positions
A successful screening system for targeted AAV2 vectors by displaying peptides directly on the AAV2 capsid has been developed based on the combination of two critical parameters: the insertion site on the capsid and the length of the displayed peptides. To explore further the potential of the combinatorial AAV display peptide system, a panel of novel AAV libraries displaying peptides of diverse length at position 588 of the capsid were designed and generated (Fig. 1a) using the previously validated three-step protocol consisting of the generation of a plasmid library, a transfer shuttle library, and the final virus display library. In addition, a 7-mer peptide library was inserted at aa position 453 at the peak of the threefold spikes of the AAV2 capsid (Xie et al., 2002) in order to test the general suitability of this capsid position for the selection of targeting ligands. Peptides inserted at this position (Fig. 1b; cyan balls) are arranged in a similar way, but with larger distance to each other, and protrude farther from the capsid surface than peptides inserted at position 588 (Fig. 1b; purple balls). All generated libraries were subsequently characterized and compared with the previously described AAV2 library (7-aa insert at position 588).

Design and characteristics of the novel AAV2 display peptide library constructs.
In a first step, AAV2 plasmid libraries with oligonucleotide insertions coding for 5, 7, 12, 19, or 26 aa in length—hereafter referred to as (NNK)5, (NNK)7, (NNK)12, (NNK)19, and (NNK)26—were generated by directional in-frame ligation of the respective oligonucleotide libraries. The resulting diversities of all newly generated plasmid libraries carrying oligonucleotide insertions of diverse length or at position 453 were in the range of 108 independent clones per library (Supplementary Table S1; Supplementary Data are available online at
Analysis of capsid assembly and genome encapsidation revealed that similar amounts of capsids and genome-containing particles were generated independent of the different peptide insertions or the different insertion site. However, genome encapsidation might have been slightly impaired in the case of the (NNK)5 transfer shuttle library (Fig. 2a). The number of replication-competent transfer shuttle particles was determined via an end-point titration replication assay (data not shown) and served as a measure for the amount of infectious particles used for the production of the final virus libraries. To produce the random AAV2 peptide display libraries, 293T cells were infected with the respective transfer shuttle library at an MOI of 1 replicative particle per cell to ensure uptake and propagation of approximately 1 library genome per cell, followed by superinfection with Ad5 at 10 pfu/cell. Again, PCR analysis demonstrated that the resulting AAV2 libraries were free of wt AAV2 particles (data not shown).
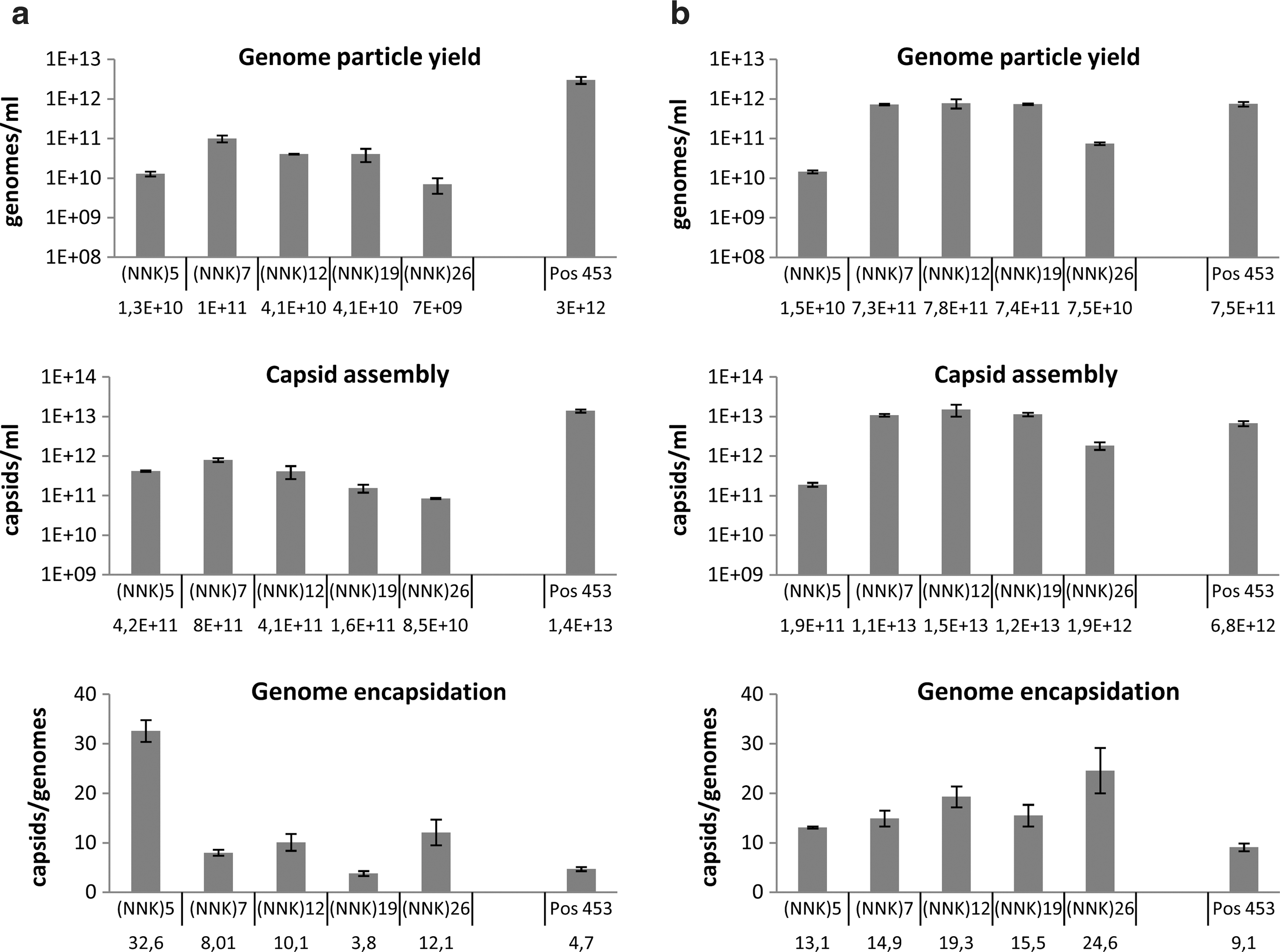
Characterization of capsid assembly and genome encapsidation. Analysis of the effects of peptide inserts of diverse length or at a novel position of the capsid protein (glycine 453) on virus particle yield and genome encapsidation of
All novel AAV2 display peptide libraries could be produced at titers sufficient for in vitro applications, and comparable yields of genome-containing virus particles and equivalent amounts of assembled capsids were obtained with capsid proteins containing the respective 7-, 12-, or 19-aa peptide insertions or the library inserted at glycine 453. In comparison, the yield of capsids and genome-containing particles was slightly reduced for the (NNK)26 library and even more for the (NNK)5 library. Despite the observed variations in the amounts of generated viral particles, the efficiency of genome encapsidation was not dramatically changed by peptide insertions of diverse length or at the different capsid position.
Analysis of the aa frequencies in the generated libraries
Up to 100 randomly assigned clones from each plasmid library were sequenced to verify the presence of random oligonucleotide inserts (at the plasmid level). DNA sequencing showed that different peptides were encoded in each clone derived from the respective plasmid library and that the frequency of aa encoded in these sequences was in accordance with the theoretical distribution expected for the (NNK) plasmid libraries (data not shown). Likewise, approximately 100 clones of each respective virus library were sequenced, which demonstrated a high degree of correlation between the aa frequencies in the individual peptide libraries and the theoretically predicted ones (Fig. 3). However, all libraries showed an underrepresentation of cysteine residues. In addition, a slight underrepresentation of phenylalanines and tryptophans, as well as lysine residues, was observed for several of the different-length libraries and the 453 library, respectively. Sequencing of the 26-mer library revealed the repeated presence of identical peptide ligands accompanied by an elevated representation of aspartic acid and asparagines, which points to a hampered incorporation of peptide ligands of this size into the AAV2 capsid.
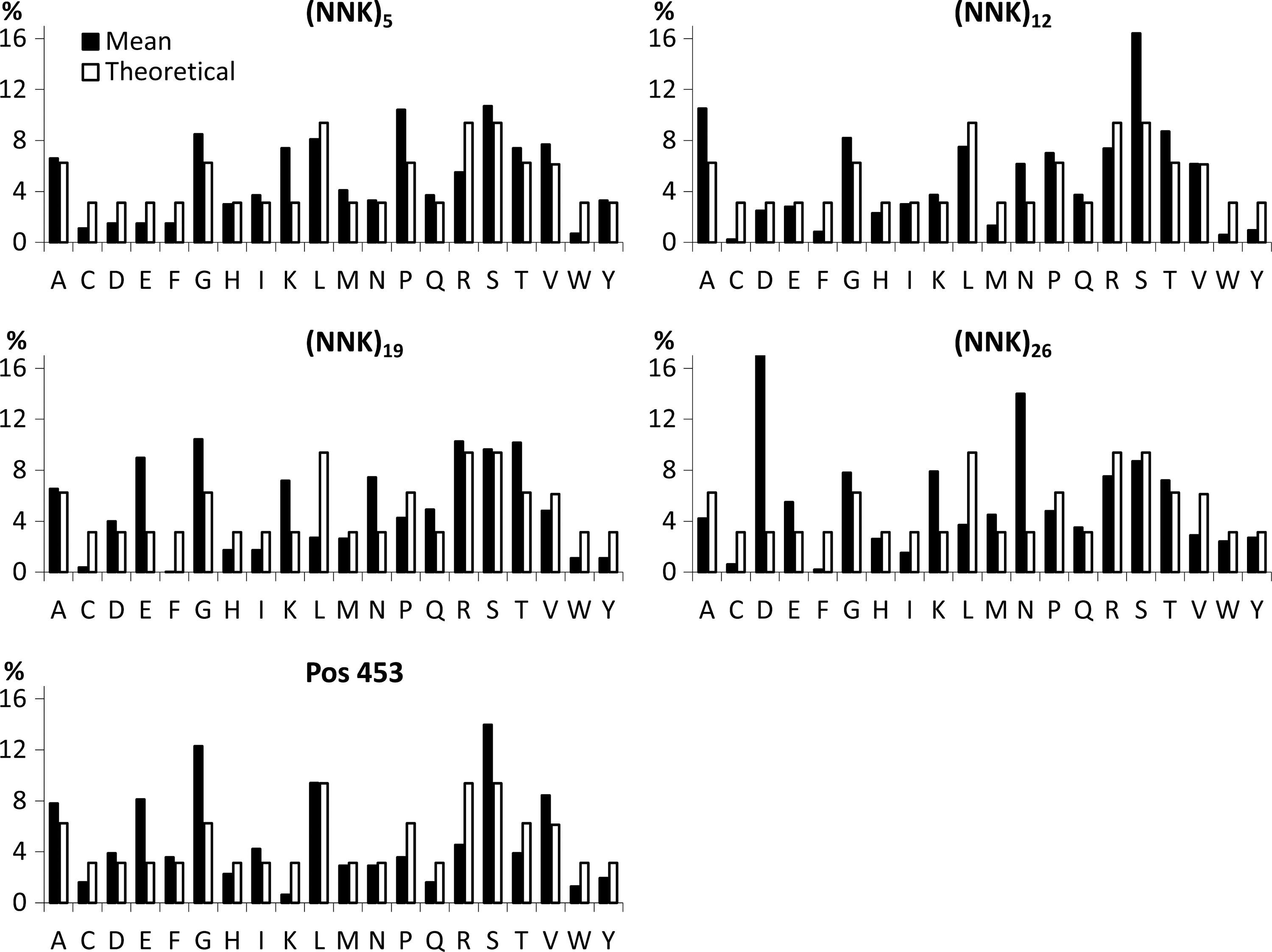
Amino acid frequencies in the generated AAV2 display peptide libraries. The sequences of approximately 100 randomly picked clones from each virus library were analyzed and evaluated regarding the frequencies (%) of aa encoded by the library inserts (black bars) in comparison with the theoretically predicted distribution of aa encoded by the respective (NNK)n oligonucleotide library (white bars). Amino acids are alphabetically ordered and displayed in one-letter code.
Selection of targeted viruses from AAV2 libraries displaying peptides of different length
To evaluate the potential of AAV2 display peptide libraries presenting peptides of different length for the selection of targeted vectors, in vitro screening was performed on two cell types of interest: HCAEC and rat cardiomyoblasts (H9C2). Viruses were subjected to repeated rounds of selection using a serial lower MOI as described previously (Waterkamp et al., 2006). DNA sequencing of virus clones selected on HCAEC revealed an enrichment of specific peptide motifs concomitant with a progressive reduction of the heterogeneity of the selected capsid mutants over four to six selection rounds with each of the libraries (Table 1). Selection of the (NNK)5 library on HCAEC resulted in the predominant isolation of one specific clone, DIIRA, with 75% of all recovered and sequenced clones after five rounds of selection. Therefore, a sixth round of selection was not performed. The (NNK)7 library has been omitted from this screening, because this library had been previously used for selection on HCAEC and the corresponding venous endothelial cells (Müller et al., 2003; Waterkamp et al., 2006). Screening of the (NNK)12 library resulted in enrichment of particular sequences, e.g., PENSVRRYGLEE, SYENVASRRPEG, and YSGQASPRTANT, which showed an increased frequency of recovery over five to six selection rounds (Table 1). Similarly, specific peptide inserts were frequently recovered after six rounds of screening with the (NNK)19 library. The most striking peptide motif was NDVWNRDNSSKRGGTTEAS, which showed a continuous enrichment between the fourth and sixth round (Table 1). Surprisingly, selection of the longest display peptide library, (NNK)26, led to the recovery of merely three independent clones, among which the clone YSPNSTGTNATRRKPEYNSTGADGNP was highly enriched with 85% and 96% recovery after the fourth and fifth selection round, respectively (Table 1).
Peptide ligands were obtained from individual selections of (NNK)5, (NNK)12, (NNK)19, and (NNK)26 libraries on HCAEC and from coselection using a mixture containing equal amounts of library capsid particles of each of the different-length peptide display libraries, including the previously described (NNK)7 library. HCAEC were infected with 10,000 capsids/cell in the first round and at serial lower MOIs in the subsequent screening rounds. Peptide sequences are displayed in one-letter code together with their frequency of recovery (%) after the indicated selection rounds. The numbers in brackets approximate the absolute count of repeatedly recovered library clones from a total number of ∼40 clones sequenced after each selection round.
N/A, not analyzed; —, not found.
To identify peptides possibly contributing to a superior ligand-directed gene transfer, library coselection was performed. For this purpose, cells were infected as described above, but using a mixture containing equal amounts of library capsid particles from each of the five different peptide display libraries, including an (NNK)7 library. Analysis of the peptide inserts encoded by the recovered clones showed that the most frequently isolated clone from the individual selection of the (NNK)5 library, DIIRA, was again highly enriched upon library coselection over six screening rounds. A 7-aa peptide motif, KGEPLSP, also demonstrated a marked frequency of recovery (40% of all recovered and sequenced clones) after six rounds and resembled a motif previously not isolated from the selection of this library on the identical cell type (Müller et al., 2003; Waterkamp et al., 2006). It is remarkable that the majority of relevant peptide motifs recovered from coselection were also those frequently enriched from individual selections of the respective single libraries. However, viruses with shorter peptide insertions (e.g., 5, 7 or 12 aa in length) were more frequently isolated by coselection than viruses with insertions of 19 or 26 aa.
To validate that the frequent recovery of particular peptide motifs from the selection on HCAEC was the result of specific ligand-mediated receptor targeting of the cells, additional library selections were carried out on the unrelated rat cardiomyoblast cell line H9C2. Successfully infecting library clones could be amplified and enriched after repeated rounds of selection on H9C2 (Table 2). Sequence analysis revealed that the H9C2-selected library clones displayed completely different peptide motifs than viruses recovered from screenings on HCAEC. In contrast to screening on HCAEC, selection of the (NNK)5 library on H9C2 resulted only in a very weak enrichment of specific peptide motifs. Yet individual screenings of the (NNK)7, (NNK)12, (NNK)19, and (NNK)26 libraries on H9C2 led to the highly frequent isolation of a number of viruses displaying distinct peptide ligands, most strikingly LSPVRPG, GARPSEVTTRPG, KMRPGAMGTTGEGTRVTRE, and WFIGDSIEVHQQGRDADSHMQGNRGG, which demonstrated a continuous increase in their frequency of isolation from round to round. The presence of an identical peptide module, RPG, in most of the predominantly enriched peptide ligands selected from different libraries thus seems remarkable. Interestingly, coselection of all libraries on H9C2 resulted in a prominent selection of 12-aa peptide sequences that were also frequently isolated by individual screening of the (NNK)12 library (Table 2). In addition, library coselection yielded very low frequencies of several 5-aa and 7-aa motifs, which were not isolated after individual library selections on H9C2 (data not shown).
Peptide ligands were obtained from individual selections of (NNK)5, (NNK)7, (NNK)12, (NNK)19, and (NNK)26 libraries on H9C2 cells and from coselection using a mixture containing equal amounts of library capsid particles of each of the different-length peptide display libraries. H9C2 cells were infected with 1,000 capsids/cell in the first round and at serial lower MOIs in subsequent screening rounds. Peptide sequences are displayed in one-letter code together with their frequency of recovery (%) after the indicated selection rounds. The numbers in brackets approximate the absolute count of repeatedly recovered library clones from a total number of ∼40 clones sequenced after each selection round. Bold letters highlight the repeatedly isolated three aa peptide motif RPG.
N/A, not analyzed; —, not found.
Selection of targeted viruses from AAV2 libraries displaying peptides at position 453
The feasibility of the application of an AAV2 library with random peptides inserted at position 453 was validated by in vitro screening over four rounds on HCAEC. Selection of this library yielded the recovery of a number of different library clones and only a slight enrichment of particular peptide ligands (Table 3).
Peptide ligands were obtained from selection of an (NNK)7 library inserted at capsid position 453 of AAV2 on HCAEC. Peptide sequences are displayed in one-letter code together with their frequency of recovery (%) after the fourth selection round. The numbers in brackets approximate the absolute count of repeatedly recovered library clones from a total number of 42 sequenced clones.
N/A, not analyzed.
Characterization of gene transduction by vectors displaying selected targeting peptides
AAV2 reporter vectors expressing the luciferase gene under control of the CMV promoter were used to investigate the efficiency and specificity of gene transfer mediated by peptide-modified AAV2 vectors selected on HCAEC or H9C2 cells. Recombinant wt AAV2 and a vector displaying a randomly chosen peptide (TEWDQPF) from an unselected library served as controls. Reporter vectors displaying the most frequently isolated peptides were chosen for analysis and could be successfully produced with titers ranging from 1.6×1010 to 1.7×1011 vg/ml (data not shown). Notably, even the insertion of longer motifs, such as 12-, 19-, or 26-mer peptides, led to a sufficient production of genome-containing rAAV2 vectors.
To determine the gene-transfer efficiency, HCAEC were transduced with 10,000 genome-containing vector particles per cell. Selected clones derived from the (NNK)5, (NNK)7, (NNK)12, or (NNK)19 libraries showed a significantly higher reporter gene expression compared with nontargeted vectors. The highest transduction efficiencies were achieved with vectors displaying the selected peptides NDVWN- (19-mer), DIIRA (5-mer), or PENSV- (12-mer), which mediated a 62–72-fold enhanced transduction of target cells compared with the wt AAV2 vector, and a 120–144-fold increased transduction compared with the nonselected random vector (Fig. 4a). Remarkably, these clones were also the ones progressively enriched through rounds of respective individual selections and also upon library coselection (Table 1). The YSPNS- (26-mer) clone, recovered with impressive frequencies from individual selection of the (NNK)26 library, surprisingly failed to transduce HCAEC in all experiments.

Characterization of gene transduction by vectors displaying selected peptide ligands
To evaluate the specificity of capsid mutants selected on HCAEC, gene transduction of nontarget HeLa cells using these clones was investigated by infection at an MOI of 1,000 vg/cell. In all cases, transduction with the selected clones resulted in a significant (73–97%) decrease in reporter gene activities compared with the wt AAV2 vector. The selected 26-mer clone YSPNS- again showed no transduction. A targeting index was calculated by multiplying the fold improvement of reporter gene expression in target cells with the fold decrease of reporter gene expression in HeLa cells as a measure for the retargeting efficacy achieved with the selected vectors (Supplementary Table S2). The highest retargeting efficacy was achieved with the 19-aa peptides NDVWN- and NRTYS-, followed by the 5-aa peptide DIIRA and the 12-aa peptide PENSV-.
Transduction of HCAEC with vectors displaying the selected peptide ligands MNVRGDL or ENVRGDL at position 453 of the viral capsid revealed a significant improvement of gene transfer compared with recombinant wt AAV2 (Fig. 4b). Despite the overall lower absolute levels of reporter gene activity in comparison with vectors displaying selected peptides at position 588, these vectors mediated a 58–89% decrease in luciferase expression in HeLa cells, resulting in modest retargeting efficacies (Supplementary Table S3). Furthermore, the efficient transduction of HeLa cells with vectors displaying the randomly chosen peptide ligand TEWDQPF at position 453 clearly indicated that the wt tropism of AAV2 is hardly affected by peptide insertions at this novel position in contrast to insertions at position 588.
Transduction analysis of H9C2 cells demonstrated a significantly higher degree of reporter gene expression mediated by all H9C2-selected clones displaying peptides of 7, 12, or 19 aa in length compared with the nontargeted wt vector, with the exception of vectors displaying the 12-mer peptide GTAMV- (Fig. 4c). The 12-mer peptides GARPS-, GNEVL-, and MSSDP-, derived from both individual and coselections, conferred a similar increase in gene-transduction efficiency as the 19-mer and 7-mer clones KMRPG- and GQHPR- recovered solely from individual selections of the respective libraries. The highest targeted transduction efficiency was achieved by presentation of the peptide LSPVRPG selected from the (NNK)7 library. The overall improvement of gene-transduction efficiency and specificity for H9C2 cells was, however, not as strong as for HCAEC. Although all H9C2-selected clones showed significantly reduced reporter gene expression in HeLa cells, the decrease in gene transfer mediated by vectors displaying the peptides LSPVRPG (7-mer), GARPS-, and KMPRG- (19-mer) was only in the range of 24–32% compared with wt vectors. This resulted also in clearly lower retargeting efficacies of these clones (Supplementary Table S4) in comparison with the HCAEC-selected peptides of diverse lengths or at capsid position 453.
Cross-transduction experiments, in which the transduction of HCAEC or H9C2 cells with vectors selected on the respective other cell line was evaluated, confirmed the overall superior specificity of gene transfer mediated by HCAEC-selected clones compared with vectors displaying H9C2-selected peptide motifs (Supplementary Fig. S1).
In conclusion, the selection of peptide libraries of diverse lengths or at an alternative capsid position yielded superior clones for targeted transduction of HCAEC, a cell type barely susceptible to wt vectors, and a moderate retargeting to H9C2, a second, unrelated cell type.
Comparative gene-transduction analysis of selected peptide-modified vectors on a large panel of human tumor cell lines
The divergence among HCAEC-selected peptide ligands regarding their aa sequence and the differential transduction of nontarget cells mediated by these peptides strongly suggested the interaction of these clones with different cellular proteins. To analyze this, transduction of 44 tumor cell lines with four HCAEC-selected vectors with different-length peptide insertions—DIIRA, KGEPLSP, SYENVASRRPEG, and NDVWNRDNSSKRGGTTEAS—was compared with the transduction achieved with wt AAV2 vectors and the vector displaying a randomly chosen peptide insert, TEWDQPF (Fig. 5a). The presentation of selected peptide ligands on the AAV2 capsid resulted in abundant alterations of the cell-transduction patterns, both in the absolute levels of reporter gene activity and in the relative permissiveness of the tumor cells to particular vectors. The transduction profiles obtained with HCAEC-selected vectors not only differed from gene transduction with wt AAV2 vectors and from the vector with the randomly chosen peptide insert, but also showed differences among each other. This further indicated that the selected vectors were bona fide retargeted to different cellular receptors and internalization pathways compared with the parental wt AAV2 vector. The COMPARE algorithm (
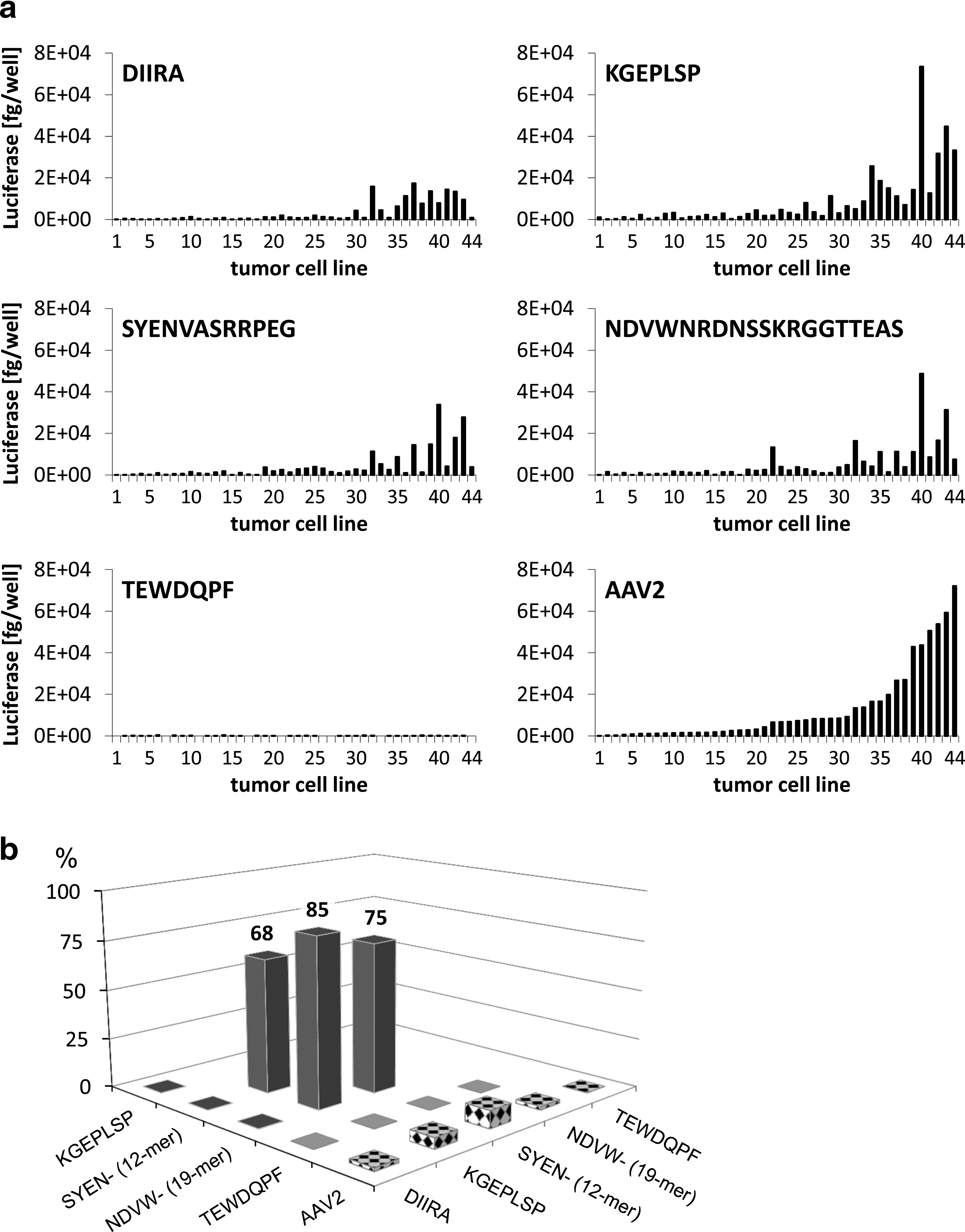
Comparative gene-transduction analysis using rAAV2 vectors displaying peptides of diverse length.
Analysis of receptor targeting through transduction competition experiments
In addition to comparative gene-transduction analysis, competition experiments were carried out using rAAV2 vectors displaying selected peptides of different length and harboring either a luciferase (Luc; used for the determination of transgene expression) or a β-galactosidase (LacZ; used for competition) reporter gene cassette. In the case of transduction via identical cellular receptors, coincubation of a Luc vector with a LacZ vector supplied in 25-fold excess should reduce the reporter gene activity obtained from the luciferase transgene. Transduction achieved with the respective luciferase reporter vectors in the absence of potentially competing vector capsids was set to 100%. As shown in Fig. 6, incubation of rAAV2-Luc vectors displaying the HCAEC-selected peptides DIIRA, KGEPLSP, SYEN- (12-mer), or NDVW- (19-mer) in the presence of an excess of LacZ vectors carrying the identical peptide ligand (“self-competition,” e.g., between DIIRA/DIIRA, KGEP-/KGEP-, …) led to a 60–72% reduction in luciferase activity. In contrast, an excess of vectors displaying the nonselected, randomly chosen ligand TEWDQPF resulted in a reduction of the luciferase signal of less than 13%, confirming the suitability of the assay, in which a decrease in the luciferase activity of more than 50% was considered significant.
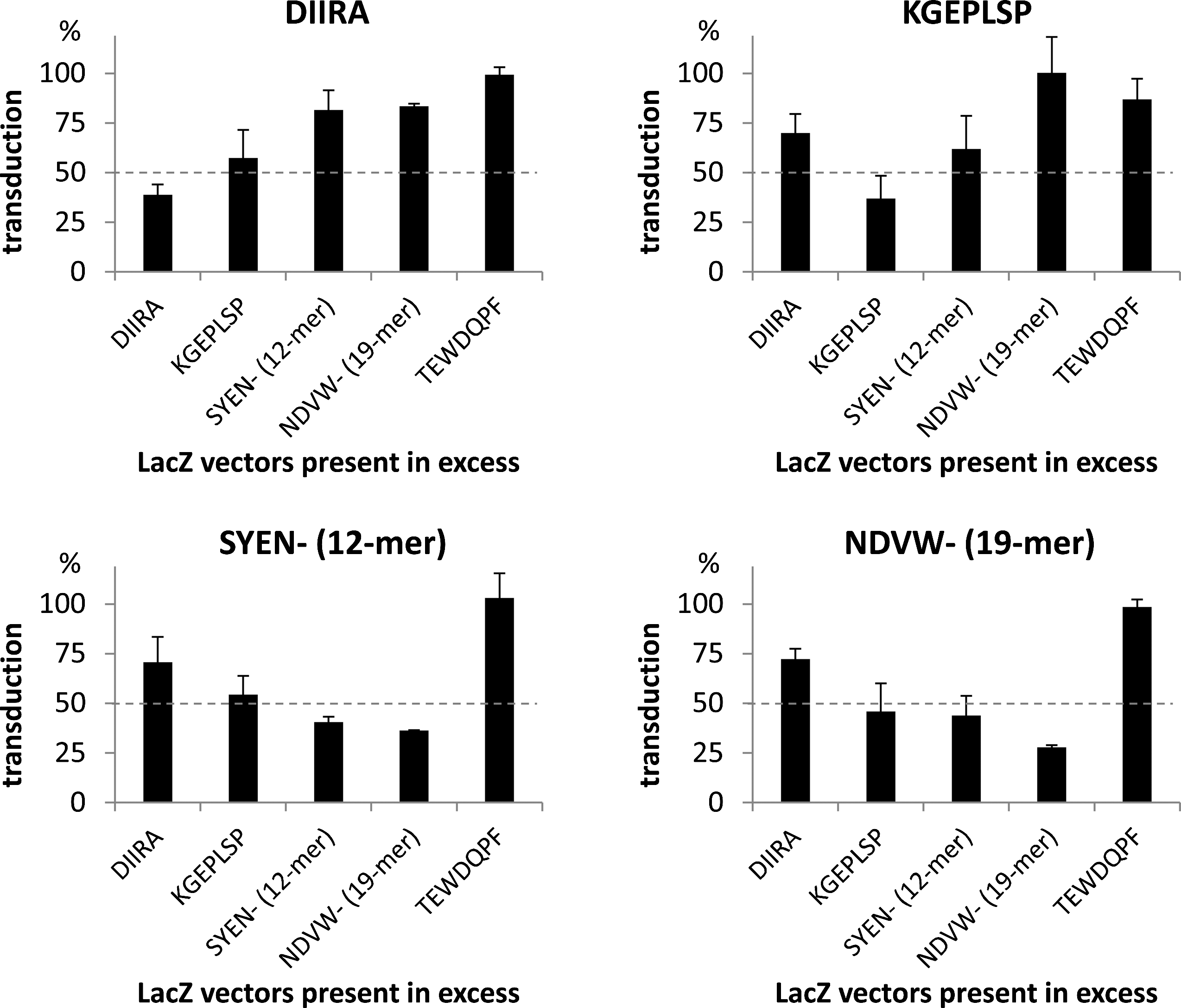
Modulation of cell transduction by rAAV2 luciferase vectors displaying peptides of diverse length in the presence of competing vector capsids. HCAEC were transduced with rAAV2 luciferase (Luc) vectors displaying the selected peptide ligands DIIRA (5-mer), KGEPLSP (7-mer), SYEN- (12-mer), or NDVW- (19-mer) at an MOI of 5,000 genome-containing particles per cell, in the presence of a 25-fold excess of vectors harboring β-galactosidase (LacZ) as a transgene and displaying the identical HCAEC-selected peptide motifs of diverse length or the randomly chosen ligand TEWDQPF. The vector solution was exchanged for fresh culture medium at 24 hr post infection. Luciferase activities were determined as relative light units (RLU) per well after 48 hr and are displayed as the proportion (%) of reporter gene activity achieved in the absence of competitors. Values are displayed as means±SD of at least 24 assays from three independent experiments. Peptide sequences are displayed in one-letter code. A reduction in reporter gene activity of more than 50% was considered significant and is indicated by a dotted line.
Besides self-competition between the respective vectors, transduction by rAAV2-Luc DIIRA was partially affected by coincubation with the vector displaying the 7-mer ligand KGEPLSP (43% reduction), whereas the presence of vectors displaying the longer 12- and 19-mer peptides only led to a slight reduction of the luciferase activity of less than 19%. In accordance with these results, rAAV2-LacZ vectors displaying DIIRA were only capable of a moderate ≤30% inhibition of cell transduction by rAAV2-Luc vectors carrying the 7-, 12-, and 19-mer peptides in the opposite experiments, indicating that transduction of the vectors with longer peptide insertions does not occur via the same receptors as transduction via the 5-mer ligand DIIRA. In contrast to this, transduction by rAAV2-Luc SYEN- (12-mer) as well as NDVW- (19-mer) was strongly inhibited by vectors displaying the 7-, 12-, and 19-mer motifs, indicating an overlapping receptor repertoire for these ligands. HCAEC transduction by the rAAV2-Luc KGEPLSP vector showed a more diverse pattern, in which rAAV2-LacZ SYEN- (12-mer) and DIIRA were capable of partial competition, whereas coincubation with rAAV2-LacZ NDVW- (19-mer) did not result in reduced luciferase expression, although the opposite experiment [rAAV2-Luc NDVW- (19-mer) competed with rAAV2-LacZ KGEPLSP] did. These differences are most likely due to indirect competition of these vectors for binding to neighboring receptors or limited intracellular processing as discussed below.
Collectively, our data show the utility of different-length peptide libraries for the selection of highly efficient vectors for gene transfer, demonstrating distinct properties especially regarding the transduction of nontarget cells and the use of alternative receptors for gene transfer.
Discussion
The aim of developing targeted vectors for gene therapy is to increase the efficiency and selectivity of gene transfer, especially for systemic applications. Besides efficient binding and uptake of the vectors by specific cellular receptors and adequate intracellular processing, inhibitory interactions of the vector with blood components, in particular neutralizing antibodies and components of the complement system, have to be overcome in order to achieve this goal.
Vector pseudotyping, the generation of capsid chimeras, and genetic modifications of the AAV capsid by insertion of known targeting ligands have been explored to meet at least some of the above-mentioned goals (Girod et al., 1999; P. Wu et al., 2000; Grimm et al., 2003; Gao et al., 2005; Muzyczka and Warrington, 2005; Z. Wu et al., 2006). However, as detailed knowledge about ligand–receptor interactions is often lacking, more recently two types of library-based approaches emerged that permit the selection of transduction-optimized, targeted AAV capsid variants using high-throughput vector engineering and selection schemes. One approach, the method of molecular evolution, combines fragmentation and staggered resynthesis of the cap genes of different AAV clades with mutagenesis by error-prone PCR, whereas the AAV2 peptide display system uses the presentation and subsequent selection of a library of peptide ligands at an exposed position of the AAV capsid that is naturally involved in receptor binding, thereby enabling the generation of AAV mutant capsid libraries (Müller et al., 2003; Perabo et al., 2003; Maheshri et al., 2006; Grimm et al., 2008; Koerber et al., 2008; Li et al., 2008).
The present study focused on two aspects of the AAV2 peptide display system: (a) Is it possible to generate AAV peptide display libraries of different insert length and at a different capsid position compared with the libraries described so far? (b) Which is the improvement achieved by generation of these novel libraries?
Concerning the first question, the well established three-step protocol for the production of AAV display libraries could be adapted without any problems for the generation of different-length peptide libraries, as well as for a random peptide library inserted at aa position 453 (instead of position 588) of the capsid. Analysis of the statistical representation of aa in the inserted peptides, however, revealed an underrepresentation of cysteine and phenylalanine in all libraries carrying insertions at aa position 588, whereas peptides presented at aa position 453 were mainly lacking lysine residues. Although some of these deficits seem to be linked to the capsid position of the inserted peptides, an influence of the peptide length on the incorporation of certain aa was not observed. The strongly reduced representation of cysteines observed in all libraries (including the library at aa 453) points to a disfavored incorporation of this aa independent of the insertion site or the length of the peptide, because similar deviations from an unbiased incorporation of aa into 7-mer peptide inserts at position 588 were described previously (Waterkamp et al., 2006). Furthermore, these findings are in line with the lack of capsid surface-exposed cysteine residues of wt AAV2 (Xie et al., 2002). Sequencing of a representative number of clones from each of the different libraries revealed a repeated occurrence of identical clones in the 26-mer library, strongly suggesting a biased, less efficient incorporation of peptide ligands of this length into the viral capsid. Together with the lack of transductional improvement by vectors with the 26-mer peptide inserts, these findings clearly demonstrate a limit for the construction of AAV2 peptide display libraries with increasing insert lengths.
The intended improvements achieved with these modified libraries were measured by selection of vectors on different cell types and by analysis of the gene transduction of target and various nontarget cell lines by these selected vectors. Positive selection of specific peptide-presenting capsids from each of the different-length libraries was readily achieved as indicated by a gradual enrichment of selected peptide motifs from one round of selection to the next. This result was impressively confirmed by the coselection approach where capsids with the same peptide insertions were selected from a mixed pool of all different-length libraries. Coselection of different-length libraries on HCAEC, however, seemed to favor selection of vectors with shorter peptide insertions, whereas coselection on H9C2 cells led to a preferred enrichment of vectors with 12-mer peptide insertions. The predominant enrichment of peptides harboring the identical aa motif RPG from individual screening and from library coselection points to a strong advantage in cell transduction mediated by these peptides and suggests binding to a common cellular receptor. Which parameter is decisive for the selective enrichment—size or sequence of the peptide insertion—may depend on the cell type, but a larger number of comparative studies would be required to draw definitive conclusions. Moreover, the influence of differently sized peptide insertions on the immunogenicity of the vectors remains to be explored.
The selection of a peptide library inserted at aa position 453 of the AAV2 capsid on HCAEC resulted only in a weak accumulation of specific peptide motifs. This is most likely due to the preserved presence of a strong heparin-binding motif on the AAV2 capsid that may interfere with the binding mediated by the presented peptide ligands (Boucas et al., 2009). Nevertheless, the selection of this library resulted in the enrichment of several peptide motifs, which conferred a significantly increased transduction of HCAEC compared with wt AAV2 vectors. Moreover, the isolated peptide motifs differed completely from ligands recovered from selections on the identical cell type with a 7-mer peptide library inserted at aa position 588 (Müller et al., 2003; Waterkamp et al., 2006), indicating that transduction by these vectors is most likely mediated through interactions with different cellular targets. Taken together, these findings highlight the obvious influence of the peptide insertion site as well as the surrounding capsid context on the selection outcome and clearly suggest an ablation of the heparan sulfate proteoglycan–driven tropism by site-directed mutagenesis of R585 and R588 for further applications of such libraries.
Comparative analysis of gene transduction using selected vectors from different-length libraries revealed two major results: (a) Gene transduction of target cells with most of the selected clones displaying different sequence motifs was highly improved, but, at the same time, none of the differently sized peptide insertions was per se exceptionally better than the others. (b) Transduction of nontarget cells by the vectors with different peptide insertions was clearly altered, indicating that peptides isolated from different-length libraries are able to target different cellular receptors and cell entry pathways. These differences can be expressed by a targeting index calculated from the “fold improvement” of gene transduction in target cells and the decreased gene transfer in nontarget cells, which demonstrated drastically improved retargeting efficacies especially for some of the library clones displaying 19-mer peptide ligands recovered from the selection on HCAEC. The COMPARE algorithm unequivocally demonstrated that transduction of the NCI-60 tumor cell lines with the vector selected from the 5-mer library on HCAEC correlated with the expression of different cellular proteins and receptors than transduction of these cell lines with clones selected from the 7-mer, 12-mer, or 19-mer libraries. In contrast to this, gene transduction of the NCI-60 cells by the analyzed 7-mer, 12-mer, and 19-mer clones correlated most frequently with the expression of identical cellular proteins, suggesting the use of common cellular pathways. These findings were further supported by competition experiments between rAAV2 luciferase- and β-galactosidase reporter vectors displaying the respective peptide ligands of diverse lengths, which clearly showed overlapping receptor targeting between the 12-mer and 19-mer peptides SYEN- and NDVW-, as well as partial competition of these ligands with the 7-mer motif KGEPLSP. On the other hand, vectors displaying the 5-mer ligand DIIRA were most efficiently outcompeted by vectors presenting the identical peptide. The reduced cell transduction observed after coincubation of vectors displaying DIIRA together with vectors displaying the longer peptides KGEPLSP, SYEN-(12x), or NDVW-(19x) results most likely from indirect competition. This could be due to different cell-binding efficiencies of the vectors and steric hindrance by binding to different but neighboring receptors on the cell surface or due to competition at intracellular steps of gene transfer (e.g., endosomal trafficking and nuclear transport) rather than on competition for the binding of the vectors to identical cellular receptors per se. Furthermore, a certain proportion of competition was also expected between vectors targeting different receptors, because interaction of the viral capsid with target cells is not limited to the peptide insertion site on the viral capsid, thereby leading to overlapping tropisms as discussed below.
Altogether, our results indicate that AAV peptide display libraries with inserts of different lengths allow the selection of vectors that transduce the same cell type via different receptors and entry pathways. Although the library technologies have been steadily improved, they are still at the beginning of the development of truly targeted vectors. Certainly, the screening procedures, in vitro and in vivo, are critical parameters of improvement of vector targeting and should also consider rounds of negative selection. A yet far unresolved problem for achieving targeted gene transfer is the presence of multiple receptor-binding sites on the wt AAV capsid (Kern et al., 2003; Opie et al., 2003; Asokan et al., 2006; Lochrie et al., 2006; Li et al., 2008), which inevitably leads to overlapping tropisms in most targeting efforts of AAV vectors presented so far. Identification of these sites in the capsid and knockout of all wt tropisms is therefore mandatory for true vector targeting. Furthermore, most receptors that can be used for cell entry are expressed on multiple cell types and tissues. This indicates that vector targeting by receptor selection will always be incomplete and requires complementation by promoter or microRNA targeting (Brown et al., 2006; Müller et al., 2006). A further limitation for the identification of highly selective receptor ligands on AAV capsids is the relatively low complexity of peptide display and capsid shuffling libraries. In this regard, libraries of different lengths, placed at a different position of the capsid or inserted in different serotype capsids, are able to expand the spectrum of selectable receptors for targeted gene transfer and represent a valuable enrichment for the field of AAV vector targeting.
Footnotes
Acknowledgments
We are grateful to B. Leuchs and the Vector Production Unit of the German Cancer Research Center, for providing rAAV vector preparations, and the Vector Core of the University Hospital of Nantes (France) supported by the Association Française contre les Myopathies (AFM) for providing Ad5 vector stocks. M.N. was supported by a grant of the Deutsche Forschungsgemeinschaft (KL 516/8-3).
Author Disclosure Statement
The authors declare no conflict of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.